Hybridization in Peppers (Capsicum spp.) to Improve the Volatile Composition in Fully Ripe Fruits: The Effects of Parent Combinations and Fruit Tissues

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Samples and Extraction of Volatiles
2.3. Extraction of the Volatile Fraction
2.4. Analysis of Volatiles
2.5. Data Analysis
3. Results and Discussion
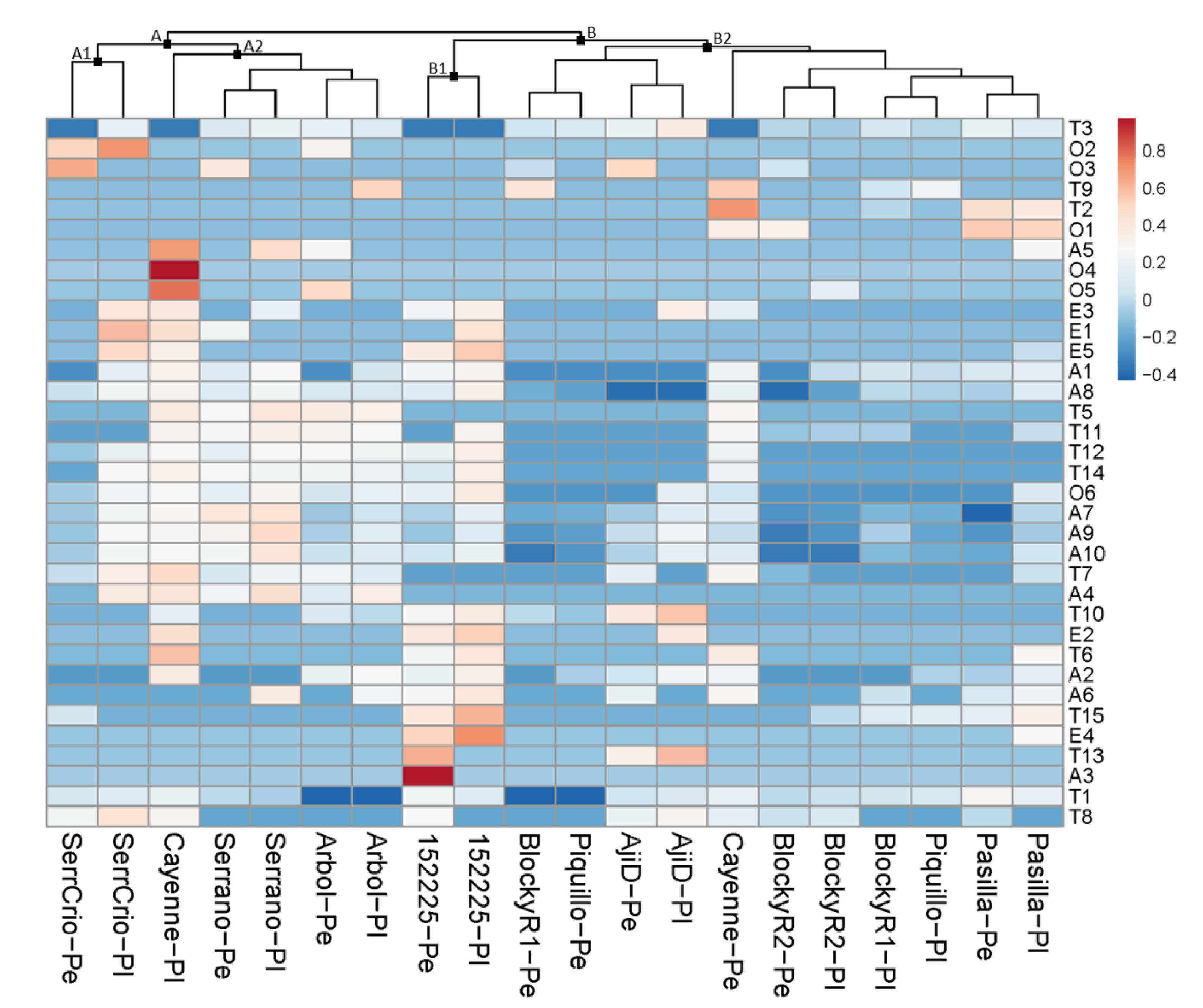
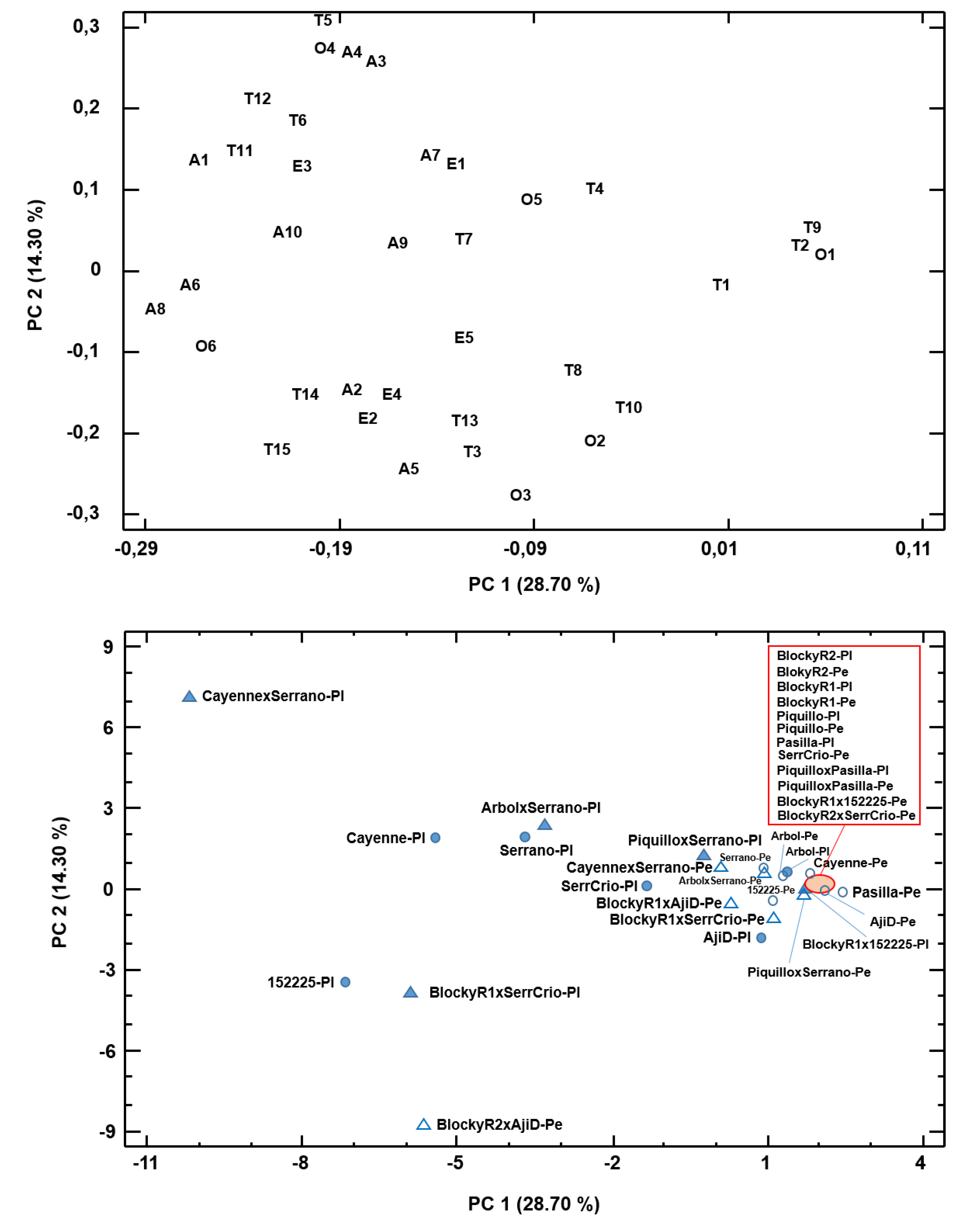
3.1. Volatile Fraction in the Pericarp
3.1.1. Parent Accessions
3.1.2. Hybrid Accessions
3.2. Volatile Fraction in Placental Tissues
3.2.1. Parent Accessions
3.2.2. Hybrid Accessions
3.3. Comparative Accumulation of Volatiles in Fruit Tissues
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garcés-Claver, A.; Arnedo-Andrés, M.S.; Abadía, J.; Gil-Ortega, R.; Álvarez-Fernández, A. Determination of capsaicin and dihydrocapsaicin in Capsicum fruits by liquid chromatography-electrospray/time-of-flight mass spectrometry. J. Agric. Food Chem. 2006, 54, 9303–9311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Castilla, L.C.; Garruña-Hernández, R.; Castillo-Aguilar, C.C.; Martínez-Hernández, A.; Ortíz-García, M.M.; Andueza-Noh, R.H. Structure and Genetic Diversity of Nine Important Landraces of Capsicum Species Cultivated in the Yucatan Peninsula, Mexico. Agronomy 2019, 9, 376. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Dias, L.; Vilanova, S.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Genetic diversity, population structure, and relationships in a collection of pepper (Capsicum spp.) landraces from the Spanish centre of diversity revealed by genotyping-by-sequencing (GBS). Hortic. Res. 2019, 6, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, K.; Ruiz, C.; Calderon, R.; Marcelo, M.; Rojas, R. Characterisation of volatile profiles in 50 native Peruvian chili pepper using solid phase microextraction-gas chromatography mass spectrometry (SPME-GCMS). Food Res. Int. 2016, 89, 471–475. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, D.; Bosland, P.W. The Complete Chile Pepper Book; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Bosland, P.W.; Votava, E. Peppers: Vegetable and Spice Capsicums. CABI Publishing: New York, NY, USA, 2000. [Google Scholar]
- Ribes-Moya, A.M.; Raigón, M.D.; Moreno-Peris, E.; Fita, A.; Rodríguez-Burruezo, A. Response to organic cultivation of heirloom Capsicum peppers: Variation in the level of bioactive compounds and effect of ripening. PLoS ONE 2018, 13, e0207888. [Google Scholar] [CrossRef] [Green Version]
- Pino, J.; González, M.; Ceballos, L.; Centurión-Yah, A.R.; Trujillo-Aguirre, J.; Latourniere-Moreno, L.; Sauri-Duch, E. Characterization of total capsaicinoids, colour and volatile compounds of Habanero chilli pepper (Capsicum chinense Jack.) cultivars grown in Yucatan. Food Chem. 2007, 104, 1682–1686. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; Kollmannsberger, H.; González-Mas, M.C.; Nitz, S.; Nuez, F. HS-SPME comparative analysis of genotypic diversity in volatile fraction and aroma contributing compounds of Capsicum fruits from the annuum–chinense–frutescens complex. J. Agric. Food Chem. 2010, 58, 4388–4400. [Google Scholar] [CrossRef]
- Bogusz, S., Jr.; Teixeira-Godoy, H.; Marchi-Tavares, A.; Teixeira, J. Analysis of the volatile compounds of Brazilian chilli peppers (Capsicum spp.) at two stages of maturity by solid phase micro-extraction and gas chromatography-mass spectrometry. Food Res. Int. 2012, 48, 98–107. [Google Scholar] [CrossRef]
