
First Detection and Characterization of Cross- and Multiple Resistance to Acetyl-CoA Carboxylase (ACCase)- and Acetolactate Synthase (ALS)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides) and Italian Ryegrass (Lolium multiflorum) Populations from Ireland

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Materials and Growing Conditions
2.2. Single-Dose Assays
2.3. Dose-Response Assays
2.4. Analysis of Target-Site Resistance Mutations
2.5. Statistical Analysis
3. Results
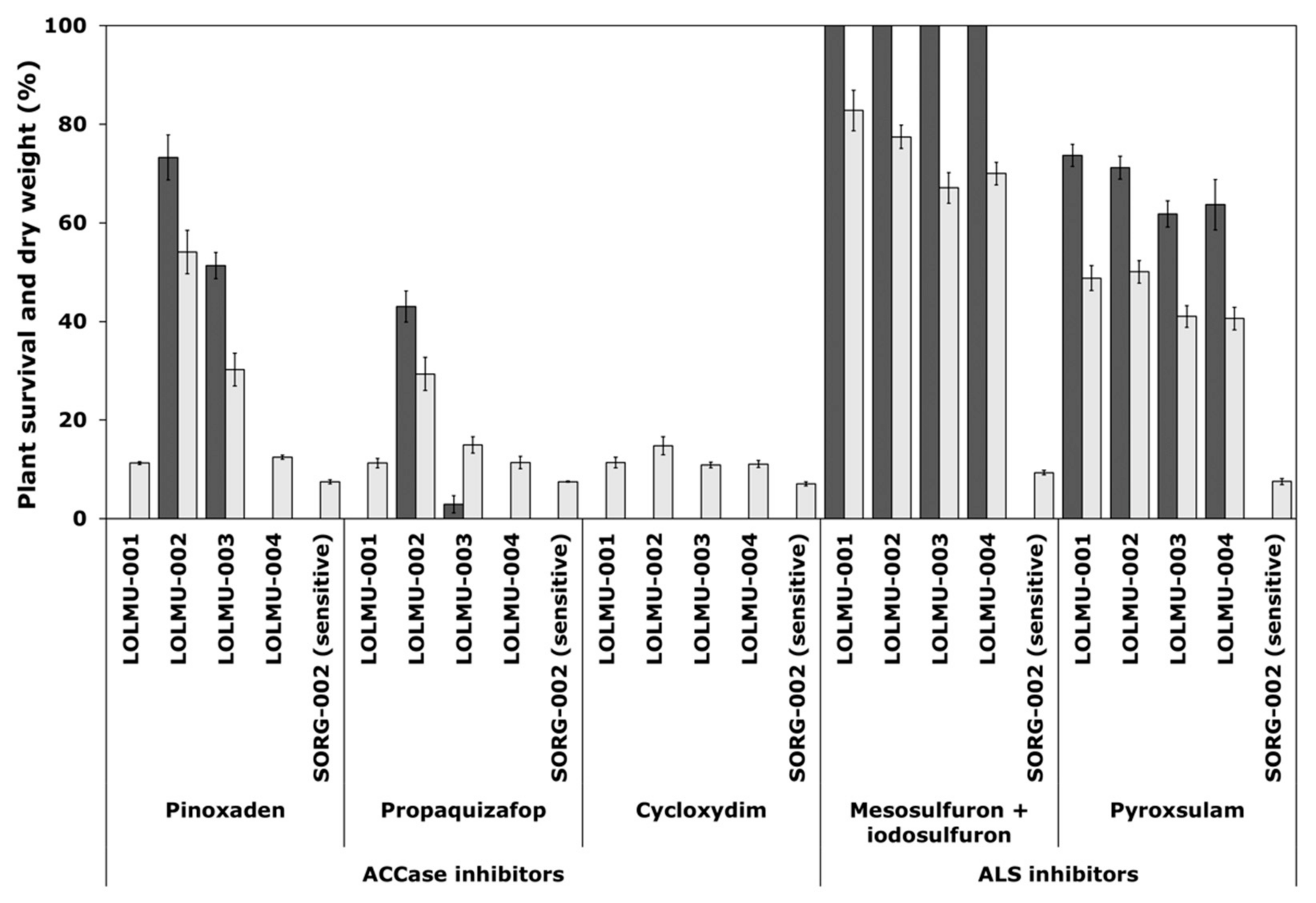
3.1. Single-Dose Assays
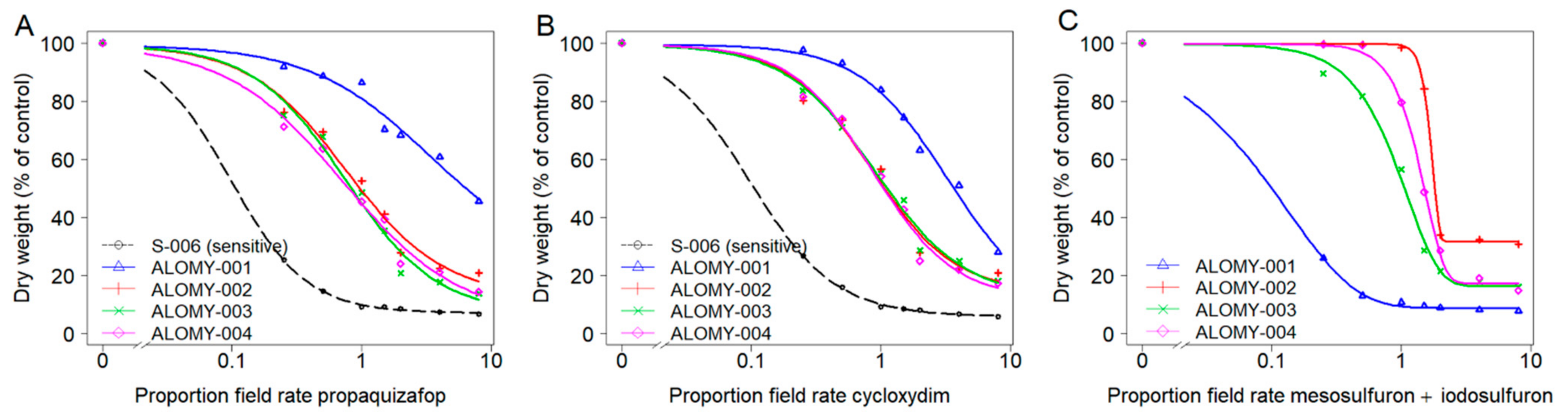
3.2. Dose-Response Assays
3.3. Analysis of Target-Site Resistance Mutations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moss, S.R.; Perryman, S.A.M.; Tatnell, L.V. Managing herbicide resistant black-grass (Alopecurus myosuroides): Theory and practice. Weed Techn. 2007, 21, 300–309. [Google Scholar] [CrossRef]
- Lutman, P.J.W.; Moss, S.R.; Cook, S.K.; Welham, S.J. A review of the effects of crop agronomy on the management of Alopecurus myosuroides. Weed Res. 2013, 53, 299–313. [Google Scholar] [CrossRef]
- Moss, S.R. Managing Herbicide-Resistant Rye-Grass: 42nd Annual Review of Weed Control; British Crop Protection Council (BCPC): Peterborough, UK, 2005; pp. 40–47. [Google Scholar]
