
S-Like Ribonuclease T2 Genes Are Induced during Mobilisation of Nutrients in Cotyledons from Common Bean

 and
and 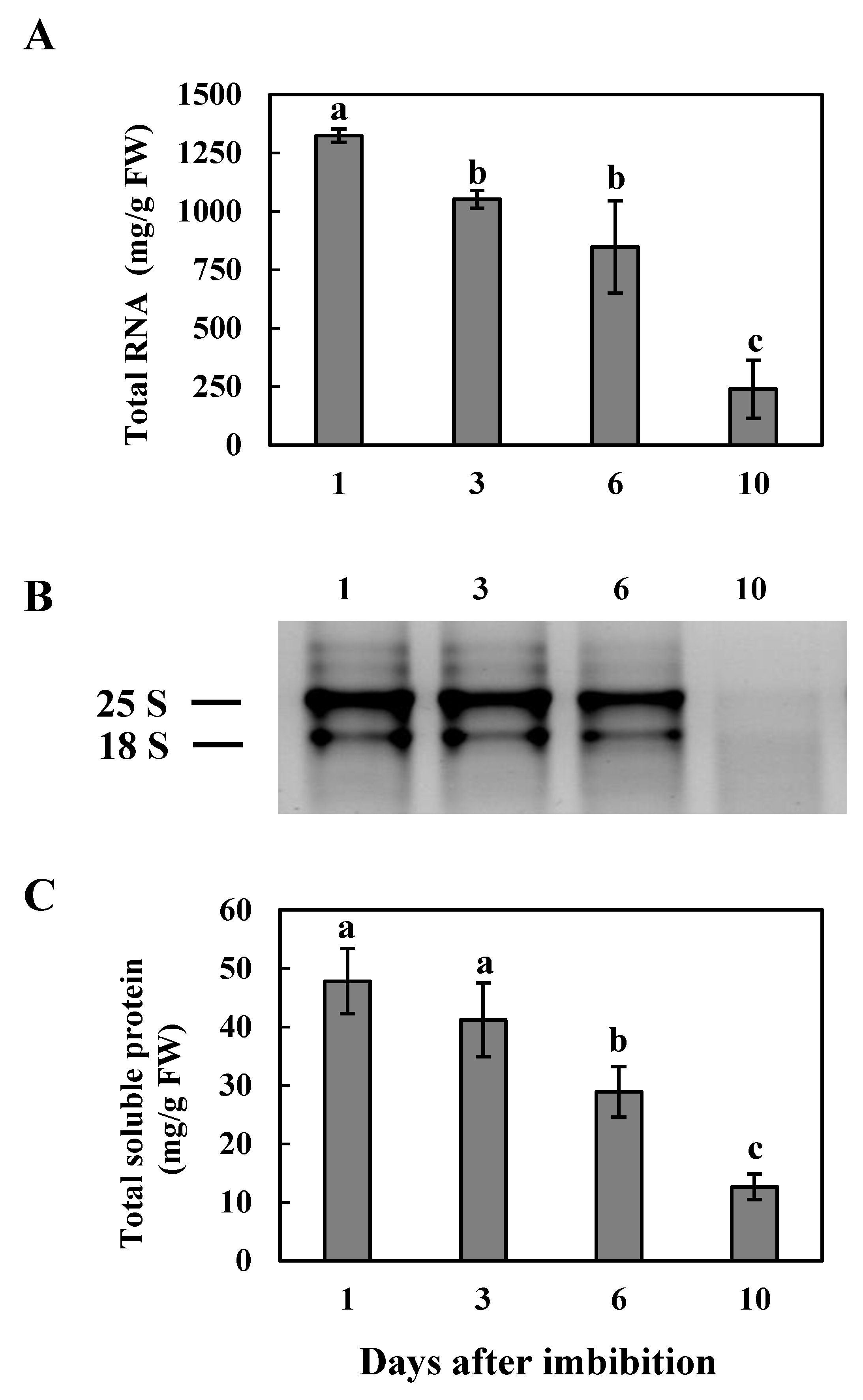{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Crude Extracts
2.3. Total Soluble Protein Determination
2.4. Determination of Enzymatic Activities
2.5. RNA Isolation
2.6. cDNA Synthesis
2.7. qRT
2.8. Statistics
3. Results
3.1. Total RNA and Protein during Cotyledon Development in Common Bean
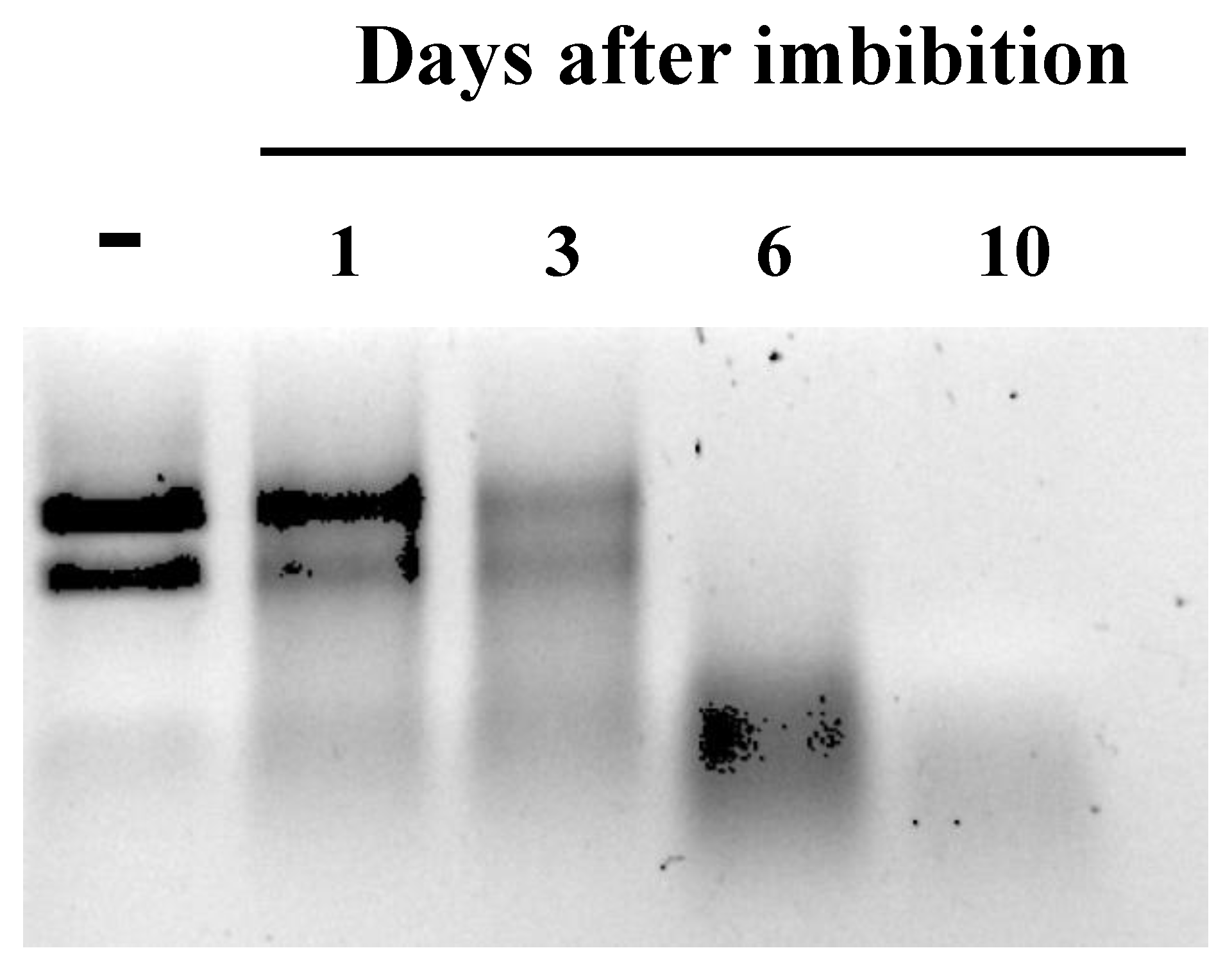
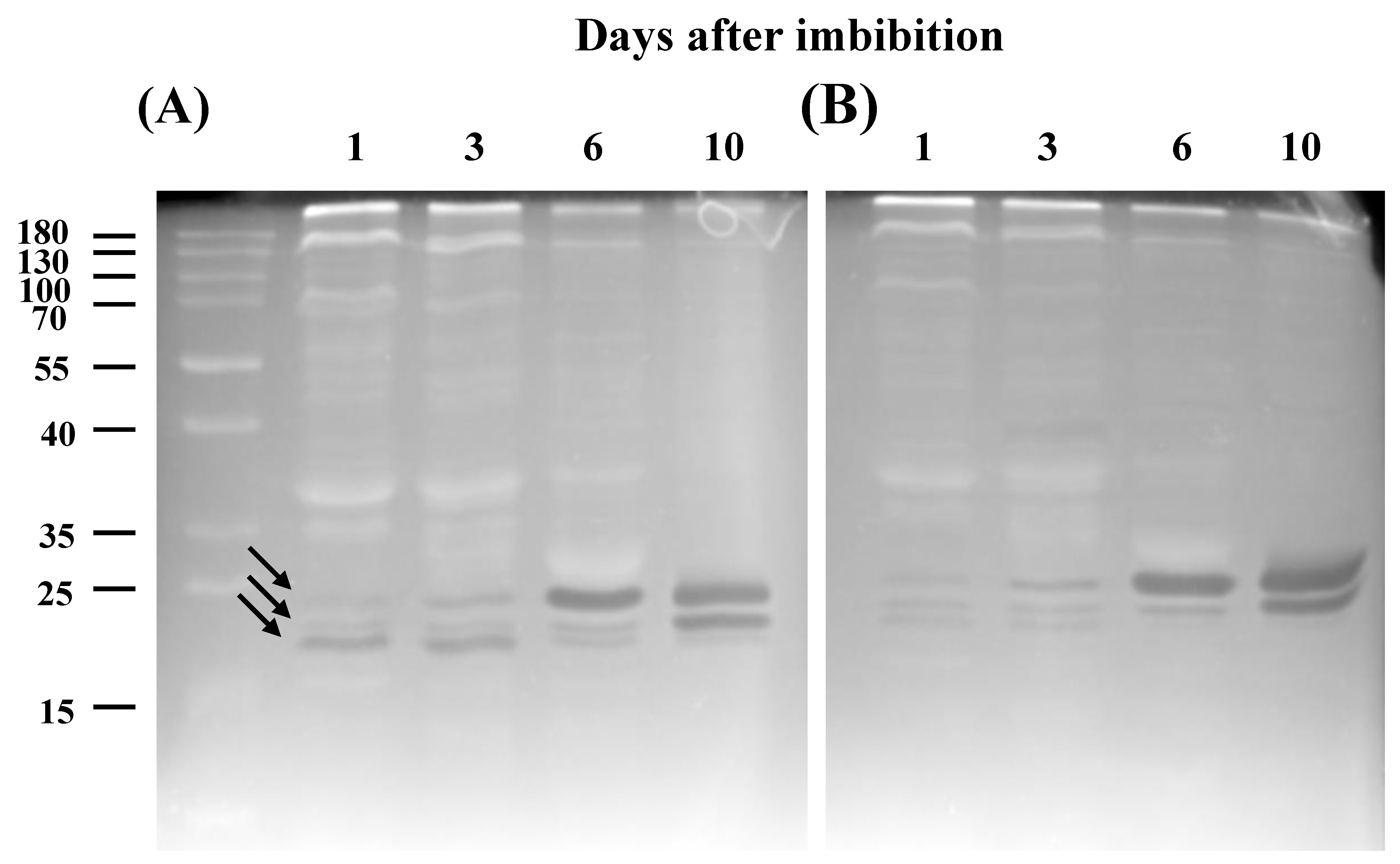
3.2. Ribonuclease Activities in Cotyledons of Common Bean
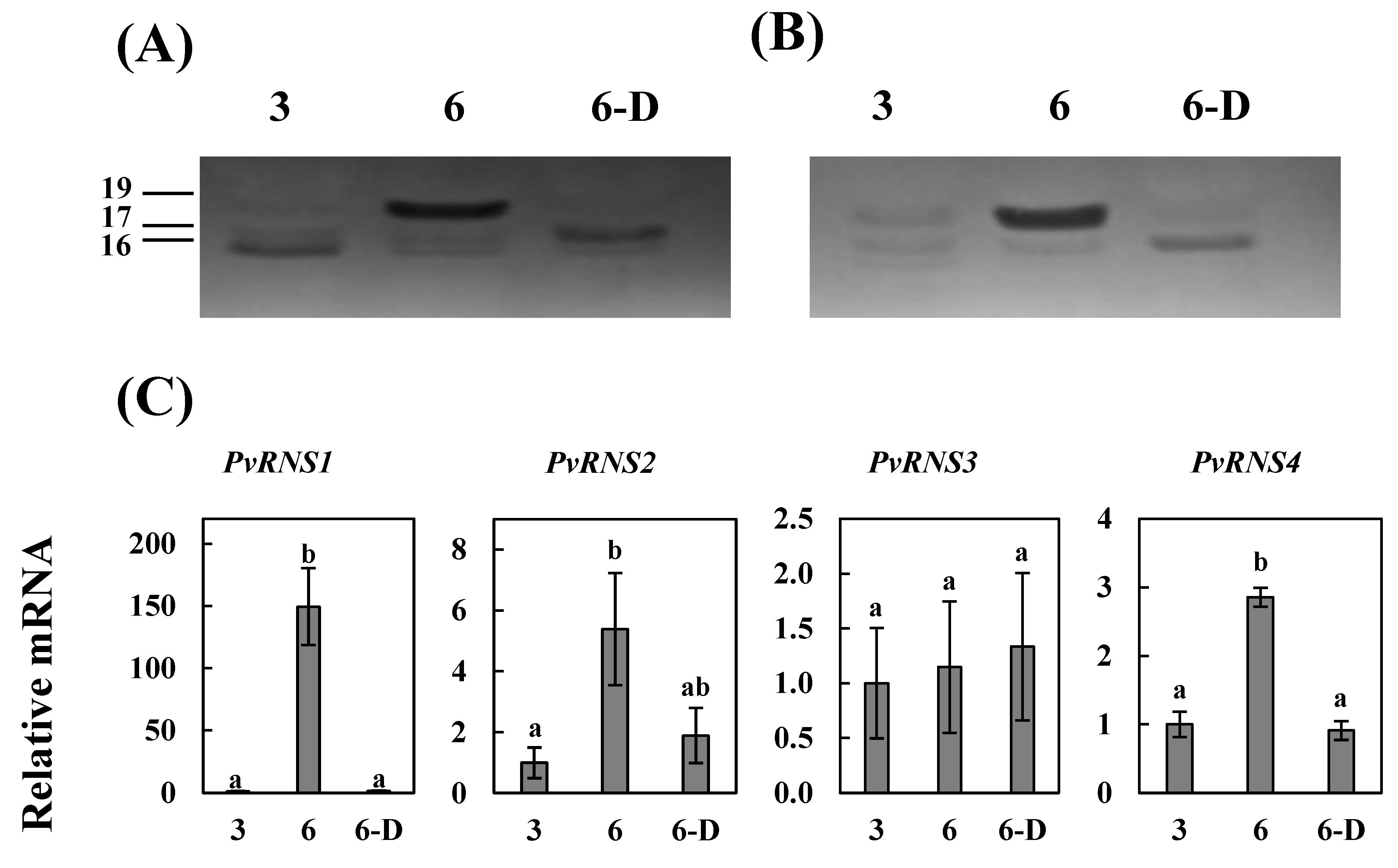
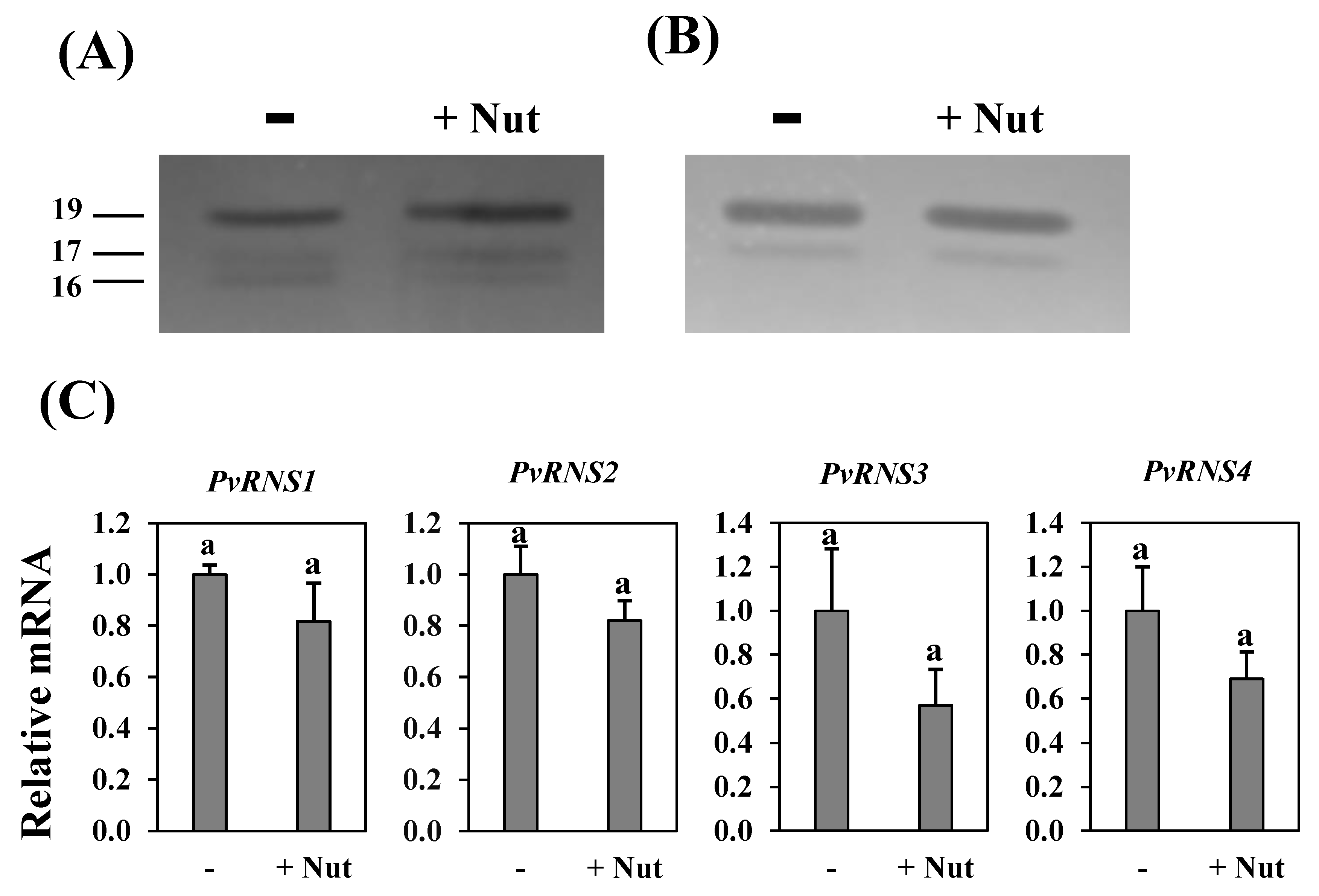
3.3. S-Like Ribonuclease Genes Expression in Cotyledons of Common Bean
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajjou, L.; Duval, M.; Galardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosental, L.; Nonogaki, H.; Fait, A. Activation and regulation of primary metabolism during seed germination. Seed Sci. Res. 2014, 24, 1–15. [Google Scholar] [CrossRef]
- Harada, T.; Miyairi, K.; Murai, N. Trimeric glycoproteins of bean seed storage protein phaseolin were purified from baculovirus-infected insect Sf9 cells for use of structural study. Plant Sci. 2010, 179, 123–132. [Google Scholar] [CrossRef]
- Lambert, R.; Cabello-Diaz, J.M.; Quiles, F.A.; Piedras, P. Identification of nucleases related to nutrient mobilization in senescing cotyledons from French bean. Acta Physiol. Plant. 2016, 38, 11. [Google Scholar] [CrossRef]