- Morales-Soriano, E.; Kebede, B.T.; Ugás, R.; Grauwet, T.; Van Loey, A.; Hendrickx, M. Flavor characterization of native Peruvian chili peppers through integrated aroma fingerprinting and pungency profiling. Food Res. Int. 2018, 109, 250–259. [Google Scholar] [CrossRef]
- Buczkowska, H.; Nurzynska-Wierdak, R.; Labuda, H.; Salata, A. Relationships of the capsaicinoid content between the fruit parts of hot pepper (Capsicum annuum L.). Acta Sci. Pol. Hortorum Cultus. 2016, 15, 185–198. [Google Scholar]
- Olguín-Rojas, J.A.; Fayos, O.; Vázquez-León, L.A.; Ferreiro-González, M.; Rodríguez-Jimenes, G.C.; Palma, M.; Garcés-Claver, A.; Barbero, G.F. Progression of the Total and Individual Capsaicinoids Content in the Fruits of Three Different Cultivars of Capsicum chinense Jacq. Agronomy 2019, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Baby, K.C.; Ranganathan, T.V. Effect of enzyme pretreatment on yield and quality of fresh green chilli (Capsicum annuum L.) oleoresin and its major capsaicinoids. Biocatal. Agric. Biotechnol. 2016, 7, 95–101. [Google Scholar] [CrossRef]
- Barchenger, D.W.; Bosland, P.W. Exogenous applications of capsaicin inhibits seed germination of Capsicum Annuum. Sci. Hortic. 2016, 203, 29–31. [Google Scholar] [CrossRef] [Green Version]
- Scoville, W.L. Note on Capsicum. J. Am. Pharm. Assoc. 1912, 1, 453–454. [Google Scholar] [CrossRef] [Green Version]
- Estrada, B.; Pomar, F.; Díaz, J.; Merino, F.; Bernal, M.A. Pungency level in fruits of the Padrón pepper with different water supply. Sci. Hort. 1999, 81, 385–396. [Google Scholar] [CrossRef]
- Jarret, R.L.; Perkins, B.; Fan, T.; Prince, A.; Guthrie, K.; Skoczenski, B. Using EIA to screen Capsicum spp. Germplasm for capsaicinoid content. J. Food Comp. Anal. 2003, 16, 189–194. [Google Scholar] [CrossRef]
- Wahyuni, Y.; Ballester, A.R.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 2013, 76, 783–793. [Google Scholar] [CrossRef]
- Lu, M.; Ho, C.T.; Huang, Q. Extraction, bioavailability, and bioefficacy of capsaicinoids. J. Food Drug Anal. 2017, 25, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Kollmannsberger, H.; Rodríguez-Burruezo, A.; Nitz, S.; Nuez, F. Volatile and capsaicinoid composition of ají (Capsicum baccatum) and rocoto (Capsicum pubescens), two Andean species of chile peppers. J. Sci. Food Agric. 2011, 91, 1598–1611. [Google Scholar] [CrossRef]
- Eggink, P.M.; Tikunov, Y.; Maliepaard, C.; Haanstra, J.P.; de Rooij, H.; Vogelaar, A.; Gutteling, E.W.; Freymark, G.; Bovy, A.G.; Visser, R.G. Capturing flavors from Capsicum baccatum by introgression in sweet pepper. Theor. Appl. Genet. 2014, 127, 373–390. [Google Scholar] [CrossRef]
- Luning, P.A.; de Rijk, T.; Wichers, H.J.; Roozen, J.P. Gas chromatography, mass spectrometry, and sniffing port analyses of volatile compounds of fresh bell peppers (Capsicum annuum) at different ripening stages. J. Agric. Food Chem. 1994, 42, 977–983. [Google Scholar] [CrossRef]