- Moss, S.R. Black-grass (Alopecurus myosuroides): Why has this weed become such a problem in Western Europe and what are the solutions? Outlooks Pest Manag. 2017, 10, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Reverte, R. Understanding and Combating the Threat Posed by Lolium multiflorum as a Weed of Arable Crops. Ph.D. Thesis, School of Plant Sciences, University of Reading, Reading, UK, 2010. [Google Scholar]
- Délye, C.; Michel, S.; Bérard, A.; Chauvel, B.; Brunel, D.; Guillemin, J.P.; Dessaint, F.; Le Corre, V. Geographical variation in resistance to acetyl-coenzyme A carboxylase-inhibiting herbicides across the range of the arable weed Alopecurus myosuroides (black-grass). New Phytol. 2010, 186, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.S.; Welles, S.R.; Silvera, K.; Heap, I.M.; Heredia, S.M.; Martinez-Berdeja, A.; Palenscar, K.T.; Sweet, L.C.; Ellstrand, N.C. Taxonomic and life history bias in herbicide resistant weeds: Implications for deployment of resistant crops. PLoS ONE 2013, 8, e71916. [Google Scholar] [CrossRef] [Green Version]
- Moss, S.R. Herbicide cross-resistance in slender foxtail (Alopecurus myosuroides). Weed Sci. 1990, 38, 492–496. [Google Scholar] [CrossRef]
- Liu, M.; Hulting, A.G.; Mallory-Smith, C.A. Characterisation of multiple-herbicide-resistant Italian ryegrass (Lolium perenne spp. multiflorum). Pest Manag. Sci. 2014, 70, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Heap, I.M. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 15 May 2021).