- Quiles, F.A.; Raso, M.J.; Pineda, M.; Piedras, P. Ureide metabolism during seedling development in French bean (Phaseolus vulgaris). Physiol. Plantarum 2009, 135, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.; Shane, M.W.; White, P.J. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, S.; Fu, Y.B.; Wang, H. Arabidopsis seed stored mRNAs are degraded constantly over aging time, as revealed by new quantification methods. Front. Plant Sci. 2020, 10, 1764. [Google Scholar] [CrossRef]
- Fleming, M.B.; Hill, L.M.; Walters, C. The kinetics of ageing in dry-stored seeds: A comparison of viability loss and RNA degradation in unique legacy seed collections. Ann. Bot. 2019, 123, 1133–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, N.; Rajjou, L.; North, H.M. Lost in translation: Physiological roles of stored mRNAs in seed germination. Plants 2020, 9, 347. [Google Scholar] [CrossRef] [Green Version]
- Rajjou, L.; Gallardo, K.; Debeaujon, I.; Vandekerckhove, J.; Job, C.; Job, D. The effect of a-Amanitin on the Arabidopsis seed proteome highlights the distinct roles of stored and neosynthesized mRNAs during germination. Plant Physiol. 2004, 134, 1598–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, N.; Permana, H.; Kumada, R.; Shinozaki, Y.; Tanabata, T.; Yamada, T.; Hirasawa, T.; Kanekatsu, M. Proteomic analysis of embryonic proteins synthesized from long-lived mRNAs during germination of rice seeds. Plant Cell Physiol. 2012, 53, 687–698. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Wang, Y.; He, Y.; Zhou, J.; Li, Y.; Liu, Q.; Xie, X. Overexpression of an S-like ribonuclease gene, OsRNS4, confers enhanced tolerance to high salinity and hyposensitivity to phytochrome-mediated light signals in rice. Plant Sci. 2014, 214, 99–105. [Google Scholar] [CrossRef]
- Luhtala, N.; Parker, R. T2 Family ribonucleases: Ancient enzymes with diverse roles. Trends Biochem. Sci. 2010, 35, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Xu, Y.; Zhao, H.; Li, Y. RNase T2 in inflammation and cancer: Immunological and biological views. Front. Immunol. 2020, 11, 1554. [Google Scholar] [CrossRef] [PubMed]
- MacIntosh, G.C.; Castandet, B. Organellar and secretory ribonucleases: Major players in plant RNA homeostasis. Plant Physiol. 2020, 183, 1438–1452. [Google Scholar] [CrossRef] [PubMed]
- Igic, B.; Kohn, J.R. Evolutionary relationships among self-incompatibility RNases. Proc. Natl. Acad. Sci. USA 2001, 98, 13167–13171. [Google Scholar] [CrossRef] [Green Version]
- MacIntosh, G.C.; Hillwig, M.S.; Meyer, A.; Flagel, L. RNase T2 genes from rice and the evolution of secretory ribonucleases in plants. Mol. Genet. Genom. 2010, 283, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Baena, M.; Galvez-Valdivieso, G.; Delgado-Garcia, E.; Pineda, M.; Piedras, P. Nuclease and ribonuclease activities in response to salt stress: Identification of PvRNS3, a T2/S-like ribonuclease induced in common bean radicles by salt stress. Plant Physiol. Biochem. 2020, 147, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J. The ribonucleases of higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1994, 45, 421–445. [Google Scholar] [CrossRef]