- Cremer, D.R.; Eichner, K. Formation of volatile compounds during heating of spice paprika (Capsicum annuum) powder. J. Agric. Food Chem. 2000, 48, 2454–2460. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.; Arrowsmith, N.; Gladis, T. Agrobiodiversity with emphasis on plant genetic resources. Naturwissenschaften 2003, 90, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Brugarolas, M.; Martinez-Carrasco, L.; Martinez-Poveda, A.; Ruiz, J.J. A competitive strategy for vegetable products: Traditional varieties of tomato in the local market. Span. J. Agric. Res. 2009, 7, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Gancel, A.L.; Ollitrault, P.; Froelicher, Y.; Tomi, F.; Jacquemond, C.; Luro, F.; Brillouet, J.M. Leaf volatile compounds of six citrus somatic allotetraploid hybrids originating from various combinations of lime, lemon, citron, sweet orange, and grapefruit. J. Agric. Food Chem. 2005, 53, 2224–2230. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; Kollmannsberger, H.; Prohens, J.; Nitz, S.; Nuez, F. Analysis of the volatile aroma constituents of parental and hybrid clones of pepino (Solanum muricatum). J. Agric. Food Chem. 2004, 52, 5663–5669. [Google Scholar] [CrossRef]
- Antonio, A.S.; Wiedemann, L.S.M.; Veiga-Junior, V.F. The genus Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv. 2018, 8, 25767–25784. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.; Schieberle, P. Important odorants of sweet bell pepper powder (Capsicum annuum cv annuum): Differences between samples of Hungarian and Morrocan origin. Eur. Food Res. Technol. 2000, 211, 175–180. [Google Scholar] [CrossRef]
- Simian, H.; Robert, F.; Blank, I. Identification and synthesis of 2-heptanethiol, a new flavor compound found in bell peppers. J. Agric. Food Chem. 2004, 52, 306–310. [Google Scholar] [CrossRef]
- Sosa-Moguel, O.; Pino, J.A.; Ayora-Talavera, G.; Sauri-Duch, E.; Cuevas-Glory, L. Biological activities of volatile extracts from two varieties of Habanero pepper (Capsicum chinense Jacq.). Int. J. Food Prop. 2017, 20 (Suppl. 3), S3042–S3051. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, G.; Lo Scalzo, R. Characterization of hot pepper spice phytochemicals, taste compounds content and volatile profiles in relation to the drying temperature. J. Food Biochem. 2018, 42, e12675. [Google Scholar] [CrossRef]
- Moreno, E.; Fita, A.; González-Mas, M.C.; Rodríguez-Burruezo, A. HS-SPME study of the volatile fraction of Capsicum accessions and hybrids in different parts of the fruit. Sci. Hort. 2012, 135, 87–97. [Google Scholar] [CrossRef]
- Gómez, E.; Ledbetter, C.A.; Hartsell, P.L. Volatile compounds in apricot, plum, and their interspecific hybrids. J. Agric. Food Chem. 1993, 41, 1669–1676. [Google Scholar] [CrossRef]
- Gancel, A.L.; Ollitrault, P.; Froelicher, Y.; Tomi, F.; Jacquemond, C.; Luro, F.; Brillouet, J.M. Leaf volatile compounds of seven citrus somatic tetraploid hybrids sharing willow leaf mandarin (Citrus deliciosa Ten.) as their common parent. J. Agric. Food Chem. 2003, 51, 6006–6013. [Google Scholar] [CrossRef]
- Nuez, F.; Gil, R.; Costa, J. El Cultivo de Pimientos, Chiles y Ajíes (The Crop of Peppers, Chilies and Ajíes); MundiPrensa: Madrid, Spain, 2003. (In Spanish) [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Cuevas-Glory, L.F.; Sosa-Moguel, O.; Pino, J.; Sauri-Duch, E. GC–MS characterization of volatile compounds in habanero pepper (Capsicum chinense Jacq.) by optimization of headspace solid-phase microextraction conditions. Food Anal. Method. 2015, 8, 1005–1013. [Google Scholar] [CrossRef]
- Orzáez, D.; Granell, A. Reverse genetics and transient gene expression in fleshy fruits: Overcoming plant stable transformation. Plant Signal. Behav. 2009, 4, 864–867. [Google Scholar] [CrossRef]
- Pinheiro, T.T.; Peres, L.E.P.; Purgatto, E.; Latado, R.R.; Maniero, R.A.; Martins, M.M.; Figueira, A. Citrus carotenoid isomerase gene characterization by complementation of the “Micro-Tom” tangerine mutant. Plant Cell Rep. 2019, 38, 623–636. [Google Scholar] [CrossRef]