- Hull, R.; Tatnell, L.V.; Cook, S.K.; Beffa, R.; Moss, S.R. Current status of herbicide-resistance weeds in the UK. Asp. Appl. Biol. 2014, 127, 261–271. [Google Scholar]
- Scarabel, L.; Panozzo, S.; Loddo, D.; Mathiassen, S.; Kristensen, M.; Kudsk, P.; Gitsopoulous, T.; Travlos, I.; Tani, E.; Chachalis, D.; et al. Diversified resistance mechanisms in multi-resistant Lolium spp. in three European countries. Front. Plant Sci. 2020, 11, 608845. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to acetyl-CoA carboxylase-inhibiting herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Délye, C.; Menchari, Y.; Guillemin, J.P.; Matéjicek, A.; Michel, S.; Camilleri, C.; Chauvel, B. Status of black-grass (Alopecurus myosuroides) resistance to acetyl-coenzyme A carboxylase inhibitors in France. Weed Res. 2007, 47, 95–105. [Google Scholar] [CrossRef]
- Marshall, R.; Moss, S.R. Characterisation and molecular basis of ALS inhibitor resistance in the grass weed Alopecurus myosuroides. Weed Res. 2008, 48, 439–447. [Google Scholar] [CrossRef]
- Hicks, H.L.; Comont, D.; Coutts, S.R.; Crook, L.; Hull, R.; Norris, K.; Neve, P.; Childs, D.Z.; Freckleton, R.P. The factors driving evolved herbicide resistance at a national scale. Nat. Ecol. Evol. 2018, 2, 520–536. [Google Scholar] [CrossRef] [Green Version]
- Scarabel, L.; Panozzo, S.; Varotto, S.; Sattin, M. Allelic variation of the ACCase gene and response to ACCase-inhibiting herbicides in pinoxaden-resistant Lolium spp. Pest Manag. Sci. 2011, 67, 932–941. [Google Scholar] [CrossRef]
- CSO. Structure of Farming in Ireland 2016, Central Statistics Office (CSO). Available online: https://www.cso.ie/en/releasesandpublications/ep/p-fss/farmstructuresurvey2016/ (accessed on 18 June 2021).
- Forristal, D.; Grant, J. The impact of break-crop and cereal rotations on crop performance and profit margin: Making Crop rotations fit for the future. Asp. Appl. Biol. 2011, 113, 29–36. Available online: https://www.aab.org.uk/aspects-of-applied-biology/ (accessed on 21 June 2021).
- BSBI. Botanical Society of Britain and Ireland (BSBI); BSBI: Durham, UK, 2021. [Google Scholar]
- DAFM. Pesticide Usage in Ireland, Arable Crops Survey Report 2016, Pesticide Registration and Control Divisions (PRCD) of the Department of Agriculture; Food and the Marine (DAFM), Co.: Kildare, Ireland, 2016. Available online: https://www.pcs.agriculture.gov.ie/media/pesticides/content/sud/pesticidestatistics/ArableReport2016Final100620.pdf (accessed on 21 June 2021).
- Alwarnaidu Vijayarajan, V.B.; Forristal, D.P.; Cook, S.K.; Staples, J.; Schilder, D.; Hennessy, M.; Barth, S. First report on assessing the severity of herbicide resistance to ACCase inhibitors pinoxaden, propaquizafop and cycloxydim in six Avena fatua populations in Ireland. Agronomy 2020, 10, 1362. [Google Scholar] [CrossRef]
- Hess, M.; Barralis, G.; Bleiholder, H.; Buhrs, L.; Eggers, T.H.; Hack, H.; Strauss, R. Use of the extended BBCH scale-general for the descriptions of the growth stages of mono- and dicotyledonous weed species. Weed Res. 1997, 37, 433–441. [Google Scholar] [CrossRef]
- Keshtkar, E.; Mathiassen, S.K.; Moss, S.R.; Kudsk, P. Resistance profile of herbicide-resistant Alopecurus myosuroides (black-grass) populations in Denmark. Crop Prot. 2015, 69, 83–89. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Tranel, P.J.; Wright, T.R.; Heap, I.M. Mutations in Herbicide-Resistant Weeds to ALS Inhibitors. Available online: http://www.weedscience.com (accessed on 18 May 2021).