- Ramanauskas, K.; Igić, B. The evolutionary history of plant T2/S-type ribonucleases. PeerJ 2017, 5, e3790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, R.; Quiles, F.A.; Cabello-Diaz, J.M.; Piedras, P. Purification and identification of a nuclease activity in embryo axes from French bean. Plant Sci. 2014, 224, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Lambert, R.; Quiles, F.A.; Galvez-Valdivieso, G.; Piedras, P. Nucleases activities during French bean leaf aging and dark-induced senescence. J. Plant Physiol. 2017, 218, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant Soil 2003, 252, 55–122. [Google Scholar] [CrossRef] [Green Version]
- Galvez-Valdivieso, G.; Alamillo, J.M.; Fernandez, J.; Pineda, M. Molecular characterization of PVAS3: An asparagine synthetase gene from common bean prevailing in developing organs. J. Plant Physiol. 2013, 170, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Melino, V.J.; Casartelli, A.; George, J.; Rupasinghe, T.; Roessner, U.; Okamoto, M.; Heuer, S. RNA catabolites contribute to the nitrogen pool and support growth recovery of wheat. Front. Plant Sci. 2018, 9, 1539. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.H.; Kawamata, T.; Horie, T.; Tsugawa, H.; Nakayama, Y.; Ohsumi, Y.; Fukusaki, E. Bulk RNA degradation by nitrogen starvation-induced autophagy in yeast. EMBO J. 2015, 34, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Ananieva, K.; Ananiev, E.D.; Mishev, K.; Georgieva, K.; Tzvetkova, N.; Van