- Rothan, C.; Diouf, I.; Causse, M. Trait discovery and editing in tomato. Plant J. 2019, 97, 73–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulet, B.E.; Roda, F.; Hopkins, R. Hybridization in plants: Old ideas, new techniques. Plant Physiol. 2017, 173, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambla, J.L.; Tikunov, Y.M.; Monforte, A.J.; Bovy, A.G.; Granell, A. The expanded tomato fruit volatile landscape. J. Exp. Bot. 2014, 65, 4613–4623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubert, C.; Milhet, C. Distribution of the volatile compounds in the different parts of a white-fleshed peach (Prunus persica L. Batsch). Food Chem. 2007, 102, 375–384. [Google Scholar] [CrossRef]
- Moing, A.; Aharoni, A.; Biais, B.; Rogachev, I.; Meir, S.; Brodsky, L.; Allwood, J.W.; Erban, A.; Dunn, W.B.; Kay, L.; et al. Extensive metabolic cross-talk in melon fruit revealed by spatial and developmental combinatorial metabolomics. New Phytol. 2011, 190, 683–696. [Google Scholar] [CrossRef]
- Deyeye, E.; Aremu, M. Chemical composition of theraw fruit coat, seed and pulp of passion fruit (Passiflora edulis). J. Trends Sci. Tech. 2017, 2, 334–341. [Google Scholar]
- Wang, L.; Qian, C.; Bai, J.; Luo, W.; Jin, C.; Yu, Z. Difference in volatile composition between the pericarp tissue and inner tissue of tomato (Solanum lycopersicum) fruit. J. Food Process. Preserv. 2018, 42, e13387. [Google Scholar] [CrossRef]
- Dardick, C.; Callahan, A.M. Evolution of the fruit endocarp: Molecular mechanisms underlying adaptations in seed protection and dispersal strategies. Front Plant Sci. 2014, 5, 284. [Google Scholar] [CrossRef]
- Bosland, P.; Coon, D.; Cooke, P.H. Novel formation of ectopic (nonplacental capsaicinoid secreting vesicles on fruit walls explains the morphological mechanism for super-hot chile peppers. J. Amer. Soc. Hort. Sci. 2015, 140, 253–256. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Accession (Abbreviation) | Varietal/Pod Type | Origin | Weight (g) | Length/Width (mm) | Aroma Quality (Intensity) 1 |
|---|---|---|---|---|---|
| C. annuum | |||||
| Blocky Red1 (BlockyR1) | Bell/large | Breeding line | 200–300 | 121/105 | Paprika (low) |
| Blocky Red2 (BlockyR2) | Bell/intermediate | Breeding line | 150–200 | 106/93 | Paprika (low) |
| Piquillo (Piquillo) | Piquillo/Ancho | PGI Piquillo Lodosa (Spain) | 25–50 | 90/50 | Paprika, green (intermediate) |
| Chile de Arbol (Arbol) | Cayenne, small | Mexico/south USA | 5–10 | 71/11 | Paprika, green, fruity notes (high) |
| Guindilla Española (Cayenne) | Cayenne, intermediate | Valencia (Spain) | 5–10 | 108/12 | Fruity, green paprika notes (high) |
| Pasilla Bajío (Pasilla) | Cayenne, large | Mexico/south USA | 10–25 | 204/29 | Green, fruity notes (intermediate) |
| Chile Serrano (Serrano) | Serrano | Mexico/south USA | 5–10 | 35/16 | Green paprika, fruity citrus notes (high) |
| Serrano Criollo (SerrCrio) | Serrano-Cayenne | Mexico | <5 | 39/12 | Green paprika (interm.-high) |
| C. chinense | |||||
| PI-152225 (152225) | Cayenne | USDA | <5 | 41/12 | Fruity, sweet, exotic (high) |
| Ají Dulce (Ají D) | Flattened (sweet) | Cuba | 5–10 | 20/31 | Fruity, sweet (interm.-high) |
| C. annuum × C. annuum (abbreviation) | C. annuum × C. chinense (abbreviation) | ||||
| Blocky Red1×Serrano Criollo (BlockyR1×SerrCrio) | Blocky Red1×PI-152225 (BlockyR1×152225) | ||||
