Estimation of Genetic Diversity in Seven Races of Native Maize from the Highlands of Mexico

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Field Assessments
2.3. Morphological Traits Measured
2.4. Assessment of Microsatellite Polymorphism
2.5. Statistical Analysis
3. Results
3.1. Analysis of Variance
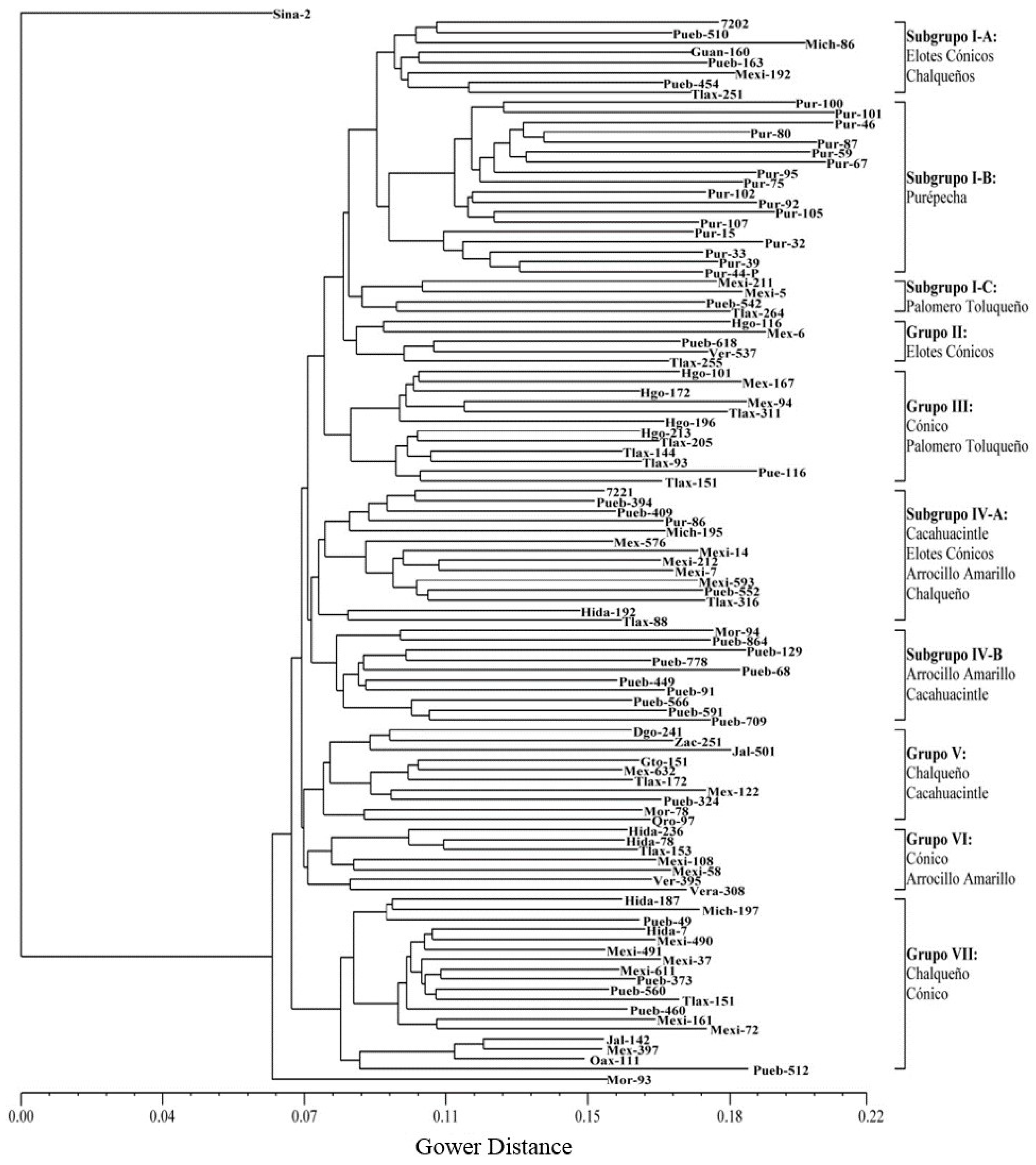
3.2. Cluster and Principal Component Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kato, Y.T.A.; Mapes, C.S.; Mera, L.M.O.; Serratos, J.A.H.; Bye, R.A.B. Origen y Diversificación del Maíz: Una Revisión Analítica; Universidad Nacional Autónoma de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2009; p. 116. [Google Scholar]
- Wellhausen, E.J.; Roberts, L.M.; Hernández, X.E.; Mangelsdorf, P.C. Races of Maize in México, their Origin, Characteristics and Distribution; The Bussey Institution of Harvard University: Cambridge, UK, 1952; p. 223. [Google Scholar]
- Hernández, X.E.; Alanís, F.G. Morphological study of five new maize races of the Sierra Madre Occidental of Mexico. Phylogenetic and phytogeographic implications. Agrociencia 1970, 5, 3–30. [Google Scholar]
- Cervantes, S.T.; Goodman, M.M.; Casas, D.E.; Rawlings, J.O. Use of genetic effects and genotype by environmental interactions for the classification of Mexican races of maize. Genetics 1978, 90, 339–348. [Google Scholar] [PubMed]
- Sánchez, G.J.J.; Goodman, M.M.; Rawlings, J.O. Appropriate characters for racial classification in maize. Econ. Bot. 1993, 47, 44–59. [Google Scholar] [CrossRef]
- Goodman, M.M.; Paterniani, E. The races of maize: III. Choices of appropriate characters for racial classification. Econ. Bot. 1969, 23, 265–273. [Google Scholar] [CrossRef]
- Doebley, J.F.; Iltis, H.H. Taxonomy of Zea (Gramineae). I. A subgeneric classification with key to taxa. Am. J. Bot. 1980, 67, 982–993. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Vigoroux, Y.; Goodman, M.M.; Sánchez, G.J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reif, J.C.; Waburton, M.L.; Xia, X.C.; Hoisington, D.A.; Crossa, J.; Taba, S.; Muminović, J.; Bohn, M.; Frisch, M.; Melchinger, A.E. Grouping of accessions of Mexican races of maize revisited with SSR markers. Theor. Appl. Genet. 2006, 113, 177–185. [Google Scholar] [CrossRef]
