
Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
Experimental Conditions
3. Experimental Factors
3.1. Amylolytic Enzymes Determination
3.2. IAA and GA3 Determination
3.3. Trans-Zeatin and Trans-Zeatin-Riboside Determination
3.4. Measurement of Pea Seedlings Organs
3.5. Statistics
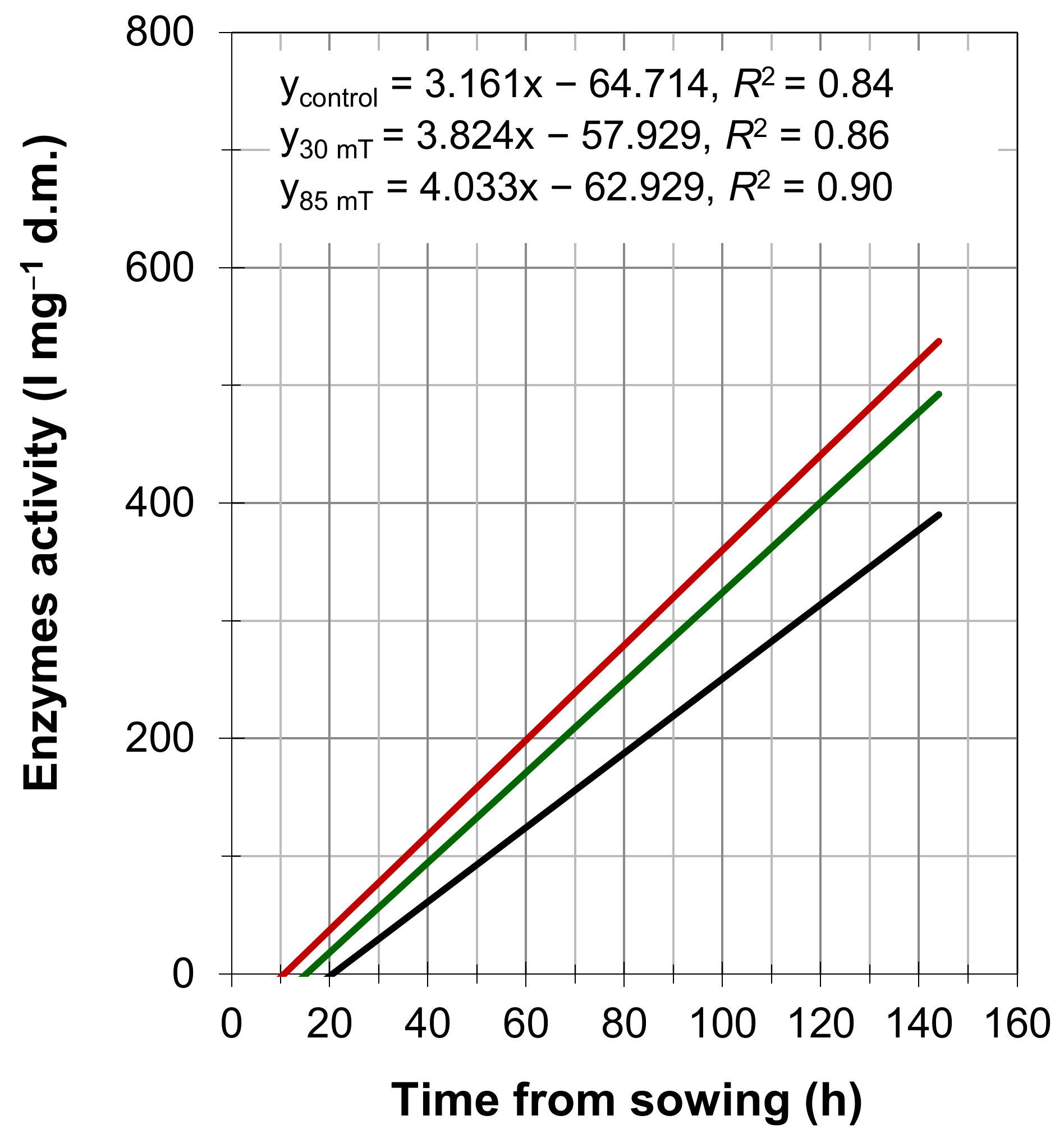
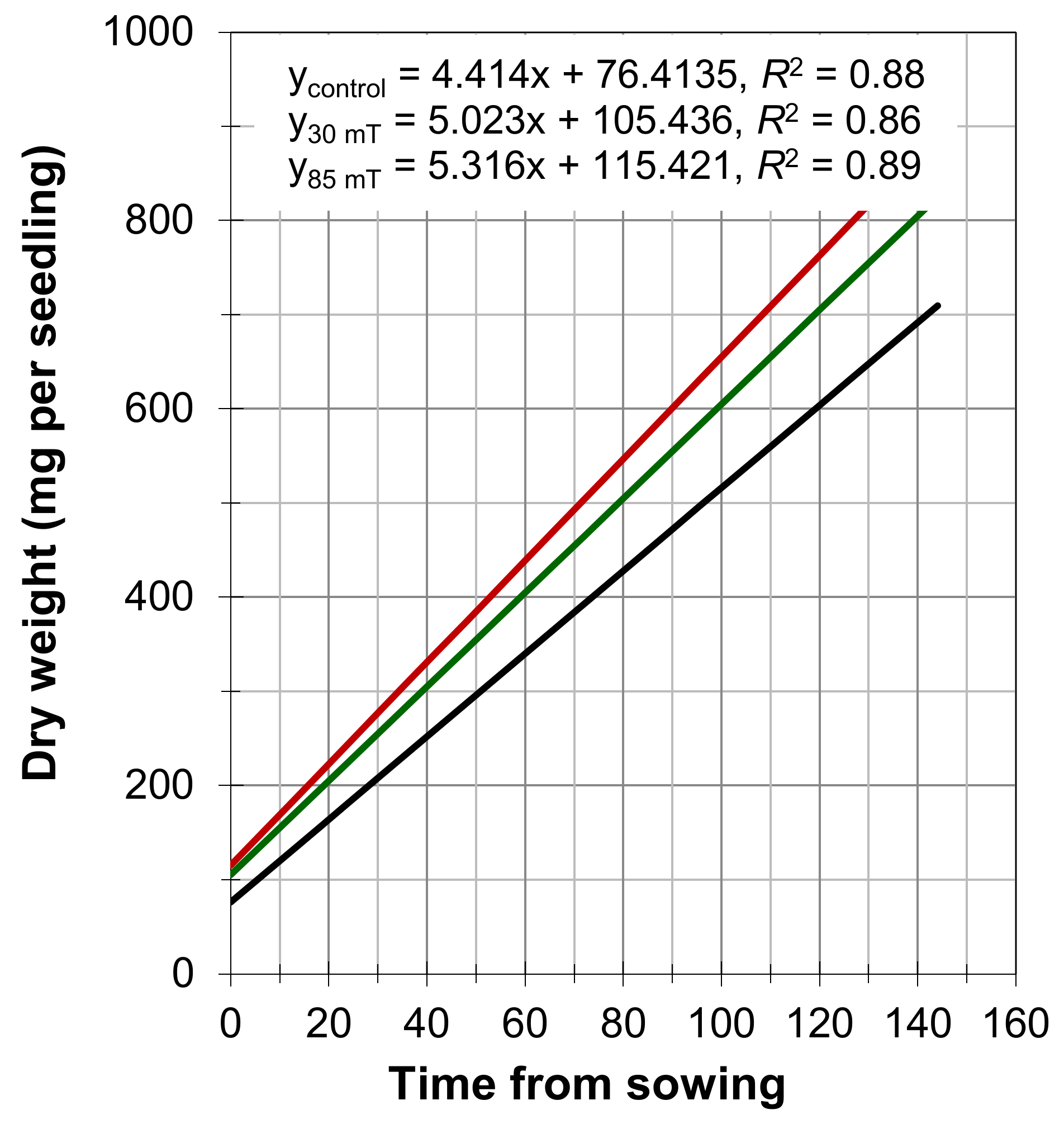