Staden, J. Changes in photosynthetic capacity and polypeptide patterns during natural senescence and rejuvenation of Cucurbita pepo L (zucchini) cotyledons. Plant Growth Regul. 2008, 54, 23–29. [Google Scholar] [CrossRef]
- Wilhelmova, N.; Prochazkova, D.; Machackova, I.; Vagner, M.; Srbova, M.; Wilhelm, J. The role of cytokinins and ethylene in bean cotyledon senescence. The effect of free radicals. Biol. Plantarum 2004, 48, 523–529. [Google Scholar] [CrossRef]
- Jasid, S.; Galatro, A.; Villordo, J.J.; Puntarulo, S.; Simontacchi, M. Role of nitric oxide in soybean cotyledon senescence. Plant Sci. 2009, 176, 662–668. [Google Scholar] [CrossRef]
- Verma, G.; Mishra, S.; Sangwan, N.; Sharma, S. Reactive oxygen species mediate axis-cotyledon signaling to induce reserve mobilization during germination and seedling establishment in Vigna Radiat. J. Plant Physiol. 2015, 184, 79–88. [Google Scholar] [CrossRef]
- Quiles, F.A.; Galvez-Valdivieso, G.; Guerrero-Casado, J.; Pineda, M.; Piedras, P. Relationship between ureidic/amidic metabolism and antioxidant enzymatic activities in legume seedlings. Plant Physiol. Biochem. 2019, 138, 1–8. [Google Scholar] [CrossRef]
- Wojtyla, L.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different Modes of Hydrogen Peroxide Action during Seed Germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gho, Y.S.; Choi, H.; Moon, S.; Song, M.Y.; Park, H.; Kim, D.H.; Ha, S.H.; Jung, K.H. Phosphate-starvation-inducible S-like RNase genes in rice are involved in phosphate source recycling by RNA decay. Front. Plant Sci. 2020, 11, 585561. [Google Scholar] [CrossRef] [PubMed]