- Vigouroux, Y.; Glaubitz, J.C.; Matsuoka, Y.; Goodman, M.M.; Sánchez, G.J.; Doebley, J. Population structure and genetic diversity of New World maize races assessed by DNA microsatellites. Am. J. Bot. 2008, 95, 1240–1253. [Google Scholar] [CrossRef]
- Rocandio-Rodríguez, M.; Santacruz-Varela, A.; Córdova-Téllez, L.; López-Sánchez, H.; Castillo-González, F.; Lobato-Ortiz, R.; García-Zavala, J.J. Detection of genetic diversity of seven maize races from the high central valleys of Mexico using microsatellites. Maydica 2014, 59, 144–151. [Google Scholar]
- Vega-Alvarez, I.; Santacruz-Varela, A.; Rocandio-Rodríguez, M.; Córdova-Téllez, L.; López-Sánchez, H.; Muñoz-Orozco, A.; Hernández-Bautista, A. Genetic diversity and structure of native maize races from Northwestern Mexico. Pesq. Agropec. Bras. 2017, 52, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, G.J.J.; Goodman, M.M. Relationships among the Mexican races of maize. Econ. Bot. 1992, 46, 72–85. [Google Scholar] [CrossRef]
- Rocandio-Rodríguez, M.; Santacruz-Varela, A.; Córdova-Téllez, L.; López-Sánchez, H.; Castillo-González, F.; Lobato-Ortiz, R.; García-Zavala, J.J.; Ortega-Paczka, R. Morphological and agronomic characterization of seven maize races from the highlands of México. Rev. Fitotec. Mex. 2014, 37, 351–361. [Google Scholar]
- Sánchez, G.J.J.; Goodman, M.M.; Stuber, C.W. Isozymatic and morphological diversity in the races of maize of Mexico. Econ. Bot. 2000, 54, 43–59. [Google Scholar] [CrossRef]
- Mijangos-Cortés, J.O.; Corona-Torres, T.; Espinosa-Victoria, D.; Muñoz-Orozco, A.; Romero-Peñaloza, J.; Santacruz-Varela, A. Differentiation among maize (Zea mays L.) landraces from the Tarasca Mountain Chain, Michoacan, Mexico and the Chalqueño complex. Genet. Resour. Crop Evol. 2007, 54, 309–325. [Google Scholar] [CrossRef]
- Santacruz-Varela, A.; Widrlechner, M.P.; Ziegler, K.E.; Salvador, R.J.; Millard, M.J.; Bretting, P.K. Phylogenetic relationships among North American popcorns and their evolutionary links to Mexican and South American popcorns. Crop. Sci. 2004, 44, 1456–1467. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.J.; Hanotte, O.; Guarino, L. Standardization of molecular genetic techniques for the characterization of germplasm collections: The case of random amplified polymorphic DNA (RAPD). Plant Genet. Resour. Newsl. 1996, 107, 50–54. [Google Scholar]