| Blocky Red2×Serrano Criollo (BlockyR2×SerrCrio) | Blocky Red1×Ají Dulce (BlockyR1×AjiD) | ||||
| Piquillo×Pasilla Bajío (Piquillo×Pasilla) | Blocky Red2×Ají Dulce (BlockyR2×AjiD) | ||||
| Piquillo×Chile Serrano (Piquillo×Serrano) | |||||
| Chile de Arbol×Chile Serrano (Arbol×Serrano) | |||||
| Guindilla Española×Chile Serrano (Cayenne×Serrano) | |||||
| Volatile Compound | Volatile Abbreviation | RI | Id 1 |
|---|---|---|---|
| Terpenoids | |||
| 3-Carene | T1 | 948 | R |
| Cyclosativene | T2 | 1125 | R |
| α-Copaene | T3 | 1221 | R |
| β-Cubebene | T4 | 1339 | MS |
| Allo-Aromadendrene | T5 | 1386 | MS |
| β-Elemene | T6 | 1398 | MS |
| α-Longipinene | T7 | 1403 | R |
| α-Muurolene | T8 | 1440 | MS |
| β-Selinene | T9 | 1469 | MS |
| δ-Cadinene | T10 | 1469 | MS |
| Valencene | T11 | 1474 | R |
| α-Himachalene | T12 | 1494 | MS |
| β-Chamigrene | T13 | 1507 | R |
| β-Himachalene | T14 | 1528 | MS |
| Longipinocarvone | T15 | 1569 | MS |
| Esters | |||
| Hexyl 2-methylpropanoate | E1 | 1118 | R |
| Pentyl 4-methylpentanoate | E2 | 1118 | R |
| Hexyl 2,2-dimethylpropanoate | E3 | 1197 | R |
| Hexyl 2-methylbutanoate | E4 | 1218 | R |
| 4-Hexen-1-yl-pentanoate | E5 | 1290 | R |
| Alkanes | |||
| Tetradecane | A1 | 1413 | R |
| 3-Methyltetradecane | A2 | 1448 | MS |
| 4-Methyltetradecane | A3 | 1448 | MS |
| Decylcyclopentane | A4 | 1555 | MS |
| Cyclotetradecane | A5 | 1679 | MS |
| Cyclopentadecane | A6 | 1799 | MS |
| Nonadecane | A7 | 1910 | R |
| 10-Methylnonadecane | A8 | 1945 | MS |
| Eicosane | A9 | 2009 | R |
| 8-Hexylpentadecane | A10 | 2045 | MS |
| Miscellany/Others | |||
| Indole | O1 | 1174 | R |
| 3-Isobutyl-2-methoxypyrazine | O2 | 1204 | R |
| Methyl salicylate | O3 | 1281 | R |
| α-Ionone | O4 | 1429 | R |
| (E)-2-hexadecen-1-ol | O5 | 1862 | MS |
| (Z)-9-octadecen-1-ol | O6 | 2061 | R |
| C. annuum | C. chinense | C. annuum × C. annuum | C. annuum × C. chinense | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | BlockyR1 | BlockyR2 | Piquillo | Arbol | Cayenne | Pasilla | Serrano | SerrCrio | 152225 | AjiD | BlockyR1 ×SerrCrio | BlockyR2 ×SerrCrio | Piquillo ×Pasilla | Piquillo ×Serrano | Arbol ×Serrano | Cayenne ×Serrano | BlockyR1 ×152225 | BlockyR1 ×AjiD | BlockyR2 ×AjiD |
| Terpenoids | |||||||||||||||||||
| T1 | -- | 2.75a 1 | -- | -- | 36.50c | 188.38e | 2.29a | 6.94a | 99.45d | 4.58a | 14.44b | -- | -- | -- | -- | -- | -- | 1.67a | 20.98b |
| T2 | -- | -- | -- | -- | 4.04 | 0.53 | -- | -- | -- | -- | 1.08 | -- | 1.37 | -- | -- | -- | tr | -- | -- |
| T3 | 0.96a | 0.44a | 1.76a | 6.53b | -- | 7.12b | 1.88a | -- | -- | 8.13b | 4.33ab | 0.57a | 2.25a | 3.75ab | 4.30ab | 6.18b | 0.62a | 9.96b | 112.29c |
| T5 | -- | -- | -- | 2.41bc | 0.98ab | -- | 0.68ab | -- | -- | -- | -- | -- | -- | 0.40a | 2.07b | 3.34c | 0.01a | -- | -- |
| T6 | -- | -- | -- | -- | 2.00b | -- | -- | -- | 0.42a | -- | 0.30a | 0.30a | 0.04a | 0.21a | -- | 5.40c | -- | -- | -- |
| T7 | -- | Tr 2 | -- | 0.26ab | 0.54b | -- | 0.06a | 0.04a | tr | 0.13a | 3.05c | 0.03a | tr | 0.04a | 0.16a | 0.20a | -- | 0.67b | tr |
| T8 | -- | 0.25a | -- | tr | 2.43b | 0.18a | tr | 9.74c | -- | 5.55bc | 23.62d | 8.49c | -- | tr | tr | tr | -- | 21.32d | 164.00e |
| T9 | 0.17a | -- | -- | -- | 0.41a | -- | -- | -- | -- | -- | -- | -- | 0.02a | -- | -- | -- | -- | -- | -- |
| T10 | 0.04a | -- | 0.01a | 0.12a | -- | -- | -- | -- | 0.83a | 5.19b | -- | -- | -- | -- | 0.11a | -- | -- | 7.63b | 18.42c |