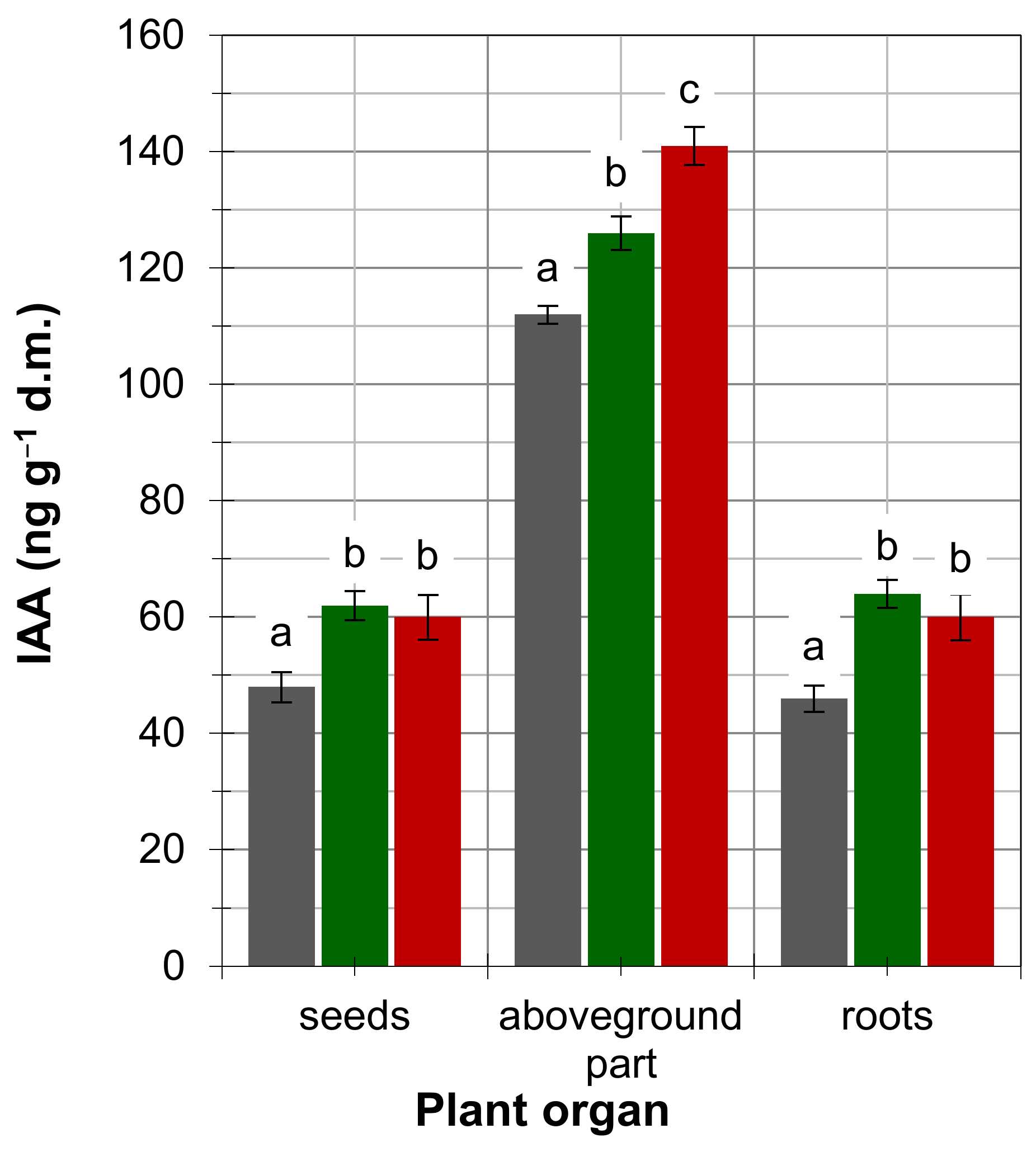
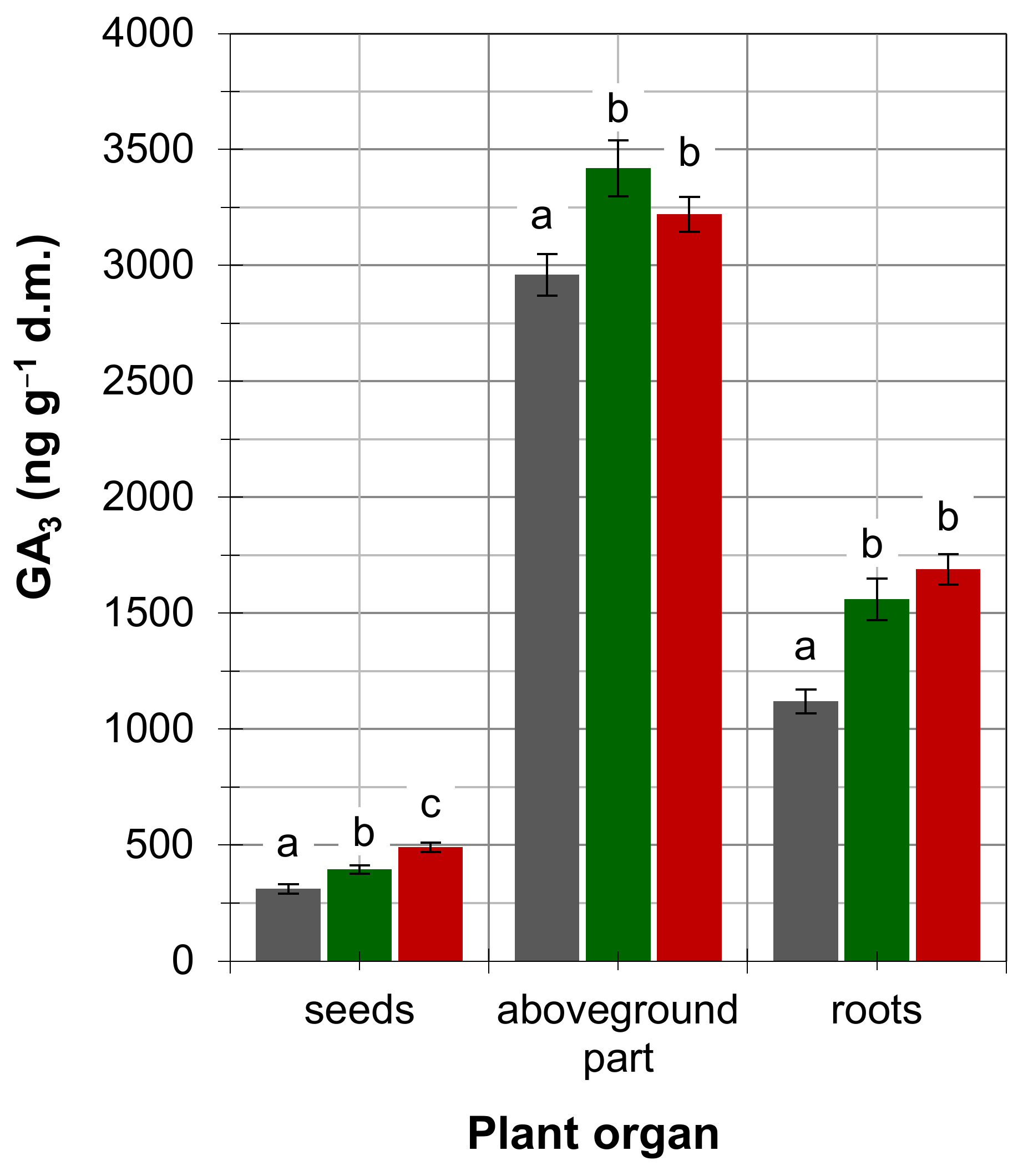