- Pan, Y.B.; Burner, D.M.; Legendre, B.L.; Grisham, M.P.; White, W.H. An assessment of the genetic diversity within a collection of Saccharum spontaneum L. with RAPD-PCR. Genet. Resour. Crop Evol. 2004, 51, 895–903. [Google Scholar] [CrossRef]
- Avise, J.C. Molecular Markers, Natural History and Evolution, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2004; p. 684. [Google Scholar]
- De Queiroz, A.; Donoghue, M.J.; Kim, J. Separate versus combined analysis of phylogenetic evidence. Annu. Rev. Ecol. Evol. Syst. 1995, 26, 657–681. [Google Scholar] [CrossRef]
- Chippindale, P.T.; Wiens, J.J. Weighting, partitioning, and combining characters in phylogenetic analysis. Syst. Biol. 1994, 43, 278–287. [Google Scholar] [CrossRef]
- Leal, A.A.; Mangolin, C.A.; do Amaral, A.T., Jr.; Gonçalves, L.S.A.; Scapim, C.A.; Mott, A.S.; Eloi, I.B.O.; Cordovés, V.; da Silva, M.F.P. Efficiency of RAPD versus SSR markers for determining genetic diversity among popcorn lines. Genet. Mol. Res. 2010, 9, 9–18. [Google Scholar] [CrossRef]
- Kluge, A.G. A concern for evidence and a phylogenetic hypothesis of relationships among Epicrates (Boidae, Serpentes). Syst. Zool. 1989, 38, 7–25. [Google Scholar] [CrossRef]
- Kluge, A.G.; Wolf, A.J. Cladistics: What’s in a word? Cladistics 1993, 9, 183–199. [Google Scholar] [CrossRef]
- Nixon, K.C.; Carpenter, J.M. On simultaneous analysis. Cladistics 1996, 12, 221–241. [Google Scholar] [CrossRef]
- Gulanbaier, T.; Seoung, B.M. Gower distance-based multivariate control charts for a mixture of continuous and categorical variables. Expert Syst. Appl. 2014, 41, 1701–1707. [Google Scholar]
- Applied, B. GeneMapper® Software Version 4.0. Reference and Troubleshooting Guide; Applied Biosystems Inc.: Foster City, CA, USA, 2005; p. 82. [Google Scholar]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Version 1.31. Microsoft Window-based Freeware for Population Genetic Analysis, Quik User Guide; University of Alberta and Center for International Forestry Research: Edmonton, AB, Canada, 1999; Available online: http://www.ualberta.ca/~fyeh/popgene.pdf (accessed on 23 March 2018).