| T11 | -- | 0.06a | -- | 205.86 | 82.62c | -- | 68.65c | -- | -- | -- | 1.66a | tr | 0.05a | 33.19b | 182.01d | 285.88e | -- | -- | 34.60b |
| T12 | -- | -- | -- | 3.12c | 1.39b | -- | 0.82ab | 0.02a | 1.08b | -- | -- | -- | -- | 0.56ab | 2.70c | 3.31c | 0.01a | -- | -- |
| T13 | -- | -- | -- | tr | -- | -- | -- | -- | 3.60b | 0.10a | 8.32c | -- | -- | tr | tr | tr | tr | -- | 118.80d |
| T14 | -- | -- | -- | 7.08c | 5.01bc | -- | 12.21d | -- | 0.55a | 5.28bc | 3.53b | 0.24a | -- | 1.43ab | 6.73c | 8.13c | -- | 8.78c | 53.32e |
| T15 | -- | -- | -- | -- | -- | 0.09a | -- | 0.03a | 0.86ab | -- | 0.19a | -- | 0.05a | -- | -- | 0.81ab | -- | 1.61b | 8.49c |
| Total terpenoids | 1.17a | 3.50a | 1.77a | 225.38h | 135.92g | 196.3h | 86.59f | 16.77c | 106.79fg | 28.96cd | 60.50e | 9.63b | 3.78a | 39.58d | 198.1h | 313.25i | 0.64a | 51.64e | 530.9j |
| Esters | |||||||||||||||||||
| E1 | -- | -- | -- | -- | -- | -- | 0.41a | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| E2 | -- | -- | -- | -- | -- | -- | -- | -- | 16.57b | 14.06b | 0.37a | -- | -- | -- | -- | -- | -- | 1.07a | 57.68c |
| E3 | -- | -- | -- | -- | 1.90a | -- | -- | -- | 5.84b | -- | 0.60a | -- | -- | 0.62a | -- | 28.03c | -- | -- | 39.23d |
| E4 | -- | -- | -- | -- | -- | -- | -- | -- | 10.15b | -- | -- | -- | -- | -- | -- | 1.48a | -- | 1.25a | 29.00c |
| E5 | -- | -- | -- | -- | -- | -- | -- | -- | 5.49a | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| Total esters | -- | -- | -- | -- | 1.90a | -- | 0.41a | -- | 38.05c | 14.06b | 0.97a | -- | -- | 0.62a | -- | 29.51c | -- | 2.32a | 125.91d |
| Alkanes | |||||||||||||||||||
| A1 | -- | -- | -- | -- | 8.32c | 1.09a | 2.22b | -- | 8.79c | 6.99c | 1.04a | 0.36a | 0.40a | 1.14a | -- | 6.42c | -- | 9.24cd | 11.72d |
| A2 | -- | -- | 0.07a | 1.51ab | 2.53b | 0.07a | -- | -- | 1.66ab | 0.20a | 1.08ab | 1.23ab | 0.05a | 1.00ab | 5.21c | -- | -- | -- | 19.34d |
| A3 | -- | -- | -- | -- | -- | -- | -- | -- | 7.05a | -- | -- | -- | -- | 4.89a | -- | -- | -- | -- | tr |
| A4 | -- | -- | -- | 0.08a | -- | -- | 0.20a | -- | -- | -- | -- | 0.07a | -- | -- | 0.60b | -- | -- | 0.33ab | -- |
| A5 | -- | -- | -- | 0.13a | -- | -- | -- | -- | -- | -- | 0.22a | 0.27a | -- | -- | -- | -- | -- | 2.35b | 15.88c |
| A6 | -- | -- | -- | -- | 14.78c | 0.50a | -- | -- | 7.58b | 3.06ab | 1.18a | -- | 0.31a | 3.14ab | -- | 20.35d | -- | 40.50e | 70.90f |
| A7 | 0.08a | 0.03a | 0.09a | 0.28a | 2.10b | tr | 53.81e | 0.26a | 0.36a | 0.29a | 0.47a | 0.45a | 0.04a | 0.48a | 1.92b | 7.09c | 0.01a | 8.20cd | 10.68d |
| A8 | 0.16a | -- | 0.08a | 8.63c | 42.68e | 1.19a | 12.36d | 3.44b | 14.02d | -- | 5.37bc | 7.05c | 0.76a | 6.46c | 11.42d | 41.92e | 0.09a | 63.96f | 158.82g |
| A9 | 0.03a | 0.01a | 0.05a | 0.31a | 0.55a | 0.03a | 10.06d | 0.19a | 0.08a | 0.53a | 0.61a | 0.63a | 0.03a | 0.28a | 2.18b | 3.19b | 0.01a | 5.70c | 15.39e |
| A10 | -- | -- | 0.02a | 0.44a | 1.05ab | 0.03a | 4.79c | 0.16a | 0.49a | 0.23a | 0.69a | 0.39a | 0.05a | 0.35a | 1.90b | 3.74bc | tr | 4.47c | 14.75d |
| Total alkanes | 0.27a | 0.04a | 0.31a | 11.38c | 72.01f | 2.91ab | 83.44f | 4.05b | 40.03e | 11.30c | 10.66c | 10.45c | 1.64ab | 17.74d | 23.23d | 82.71f | 0.11a | 134.75g | 317.48h |
| Others | |||||||||||||||||||
| O1 | -- | 0.23a | -- | -- | 0.31a | 1.75b | -- | -- | -- | -- | -- | 0.12a | -- | -- | -- | -- | -- | -- | -- |
| O2 | -- | -- | -- | 0.04a | -- | -- | -- | 0.12a | -- | -- | 1.59b | tr | -- | 0.51ab | 0.12a | tr | 0.12a | -- | 1.14b |