- Hillwig, M.S.; Contento, A.L.; Meyer, A.; Ebany, D.; Bassham, D.C.; Macintosh, G.C. RNS2, a conserved member of the RNase T2 family, is necessary for ribosomal RNA decay in plants. Proc. Natl. Acad. Sci. USA 2011, 108, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Floyd, B.E.; Mugume, Y.; Morriss, S.C.; MacIntosh, G.C.; Bassham, D.C. Localization of RNS2 ribonuclease to the vacuole is required for its role in cellular homeostasis. Planta 2017, 245, 779–792. [Google Scholar] [CrossRef]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The master of bulk and selective recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Chen, Q.; Have, M. Regulation of nutrient recycling via autophagy. Curr. Opin. Plant Biol. 2017, 39, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.W.; Shinozaki, D.; Luo, J.; Pottier, M.; Have, M.; Marmagne, A.; Reisdorf-Cren, M.; Chardon, F.; Thomine, S.; Yoshimoto, K.; et al. Autophagy and nutrients management in plants. Cells 2019, 8, 1426. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lin, Y.; Li, X.; Guo, S.; Huang, Y.; Xie, Q. Autophagy dances with phytohormones upon multiple stresses. Plants 2020, 9, 1038. [Google Scholar] [CrossRef] [PubMed]