- SAS, Institute. SAS/STAT User’s Guide, Software version 9.0.; SAS Institute Inc.: Cary, NC, USA, 2002; p. 4424. [Google Scholar]
- Martínez, G.A. Manual de diseño y análisis de los látices. Monografías y Manuales de Estadística y Cómputo. Vol. 8, Núm. 3; Centro de Estadística y Cálculo, Colegio de Postgraduados: Chapingo, Mexico, 1989; p. 71. [Google Scholar]
- Pla, L.E. Análisis Multivariado: Métodos de Componentes Principales. Programa Regional de Desarrollo Científico y Tecnológico; Secretaria General de la Organización de los Estados Americanos: Washington, DC, USA, 1986; p. 94. [Google Scholar]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–874. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Rohlf, F.J. NTSYSpc: Numerical Taxonomy System, Version 2.21c; Exeter Software, Setauket: New York, NY, USA, 2009. [Google Scholar]
- Herrera, C.B.E.; Castillo, G.F.; Sánchez, G.J.J.; Ortega, P.R.; Goodman, M.M. Morphological traits to assess diversity among maize populations in a region: Chalqueño landrace case. Rev. Fitotec. Mex. 2000, 23, 335–354. [Google Scholar]
- López-Romero, G.; Santacruz-Varela, A.; Muñoz-Orozco, A.; Castillo-González, F.; Córdova-Téllez, L.; Vaquera-Huerta, H. Caracterización morfológica de poblaciones nativas de maíz del Istmo de Tehuantepec, México. Interciencia 2005, 30, 284–290. [Google Scholar]
- Maturana, M.S.; Sánchez, C.J. Las comunidades de la Meseta Tarasca, Un Estudio Socioeconómico; Centro de Investigaciones Agrarias: Mexico City, Mexico, 1970; p. 124. [Google Scholar]
- Romero, P.J.; Castillo, G.F.; Ortega, P.R. Crosses of native populations of maize from Chalqueño race: II. Genetic groups, genetic divergence and heterosis. Rev. Fitotec. Mex. 2002, 25, 107–115. [Google Scholar]
- Doebley, J.F.; Goodman, M.M.; Stuber, C.W. Isozyme variation in the races of maize from México. Am. J. Bot. 1985, 72, 629–639. [Google Scholar] [CrossRef]
- Ángeles-Gaspar, E.; Ortiz-Torres, E.; López, P.A.; López-Romero, G. Characterization and yield performance of native maize populations from Molcaxac, Puebla. Rev. Fitotec. Mex. 2010, 33, 287–296. [Google Scholar]
- Myles, S.; Peiffer, J.; Brown, P.J.; Ersoz, E.S.; Zhang, Z.; Costich, D.E.; Buckler, E.S. Association mapping: Critical considerations shift from genotyping to experimental design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Traits |
|---|
| Phenological traits: days to male (MF) and female (FF) flowering; anthesis-silking interval (ASI). Vegetative traits: number of tillers per plant (SP), plant height (PH), ear height (EH), length (LL) and width (LW) of the leaf at the primary ear in cm, total number of leaves (NL) and number of leaves above the ear (NLE). Tassel traits: peduncle length (PLS), length of the branched part of the tassel (LBT), length of the central spike (LCS), total tassel length (TL) in cm, number of primary branches (PB). Ear traits: length (EL) and ear diameter (ED); peduncle length (PLE); cob diameter (CD), in cm, number of kernel rows (KR), ear grain percentage (EGP = (grain weight/total weight of maize ear) × 100). Kernel traits: length (KL), width (KW) and thickness (KT) in mm in 10 kernels of each of the five ears of the sampled plants, weight of 100 kernels (W100K) in g, volume of 100 kernels (V100K) in mL. Ratios: EH/PH, ED/EL, LBT/TL, KW/KL, KT/KL and W100K/V100K. |