- Yu, Q.; Han, H.; Powles, S.B. Mutations of the ALS gene endowing resistance to ALS-inhibiting herbicides in Lolium rigidum populations. Pest Manag. Sci. 2008, 64, 1229–1236. [Google Scholar] [CrossRef]
- Henriet, F.; Marechal, P.Y. Black-grass resistance to herbicides: Three years of monitoring in Belgium. Commun. Agric. Appl. Biol. Sci. 2009, 74, 471–478. [Google Scholar]
- Marshall, R.; Hanley, S.J.; Hull, R.; Moss, S.R. The presence of two different target-site resistance mechanisms in individual plants of Alopecurus myosuroides Huds, identified using a quick molecular test for the characterization of six and seven ACCase SNPs. Pest Manag. Sci. 2013, 69, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Torra, J.; Montull, J.M.; Taberner, A.; Onkokesung, N.; Boonham, N.; Edwards, R. Target-site and non-target-site resistance mechanisms confer multiple and cross- resistance to ALS and ACCase inhibiting herbicides in Lolium rigidum From Spain. Front. Plant Sci. 2021, 12, 625138. [Google Scholar] [CrossRef] [PubMed]
- Collavo, A.; Sattin, M. First glyphosate-resistant Lolium spp. biotypes found in a European annual arable cropping systems also affected by ACCase and ALS resistance. Weed Res. 2014, 54, 325–334. [Google Scholar] [CrossRef]
- Tardif, F.J.; Powles, S.B. Herbicide multiple-resistance in a Lolium rigidum biotype is endowed by multiple mechanisms-Isolation of a subset with resistant acetyl-CoA carboxylase. Physiol. Plant 1994, 91, 488–494. [Google Scholar] [CrossRef]
- Baucom, R.S. Evolutionary and ecological insights from herbicide resistant weeds: What have we learned about plant adaptation, and what is left to uncover? New Phytol. 2019, 223, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.S.; Tranel, P.J.; Stewart, C.N. Non-target-site herbicide resistance: A family business. Trends Plant Sci. 2007, 12, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Cocker, K.M.; Coleman, J.O.D.; Blair, A.M.; Clarke, J.H.; Moss, S.R. Biochemical mechanisms of cross-resistance to aryloxyphenoxypropionate and cyclohexanediones herbicides in populations of Avena spp. (wild oats). Weed Res. 2000, 40, 323–334. [Google Scholar] [CrossRef]
- Owen, M.; Goggin, D.E.; Powles, S.B. Non-target-site-based resistance to ALS-inhibiting herbicides in six Bromus rigidus populations from Western Australia cropping fields. Pest Manag. Sci. 2012, 68, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.; Onkokesung, N.; Brazier-Hicks, M.; Edwards, R.; Moss, S. Detection and characterization of resistance to acetolactate synthase inhibiting herbicides in Anisantha and Bromus species in the United Kingdom. Pest Manag. Sci. 2020, 76, 2473–2482. [Google Scholar] [CrossRef] [PubMed]
- Cocker, K.M.; Northcroft, D.S.; Coleman, J.O.D.; Moss, S.R. Resistance to ACCase-inhibiting herbicides and isoproturon in UK populations of Lolium multiflorum: Mechanisms of resistance and implications for control. Pest Manag. Sci. 2001, 57, 587–597. [Google Scholar] [CrossRef]
- Petit, C.; Duhieu, B.; Boucansaud, K.; Délye, C. Complex genetic control of non-target-site based resistance to herbicides inhibiting acetyl-coenzyme A carboxylase and acetolactate synthase in Alopecurus myosuroides Huds. Plant Sci. 2010, 178, 501–509. [Google Scholar] [CrossRef]
- Moss, S.R.; Riches, C.; Stormonth, D. Clethodim: It’s potential to combat herbicide-resistant Alopecurus myosuroides (black-grass). Asp. Appl. Biol. 2012, 117, 39–45. [Google Scholar]