- Avin-Wittenberg, T. Autophagy and its role in plant abiotic stress management. Plant Cell Environ. 2019, 42, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Tarkowski, K.P.; den Ende, W.V.; Bassham, D.C. Linking autophagy to abiotic and biotic stress response. Trends Plant Sci. 2019, 24, 413–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bariola, P.A.; Howard, C.J.; Taylor, C.B.; Verburg, M.T.; Jaglan, V.D.; Green, P.J. The Arabidopsis ribonuclease gene RNS1 is tightly controlled in response to phosphate limitation. Plant J. 1994, 6, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Have, M.; Marmagne, A.; Chardon, F.; Masclaux-Daubresse, C. Nitrogen remobilization during leaf senescence: Lessons from Arabidopsis to crops. J. Exp. Bot. 2017, 68, 2513–2529. [Google Scholar] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Baena, M.; Delgado-García, E.; Pineda, M.; Galvez-Valdivieso, G.; Piedras, P. S-Like Ribonuclease T2 Genes Are Induced during Mobilisation of Nutrients in Cotyledons from Common Bean. Agronomy 2021, 11, 490. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030490
Diaz-Baena M, Delgado-García E, Pineda M, Galvez-Valdivieso G, Piedras P. S-Like Ribonuclease T2 Genes Are Induced during Mobilisation of Nutrients in Cotyledons from Common Bean. Agronomy. 2021; 11(3):490. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030490
Chicago/Turabian StyleDiaz-Baena, Mercedes, Elena Delgado-García, Manuel Pineda, Gregorio Galvez-Valdivieso, and Pedro Piedras. 2021. "S-Like Ribonuclease T2 Genes Are Induced during Mobilisation of Nutrients in Cotyledons from Common Bean" Agronomy 11, no. 3: 490. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030490