| Locus | N. Bin. | Forward Primer//Reverse Primer |
|---|---|---|
| phi127 | 2.07 | ROX-ATATGCATTGCCTGGAACTGGAAGGA//AATTCAAACACGCCTCCCGAGTGT |
| phi051 | 7.06 | 6-FAM-GCGAAAGCGAACGACAACAATCTT//ACATCGTCAGATTATATTGCAGACCA |
| phi115 | 8.03 | HEX-GCTCCGTGTTTCGCCTGAA//ACCATCACCTGAATCCATCACA |
| phi015 | 8.08 | HEX-GCAACGTACCGTACCTTTCCGA//ACGCTGCATTCAATTACCGGGAAG |
| phi033 | 9.02 | 6-FAM-ATCGAAATGCAGGCGATGGTTCTC//ATCGAGATGTTCTACGCCCTGAAGT |
| phi053 | 3.05 | ROX-CTGCCTCTCAGATTCAGAGATTGAC//AACCCAACGTACTCCGGCAG |
| phi072 | 4.01 | 6-FAM-GTGCATGATTAATTTCTCCAGCCTT//GACAGCGCGCAAATGGATTGAACT |
| phi093 | 4.08 | ROX-GTGCGTCAGCTTCATCGCCTACAAG//CCATGCATGCTTGCAACAATGGATACA |
| phi024 | 5.01 | HEX-CTCCGCTTCCACTGTTCCA//TGTCCGCTGCTTCTACCCA |
| phi085 | 5.06 | 6-FAM-AGCAGAACGGCAAGGGCTACT//TTTGGCACACCACGACGA |
| phi034 | 7.02 | HEX-TAGCGACAGGATGGCCTCTTCT//GGGGAGCACGCCTTCGTTCT |
| phi121 | 8.04 | 6-FAM-AGGAAAATGGAGCCGGTGAACCA//TTGGTCTGGACCAAGCACATACAC |
| phi056 | 1.01 | ROX-ACTTGCTTGCCTGCCGTTAC//CGCACACCACTTCCCAGAA |
| phi064 | 1.11 | HEX-CGAATTGAAATAGCTGCGAGAACCT//ACAATGAACGGTGGTTATCAACACGC |
| phi050 | 10.03 | ROX-AACATGCCAGACACATACGGACAG//ATGGCTCTAGCGAAGCGTAGAG |
| phi96100 | 2.01 | 6-FAM-AGGAGGACCCCAACTCCTG//TTGCACGAGCCATCGTAT |
| phi101249 | Unknown | 6-FAM-TTCCTCCTCCACTGCCTC//AAGAACAGCGAAGCAGAGAAGG |
| phi109188 | 5.03 | HEX-AAGCTCAGAAGCCGGAGC//GGTCATCAAGCTCTCTGATCG |
| phi029 | 3.04 | ROX-TCTTTCTTCCTCCACAAGCAGCGAA//TTTCCAGTTGCCACCGACGAAGAACTT |
| phi073 | 3.05 | HEX-GTGCGAGAGGCTTGACCAA//AAGGGTTGAGGGCGAGGAA |
| phi96342 | 10.02 | 6-FAM-GTAATCCCACGTCCTATCAGCC//TCCAACTTGAACGAACTCCTC |
| phi109275 | 1.03 | 6-FAM-CGGTTCATGCTAGCTCTGC//GTTGTGGCTGTGGTGGTG |
| phi427913 | 1.01 | ROX-CAAAAGCTAGTCGGGGTCA//ATTGTTCGATGACACACTACGC |
| phi265454 | 1.11 | 6-FAM-CAAGCACCTCAACCTCTTCG//TCCACGCTGCTCACCTTC |
| phi402893 | 2.XX | HEX-GCCAAGCTCAGGGTCAAG//CACGAGCGTTATTCGCTGT |
| phi346482 | 1.XX | HEX-GCATCACACTTCACACAACAA//GTGGAATAGGAGGCGAGAGAGG |
| phi308090 | 4.04–4.05 | 6-FAM-CAGTCTGCCACGAAGCAA//CTGTCGGTTTCGGTCTTCTT |
| phi330507 | 5.02–5.06 | ROX-GTAAAGTACGATGCGCCTCCC//CGGGGTAGAGGAGAGTTGTG |
| phi213398 | 4.01–4.04 | 6-FAM-GTGACCTAAACTTGGCAGACCC//CAAGAGGTACCTGCATGGC |
| phi339017 | 1.03 | HEX-ACTGCTGTTGGGGTAGGG//GCAGCTTGAGCAGGAAGC |
| phi159819 | 6.00–6.08 | 6-FAM-GATGGGCCCTAGACCAGCTT//GCCTCTCCCATCTCTCGGT |
| Trait | Populations | Localities | Pob × Loc | Error | Mean | Standard Deviation |
|---|---|---|---|---|---|---|
| Male flowering | 932.5 ** | 26,311.7 ** | 39.4 ** | 13.5 | 100.64 | 15.79 |
| Female flowering | 813.2 ** | 34,724.2 ** | 33.0 ** | 15.7 | 93.46 | 15.71 |
| Anthesis-silking interval | 33.5 ** | 659.8 ** | 9.965 ** | 5.5 | 7.18 | 3.65 |