| O3 | 0.02a | 0.02a | -- | -- | -- | -- | 0.59a | 6.28c | -- | 1.60ab | -- | 3.11b | -- | -- | 5.43c | tr | 0.31a | 2.94b | 10.63d |
| O4 | -- | -- | -- | -- | tr | -- | -- | tr | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| O5 | -- | -- | -- | 3.18a | -- | -- | -- | -- | -- | -- | -- | 1.89a | -- | -- | 9.53b | -- | -- | -- | -- |
| O6 | -- | -- | -- | 0.28a | 0.23a | -- | 1.07b | 0.06a | 1.15b | -- | 0.41a | 0.55ab | 0.02a | 0.11a | 1.73b | 1.78b | 0.01a | 1.40b | 6.23c |
| Total volatiles | 1.46a | 3.79a | 2.08a | 242.34e | 210.37e | 200.96de | 172.1d | 27.28b | 186.02d | 55.92bc | 74.13c | 25.75b | 5.44a | 58.56bc | 237.91e | 427.25f | 1.19a | 193.05d | 992.29g |
| C. annuum | C. chinense | C. annuum × C. annuum | C. annuum × C. chinense | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | BlockyR1 | BlockyR2 | Piquillo | Arbol | Cayenne | Pasilla | Serrano | SerrCrio | 152225 | AjiD | BlockyR1 ×SerrCrio | Piquillo ×Pasilla | Piquillo ×Serrano | Arbol ×Serrano | Cayenne ×Serrano | BlockyR1 ×152225 |
| Terpenoids | ||||||||||||||||
| T1 | 5.56b 1 | 4.05b | 8.13c | -- | 51.73g | 37.50f | 1.42a | 15.84de | 15.20de | 11.98d | 17.07e | -- | 2.03a | -- | 2.94a | -- |
| T2 | Tr 2 | -- | tr | -- | -- | 0.29a | -- | -- | -- | -- | -- | 0.17a | -- | -- | -- | -- |
| T3 | 1.35ab | 0.28a | 0.47a | 2.67b | -- | 2.78b | 7.89c | 5.95c | -- | 68.88f | 14.38d | 3.18b | 8.50c | 12.55d | 25.28e | 0.44a |
| T4 | -- | -- | -- | tr | -- | -- | -- | -- | -- | -- | -- | -- | -- | 0.68 | -- | -- |
| T5 | -- | -- | -- | 1.29a | 2.79ab | -- | 3.94b | -- | -- | -- | -- | -- | 3.53b | 8.63c | 15.87d | -- |
| T6 | -- | -- | -- | -- | 27.14e | 0.87a | -- | -- | 3.21b | -- | 8.82d | 1.95b | 4.26c | 21.39e | 0.38a | |
| T7 | 0.01a | -- | -- | 0.11a | 2.94c | 0.04a | 0.22a | 0.83b | -- | -- | 2.02c | tr | 0.25a | 0.42ab | 0.78b | 0.05a |
| T8 | -- | 0.73a | -- | tr | 35.23d | tr | tr | 301.91e | 24.86c | 37.82d | -- | 4.93b | -- | -- | -- | 22.25c |
| T9 | 0.01a | -- | 0.05a | 0.31b | -- | -- | -- | -- | -- | -- | -- | 0.05a | -- | -- | -- | -- |
| T10 | -- | -- | -- | 0.04a | 0.27ab | -- | -- | -- | 4.24c | 39.26d | -- | -- | -- | 0.33ab | 0.68b | -- |
| T11 | 0.17a | 0.14a | -- | 110.69b | 253.97cd | 0.55a | 404.62e | -- | 229.25c | -- | 1297.48g | 0.08a | 297.91d | 730.40f | 1311.56g | -- |
| T12 | -- | -- | -- | 1.93ab | 3.69b | -- | 3.43b | 1.20ab | 9.48de | -- | -- | -- | 4.49c | 8.72d | 11.42e | 0.11a |
| T13 | -- | -- | -- | tr | -- | -- | -- | tr | -- | 0.89a | 422.36b | tr | -- | tr | -- | -- |
| T14 | -- | -- | -- | 4.57ab | 29.31e | tr | 8.27bc | 11.20c | 56.55f | -- | 174.29g | 1.65a | 10.40c | 22.26d | 28.97e | -- |
| T15 | 0.06a | 0.02a | 0.07a | -- | -- | 0.46b | -- | -- | 5.87d | -- | 2.59c | 0.21ab | -- | -- | 3.92c | 0.05a |
| Total terpenoids | 7.16a | 5.22a | 8.72ab | 121.61f | 407.07i | 42.49e | 429.79i | 336.93h | 348.66h | 158.83g | 1516.65k | 12.22b | 331.43h | 783.99j | 1422.81k | 23.28c |
| Esters | ||||||||||||||||
| E1 | -- | -- | -- | -- | 7.74b | -- | -- | 37.03d | 5.27b | -- | -- | -- | -- | 1.22a | 19.60c | 3.83ab |
| E2 | -- | -- | -- | -- | 43.09b | -- | -- | -- | 149.47c | -- | 5.75a | -- | -- | 8.85a | -- | -- |
| E3 | -- | -- | -- | -- | 117.49g | -- | 2.46b | 144.19h | 38.17e | 52.22f | 8.14c | -- | 0.98a | 15.15d | 142.80h | 8.52c |
| E4 | -- | -- | -- | -- | -- | 0.52a | -- | -- | 99.01c | -- | 0.91a | -- | -- | -- | 10.66b | 0.97a |
| E5 | -- | -- | -- | -- | 2.54c | 0.03a | -- | 22.91d | 49.46e | -- | -- | -- | -- | -- | -- | 0.55b |