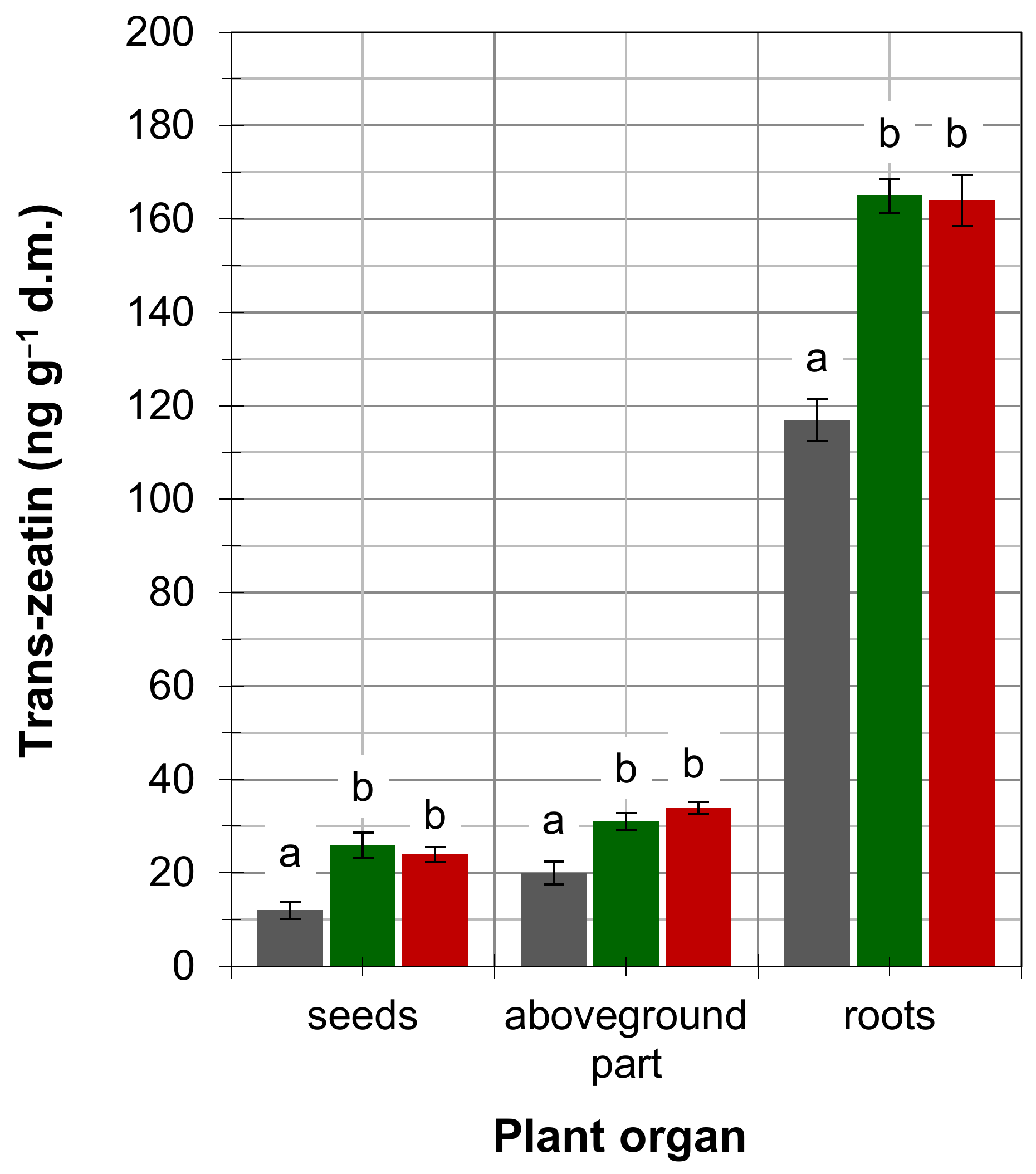
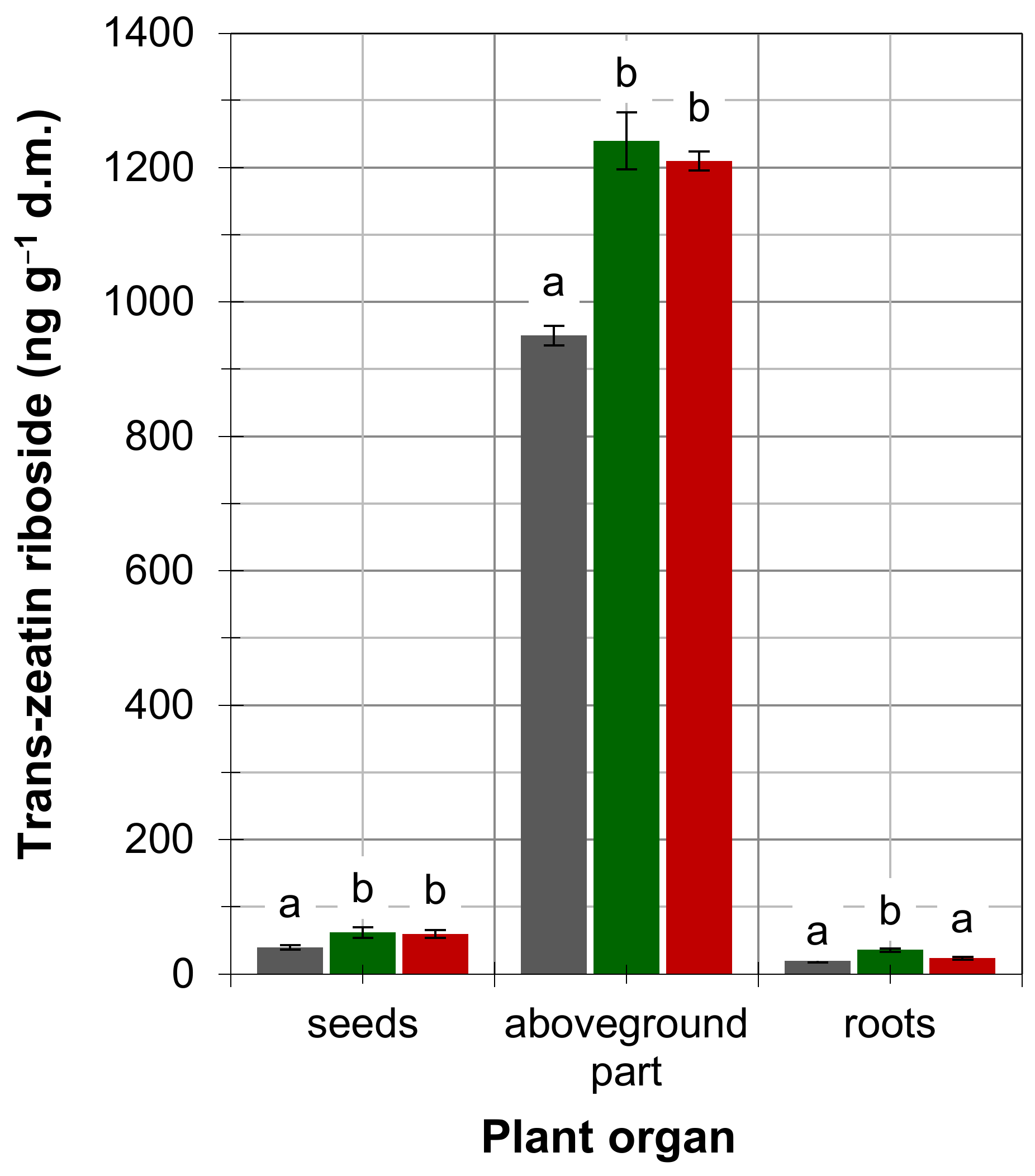