| Shoots per plant | 0.1 ** | 15.0 ** | 0.1 ** | 0.1 | 0.21 | 0.32 |
| Plant height | 10,455.7 ** | 421,681.3 ** | 330.9 ns | 288.2 | 250.23 | 56.16 |
| Ear height | 5441.2 ** | 326,631.3 ** | 254.4 ** | 206.4 | 119.72 | 44.63 |
| Ear height/plant height | 0.0 ** | 1.2 ** | 0.0 ** | 0.0 | 0.46 | 0.09 |
| Number of leaves | 21.5 ** | 25.4 ns | 1.2 ** | 0.6 | 12.06 | 2.08 |
| Number of leaves above ear | 2.1 ** | 10.6 ** | 0.1 ** | 0.1 | 4.81 | 0.69 |
| Leaf length | 393.7 ** | 57,622.6 ** | 57.5 ** | 29.2 | 75.08 | 16.07 |
| Leaf width | 7.7 ** | 955.6 ** | 1.7 * | 1.4 | 8.81 | 2.28 |
| Total length | 155.3 ** | 210.7 ns | 30.0 ** | 25.6 | 63.27 | 6.85 |
| Peduncle length of the tassel | 30.6 ** | 248.5 ns | 12.9 ns | 11.1 | 25.21 | 3.97 |
| Length of central spike | 64.6 ** | 265.0 ** | 10.6 ** | 7.7 | 31.58 | 4.34 |
| Length of branched part of tassel | 23.6 ** | 1190.5 ** | 2.4 ns | 2.2 | 6.65 | 3.02 |
| Length of branched part of tassel/total length | 0.0 ** | 0.3 ** | 0.0 ** | 0.0 | 0.10 | 0.05 |
| Number of primary branches | 38.2 ** | 1180.2 ** | 6.2 ** | 4.8 | 8.17 | 3.75 |
| Ear length | 34.8 ** | 423.7 ** | 2.9 ns | 2.7 | 14.54 | 3.05 |
| Number of kernel rows | 25.0 ** | 186.7 ** | 1.3 ** | 1. | 14.48 | 2.37 |
| Peduncle length of the ear | 26.1 ** | 1549.9 ** | 8.5** | 6. | 12.01 | 3.83 |
| Ear diameter | 0.9 ** | 34.6 ** | 0.1 ** | 0.1 | 4.11 | 0.56 |
| Ear diameter/ear length | 0.0 ** | 0.0 ns | 0.0 ** | 0.0 | 0.29 | 0.05 |
| Cob diameter | 0.6 ** | 12.3 ** | 0.1 ** | 0.1 | 1.95 | 0.45 |
| Kernel length | 783.1 ** | 14,837.6 ** | 97.8 ** | 64.1 | 12.67 | 1.54 |
| Kernel width | 826.7 ** | 1500.4 ** | 20.9 ns | 19.0 | 7.73 | 1.25 |
| Kernel thickness | 156.9 ** | 138.3** | 9.7 ** | 7.3 | 4.16 | 0.58 |
| Ear grain percentage | 56.6 ** | 223.1 ** | 14.4 ns | 14.7 | 87.30 | 4.70 |
| Weight of 100 kernels | 248.1 ** | 1114.0 ** | 22.2 ns | 18.8 | 27.79 | 7.81 |
| Volume of 100 kernels | 811.2 ** | 2990.6 ** | 50.1 ns | 47.1 | 42.86 | 13.52 |
| Kernel width/kernel length | 0.1 ** | 0.1 ** | 0.0 ** | 0.0 | 0.62 | 0.11 |
| Kernel thickness/kernel length | 0.0 ** | 0.1 ** | 0.0 ns | 0.0 | 0.33 | 0.07 |
| Weight of 100 kernels/volume of 100 kernels | 0.0 ** | 0.0 ns | 0.0 ns | 0.0 | 0.66 | 0.06 |
| Trait | σ2g | σ2l | σ2g×l | σ2e | H2 | r |
|---|---|---|---|---|---|---|
| Female flowering † | 148.84 | 108.4238 | 12.9696 | 13.488 | 84.91 | 1.2261 |
| Male flowering | 130.024 | 143.2363 | 8.6491 | 15.734 | 84.21 | 0.8561 |
| Anthesis-silking interval | 3.935 | 2.7019 | 2.1987 | 5.467 | 33.92 | 0.8030 |
| Shoots per plant | 0.001 | 0.0618 | 0.0098 | 0.052 | 1.59 | 0.0143 |
| Plant height | 1687.46 | 1734.8050 | 21.3440 | 288.216 | 84.50 | 0.9609 |
| Ear height | 864.46 | 1346.0031 | 24.0306 | 206.367 | 78.96 | 0.6310 |
| Ear height/plant height | 0.002 | 0.0050 | 0.0005 | 0.002 | 44.44 | 0.3706 |
| Number of leaves † | 3.388 | 0.0830 | 0.2757 | 0.6 | 79.46 | 9.4447 |
| Number of leaves above ear † | 0.325 | 0.0421 | 0.0193 | 0.108 | 71.85 | 5.3000 |