| Total esters | -- | -- | -- | -- | 170.86f | 0.55a | 2.46b | 204.13f | 341.38g | 52.22e | 14.8c | -- | 0.98a | 25.22d | 173.06f | 13.87c |
| Alkanes | ||||||||||||||||
| A1 | 0.76a | 0.40a | 0.45a | 0.83a | 42.74 | 3.37b | 16.91d | 3.11b | 31.44e | -- | 3.19b | 0.97a | 6.48c | 5.67c | 43.58f | 0.38a |
| A2 | -- | -- | 0.09a | 4.55d | 19.27f | 0.86a | -- | -- | 9.64e | 2.33c | 45.81g | -- | -- | 24.75f | -- | 1.25b |
| A3 | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | 248.29a | -- |
| A4 | -- | -- | -- | 0.69b | 1.31c | -- | 2.01d | 0.86b | -- | -- | -- | 0.11a | 1.13bc | 3.19e | 2.40d | tr |
| A5 | -- | -- | -- | -- | 6.49d | 0.13a | 1.03b | -- | -- | -- | 3.90c | 0.11a | 0.78ab | 1.76b | -- | -- |
| A6 | 0.16a | -- | -- | 4.30b | -- | 3.94b | 64.53e | -- | 103.96f | -- | 33.51c | 4.46b | -- | 42.59d | 96.35f | 3.22b |
| A7 | 0.12a | 0.04a | 0.08a | 0.94a | 15.91e | 0.51a | 75.66g | 8.04c | 3.34b | 2.18b | 7.44c | 0.79a | 7.56c | 9.15d | 30.92f | 0.28a |
| A8 | 2.35b | 0.09a | 1.41a | 8.25c | 255.74g | 13.98d | 83.55f | 85.63f | 430.49i | -- | 321.72h | 8.32c | 57.01e | 51.91e | 272.00g | 7.31c |
| A9 | 0.31a | 0.03a | 0.06a | 1.85b | 5.68c | 0.27a | 62.84g | 6.05c | 1.66b | 4.30c | 22.36f | 0.80a | 5.07c | 8.64de | 12.12e | 0.39a |
| A10 | 0.07a | -- | 0.04a | 1.13ab | 8.10c | 0.45a | 36.74e | 4.97b | 2.84ab | 2.26ab | 14.58d | 1.42ab | 5.73b | 8.70c | 15.52d | 0.41a |
| Total alkanes | 3.77b | 0.56a | 2.13b | 22.54d | 355.24h | 23.51d | 343.27h | 108.66f | 583.37j | 11.07c | 452.51i | 16.98c | 83.74e | 156.36g | 721.18k | 13.24c |
| Others | ||||||||||||||||
| O1 | -- | -- | -- | -- | -- | 1.29a | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| O2 | -- | -- | -- | -- | -- | -- | tr | 0.33b | -- | -- | 0.62c | -- | 0.03a | tr | -- | 0.22ab |
| O4 | -- | -- | -- | -- | 0.83b | -- | -- | -- | -- | -- | -- | tr | -- | -- | 4.12c | 0.07a |
| O5 | -- | 0.07a | -- | -- | 80.61b | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- | -- |
| O6 | -- | -- | -- | 1.54b | 4.18d | 0.43a | 7.46e | 2.51c | 19.84g | 1.10ab | 14.51f | 1.33b | 5.32de | 6.76e | 6.44e | 0.84a |
| Total volatiles | 10.93b | 5.85a | 10.85b | 145.69e | 1018.79i | 68.27d | 782.98h | 652.56h | 1293.25j | 223.22f | 1999.09k | 30.53c | 421.5g | 972.33i | 2327.61l | 51.52d |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Peris, E.; Cortés-Olmos, C.; Díez-Díaz, M.; González-Mas, M.C.; de Luis-Margarit, A.; Fita, A.; Rodríguez-Burruezo, A. Hybridization in Peppers (Capsicum spp.) to Improve the Volatile Composition in Fully Ripe Fruits: The Effects of Parent Combinations and Fruit Tissues. Agronomy 2020, 10, 751. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050751
Moreno-Peris E, Cortés-Olmos C, Díez-Díaz M, González-Mas MC, de Luis-Margarit A, Fita A, Rodríguez-Burruezo A. Hybridization in Peppers (Capsicum spp.) to Improve the Volatile Composition in Fully Ripe Fruits: The Effects of Parent Combinations and Fruit Tissues. Agronomy. 2020; 10(5):751. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050751
Chicago/Turabian StyleMoreno-Peris, Estela, Carles Cortés-Olmos, Mónica Díez-Díaz, M. Carmen González-Mas, Ana de Luis-Margarit, Ana Fita, and Adrián Rodríguez-Burruezo. 2020. "Hybridization in Peppers (Capsicum spp.) to Improve the Volatile Composition in Fully Ripe Fruits: The Effects of Parent Combinations and Fruit Tissues" Agronomy 10, no. 5: 751. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10050751