- Yu, Q.; Collavo, A.; Zheng, M.; Owen, M.; Sattin, M.; Powles, S.B. Diversity of acetyl-coenzyme a carboxylase mutations in resistant Lolium populations: Evaluation using clethodim. Plant Physiol. 2007, 145, 547–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauvel, B.; Guillemin, J.P.; Colbach, N. Evolution of a herbicide-resistant population of Alopecurus myosuroides Huds in a long-term cropping system experiment. Crop Prot. 2009, 28, 343–349. [Google Scholar] [CrossRef]
- Varah, A.; Ahodo, K.; Coutts, S.R.; Hicks, H.L.; Comont, D.; Crook, L.; Hull, R.; Neve, P.; Childs, D.Z.; Freckleton, R.; et al. The cost of human-induced evolution in an agricultural system. Nat. Sustain. 2020, 3, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.R. Integrated weed management (IWM): Why are farmers reluctant to adopt non-chemical alternatives to herbicides? Pest Manag. Sci. 2019, 75, 1205–1211. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | County | Field Position | Year | Establishment System | Crop | ALOMY-Active Herbicides Applied |
|---|---|---|---|---|---|---|
| ALOMY-001 | Cork | 51°71′ N–8°65′ W | 2016–2020 | Plough | Winter barley | None |
| ALOMY-002 | Meath | 53°48′ N–6°41′ W | 2020 2019 2018 2017 2016 | Plough Plough Plough Plough Plough | Spring barley Winter wheat Winter wheat Winter oats Winter wheat | None |
| ALOMY-003 | Meath | 53°56′ N–6°58′ W | 2016–2020 | Plough | Winter wheat | Mesosulfuron + iodosulfuron applied annually |
| ALOMY-004 | Waterford | 52°22′ N–7°42′ W | 2020 2019 2018 2017 2016 | NIT NIT NIT Plough Plough | Spring barley Spring barley Spring barley Winter wheat Spring oats | None None None Mesosulfuron + iodosulfuron None |
| S-006 | Dublin | 53°56′ N–6°29′ W | 2020 2019 2018 2017 2016 | Plough Plough Plough Plough Plough | Winter oats Winter wheat Winter oilseed rape Winter wheat Spring beans | None Mesosulfuron + iodosulfuron Propaquizafop Mesosulfuron + iodosulfuron Propaquizafop |
| Population | County | Field Position | Year | Establishment System | Crop | LOLMU-Active Herbicides Applied |
|---|---|---|---|---|---|---|
| LOLMU-001 | Tipperary | 52°35′ N–7°75′ W | 2020 2019 2018 2017 2016 | NIT NIT NIT NIT NIT | Winter wheat Winter wheat Spring oats Winter wheat Winter wheat | Pyroxsulam Mesosulfuron + iodosulfuron None Mesosulfuron + iodosulfuron Mesosulfuron + iodosulfuron |
| LOLMU-002 | Cork | 52°18′ N–8°84′ W | 2020 2019 2018 2017 2016 | NIT DD NIT NIT NIT | Winter wheat Spring beans Winter barley Winter barley Winter barley | Pyroxsulam Cycloxydim Pinoxaden Pinoxaden Pinoxaden |
| LOLMU-003 | Meath | 53°48′ N–6°42′ W | 2020 2019 2018 2017 2016 | Plough Plough Plough Plough Plough | Winter wheat Winter wheat Winter wheat Winter wheat Winter wheat | Mesosulfuron + iodosulfuron Pyroxsulam Mesosulfuron + iodosulfuron Mesosulfuron + iodosulfuron Pyroxsulam |
| LOLMU-004 | Meath | 53°65′ N–6°36′ W | 2020 2019 2018 2017 2016 | Plough Plough Plough Plough Plough | Winter wheat Winter wheat Winter oats Winter wheat Winter wheat | Pinoxaden Pinoxaden None Pinoxaden Pinoxaden |
| Mode of Action | Chemical Families | Herbicide | Dose (g Active Substance ha−1) |
|---|---|---|---|
| Alopecurus myosuroides | |||
| ACCase | FOP | Propaquizafop | 25, 50, 100, 150, 200, 400, 800 and 0 |
| DIM | Cycloxydim | 50, 100, 200, 300, 400, 800, 1600 and 0 | |
| ALS | SU | Mesosulfuron + iodosulfuron | 3.8 + 1.3, 7.5 + 2.5, 15 + 5, 22.5 + 7.5, 30 + 10, 60 + 20, 120 + 40 and 0 |
| Lolium multiflorum | |||