| Leaf length | 56.02 | 237.4093 | 14.1348 | 29.239 | 56.36 | 0.2227 |
| Leaf width | 0.985 | 3.9453 | 0.1478 | 1.439 | 38.30 | 0.2408 |
| Total length † | 20.883 | 0.2329 | 2.2085 | 25.596 | 42.89 | 8.5538 |
| Peduncle length of the tassel † | 2.949 | 0.8468 | 0.8712 | 11.149 | 19.70 | 1.7169 |
| Length of central spike † | 9.012 | 0.9987 | 1.4356 | 7.69 | 49.69 | 3.7021 |
| Length of branched part of tassel | 3.538 | 4.9074 | 0.1090 | 2.201 | 60.50 | 0.7055 |
| Length of branched part of tassel/total length | 0.0008 | 0.0011 | 0.0001 | 0.001 | 42.11 | 0.6358 |
| Number of primary branches | 5.335 | 4.8565 | 0.7163 | 4.79 | 49.21 | 0.9574 |
| Ear length † | 5.316 | 1.7033 | 0.1233 | 2.696 | 65.34 | 2.9107 |
| Number of kernel rows † | 3.932 | 0.7603 | 0.1303 | 1.044 | 77.00 | 4.4151 |
| Peduncle length of the ear | 2.923 | 6.3434 | 1.0325 | 6.46 | 28.06 | 0.3963 |
| Ear diameter | 0.149 | 0.1415 | 0.0111 | 0.054 | 69.59 | 0.9795 |
| Ear diameter/Ear length † | 0.001 | 0.0001 | 0.0001 | 0.001 | 47.62 | 8.0939 |
| Cob diameter † | 0.085 | 0.0506 | 0.0093 | 0.074 | 50.51 | 1.4316 |
| Kernel length † | 114.213 | 60.0425 | 16.8775 | 64.056 | 58.53 | 1.4848 |
| Kernel width † | 134.291 | 6.1751 | 0.9700 | 18.99 | 87.06 | 18.7950 |
| Kernel thickness † | 24.545 | 0.5327 | 1.1700 | 7.33 | 74.28 | 14.4151 |
| Ear grain percentage † | 7.025 | 0.9063 | −0.1294 | 14.699 | 32.53 | 9.0428 |
| Weight of 100 kernels † | 37.655 | 4.4013 | 1.733 | 18.755 | 64.76 | 6.1385 |
| Volume of 100 kernels † | 126.85 | 12.1132 | 1.5034 | 47.084 | 72.30 | 9.3161 |
| Kernel width/kernel length † | 0.008 | 0.0005 | 0.0005 | 0.003 | 69.57 | 9.2262 |
| Kernel thickness/kernel length † | 0.002 | 0.0005 | 0.0002 | 0.002 | 47.62 | 3.1202 |
| Weight of 100 kernel/volume of 100 kernels † | 0.001 | 0.0001 | 0.0001 | 0.002 | 32.26 | 10.2235 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocandio-Rodríguez, M.; Santacruz-Varela, A.; Córdova-Téllez, L.; López-Sánchez, H.; Hernández-Bautista, A.; Castillo-González, F.; Lobato-Ortiz, R.; García-Zavala, J.J.; Antonio López, P. Estimation of Genetic Diversity in Seven Races of Native Maize from the Highlands of Mexico. Agronomy 2020, 10, 309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020309
Rocandio-Rodríguez M, Santacruz-Varela A, Córdova-Téllez L, López-Sánchez H, Hernández-Bautista A, Castillo-González F, Lobato-Ortiz R, García-Zavala JJ, Antonio López P. Estimation of Genetic Diversity in Seven Races of Native Maize from the Highlands of Mexico. Agronomy. 2020; 10(2):309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020309
Chicago/Turabian StyleRocandio-Rodríguez, Mario, Amalio Santacruz-Varela, Leobigildo Córdova-Téllez, Higinio López-Sánchez, Aurelio Hernández-Bautista, Fernando Castillo-González, Ricardo Lobato-Ortiz, J. Jesús García-Zavala, and Pedro Antonio López. 2020. "Estimation of Genetic Diversity in Seven Races of Native Maize from the Highlands of Mexico" Agronomy 10, no. 2: 309. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10020309