| ACCase | DEN | Pinoxaden | 7.6, 15.1, 30.3, 45.4, 60.5, 121, 242 and 0 |
| FOP | Propaquizafop | 37.5, 75, 150, 225, 300, 600, 1200 and 0 | |
| DIM | Cycloxydim | 37.5, 75, 150, 225, 300, 600, 1200 and 0 | |
| ALS | SU | Mesosulfuron + iodosulfuron | 3.8 + 1.3, 7.5 + 2.5, 15 + 5, 22.5 + 7.5, 30 + 10, 60 + 20, 120 + 40 and 0 |
| TP | Pyroxsulam | 4.7, 9.4, 18.8, 28.1, 37.5, 75.0, 150.8 and 0 |
| Propaquizafop | Cycloxydim | Mesosulfuron + Iodosulfuron | Mutant ACCase | Mutant ALS | ||||
|---|---|---|---|---|---|---|---|---|
| Population | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | Ile-1781 | Pro-197 |
| S-006 | 9.6 (5.24) | - | 19.2 (8.65) | - | <3.8 + <1.3 | - | - | - |
| ALOMY-001 | 340.6 (297.2) | 35.5 | 653.8 (237.5) | 34.1 | 1.3 (0.6) + 0.4 (0.2) | >0.3 | (6/2) | - |
| ALOMY-002 | 75.6 (10.56) | 7.9 | 167.1 (17.91) | 8.7 | 25.1 (0.49) + 8.4 (0.16) | >6.6 | (5/3) | (1/7) |
| ALOMY-003 | 74.2 (8.87) | 7.7 | 178.9 (22.29) | 9.3 | 13.6 (0.52) + 4.5 (0.17) | >3.6 | (2/3) | (0/5) |
| ALOMY-004 | 71.7 (13.53) | 7.5 | 173.1 (17.8) | 9 | 20.4 (0.49) + 6.8 (0.16) | >5.4 | (0/7) | (0/7) |
| Pinoxaden | Propaquizafop | Cycloxydim | Mesosulfuron + Iodosulfuron | Pyroxsulam | Mutant ALS | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Population | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | GR50 (g a.s ha−1) | RF | Pro-197 |
| SORG-002 | <7.6 | - | <37.5 | - | <37.5 | - | 1.3 (0.24) + 0.4 (0.08) | - | <4.7 | - | - |
| LOLMU-001 | 1.6 (0.54) | >0.2 | 4.3 (1.68) | >0.1 | 5.7 (1.79) | >0.2 | 19 (0.63) + 6.3 (0.21) | 14.6 | 13.6 (0.65) | >2.9 | (6/2) |
| LOLMU-002 | 25.9 (1.92) | >3.4 | 49.7 (4.65) | >1.3 | 18.3 (1.61) | >0.5 | 17.8 (0.52) + 5.9 (0.17) | 13.7 | 14.5 (0.74) | >3.1 | (3/1) |
| LOLMU-003 | 17.3 (1.15) | >2.3 | 11.1 (2.39) | >0.3 | 10.9 (1.85) | >0.3 | 15.5 (0.64) + 5.2 (0.21) | 11.9 | 10.8 (0.6) | >2.3 | - |
| LOLMU-004 | 1.4 (0.48) | >0.2 | 3.3 (1.63) | >0.1 | 5.4 (1.84) | >0.1 | 16.5 (0.6) + 5.5 (0.2) | 12.7 | 11.8 (0.56) | >2.5 | (5/0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alwarnaidu Vijayarajan, V.B.; Forristal, P.D.; Cook, S.K.; Schilder, D.; Staples, J.; Hennessy, M.; Barth, S. First Detection and Characterization of Cross- and Multiple Resistance to Acetyl-CoA Carboxylase (ACCase)- and Acetolactate Synthase (ALS)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides) and Italian Ryegrass (Lolium multiflorum) Populations from Ireland. Agriculture 2021, 11, 1272. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121272
Alwarnaidu Vijayarajan VB, Forristal PD, Cook SK, Schilder D, Staples J, Hennessy M, Barth S. First Detection and Characterization of Cross- and Multiple Resistance to Acetyl-CoA Carboxylase (ACCase)- and Acetolactate Synthase (ALS)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides) and Italian Ryegrass (Lolium multiflorum) Populations from Ireland. Agriculture. 2021; 11(12):1272. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121272
Chicago/Turabian StyleAlwarnaidu Vijayarajan, Vijaya Bhaskar, Patrick D Forristal, Sarah K Cook, David Schilder, Jimmy Staples, Michael Hennessy, and Susanne Barth. 2021. "First Detection and Characterization of Cross- and Multiple Resistance to Acetyl-CoA Carboxylase (ACCase)- and Acetolactate Synthase (ALS)-Inhibiting Herbicides in Black-Grass (Alopecurus myosuroides) and Italian Ryegrass (Lolium multiflorum) Populations from Ireland" Agriculture 11, no. 12: 1272. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121272