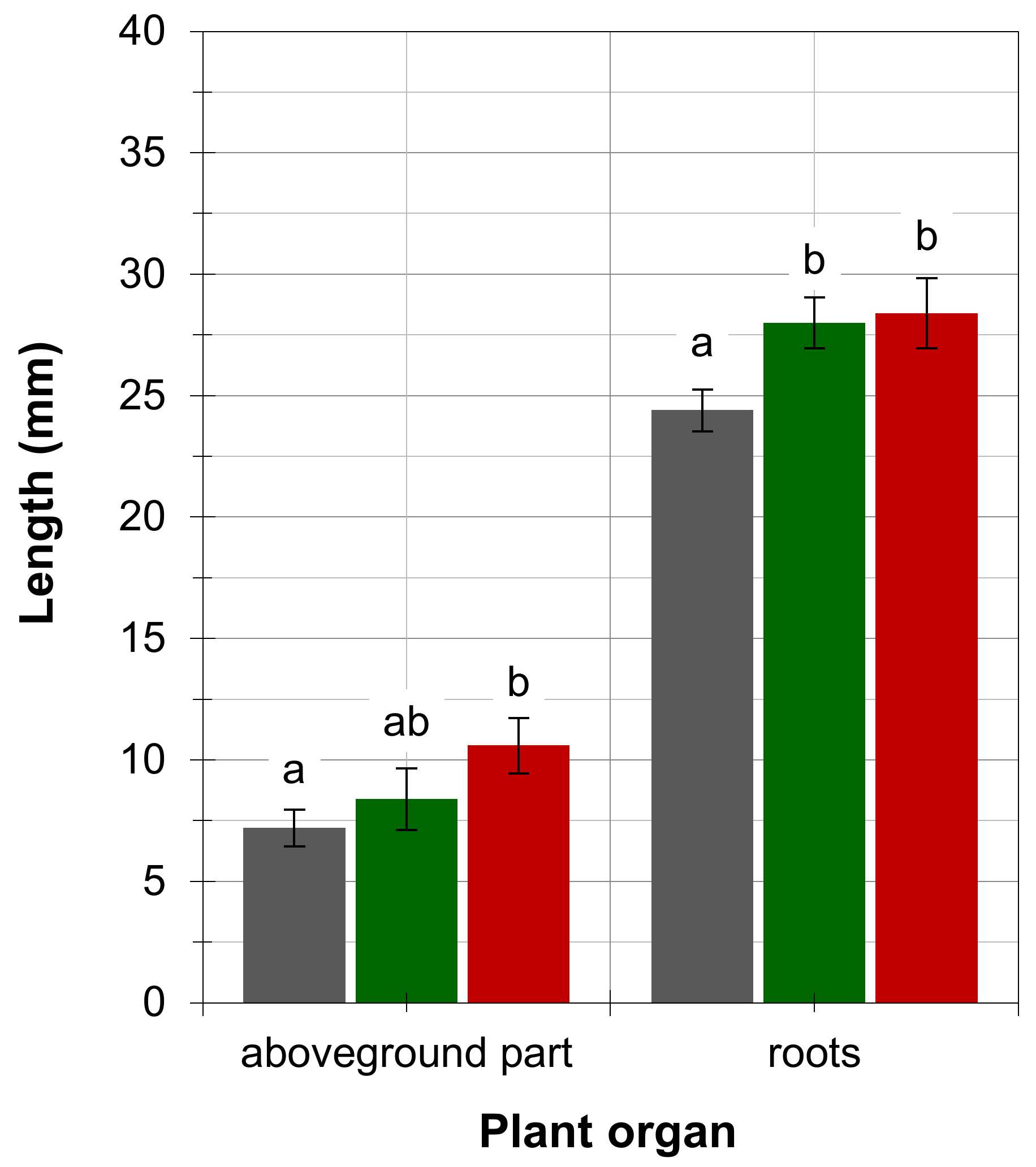
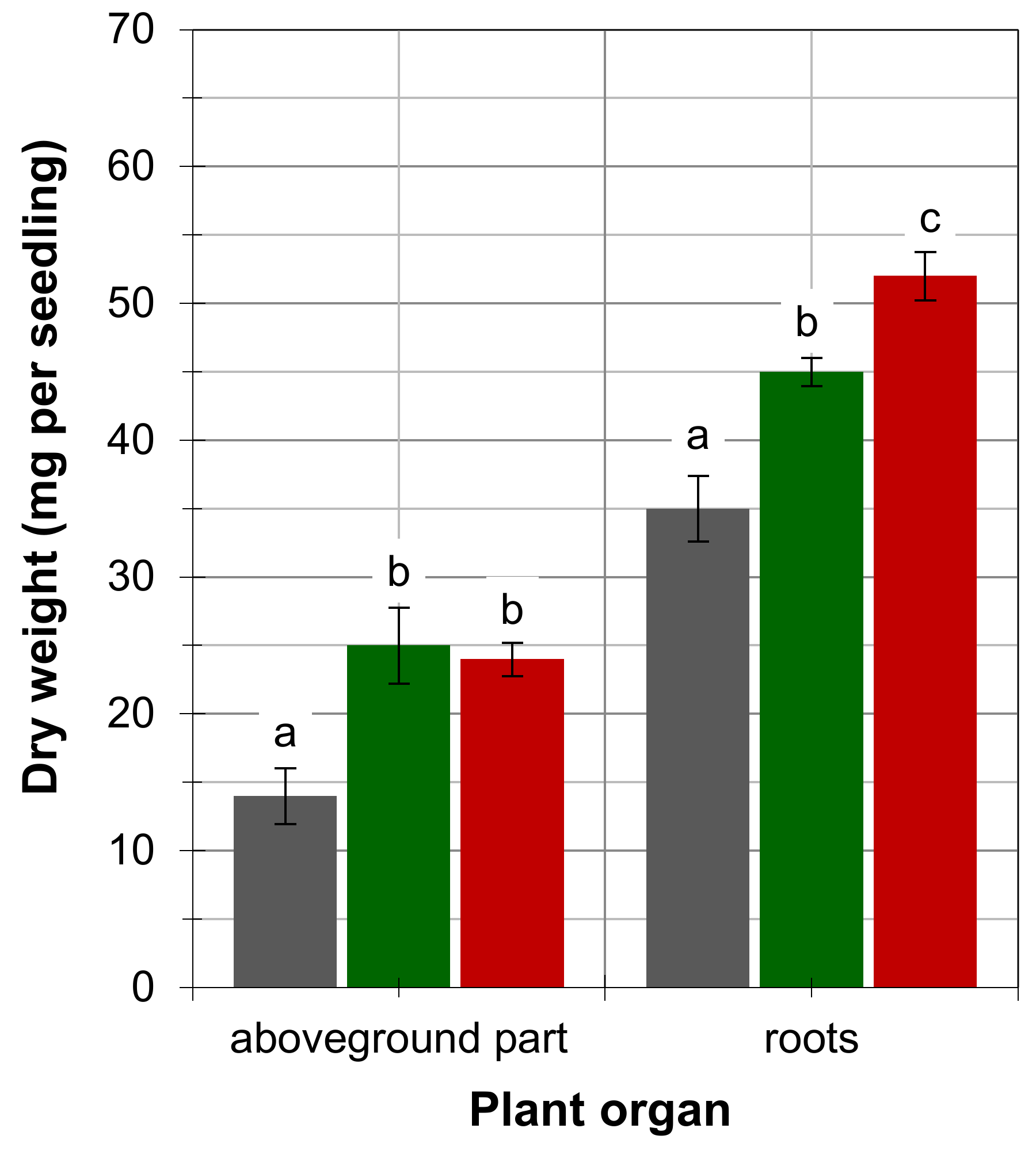
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oleksiak, T. Winter wheat yields depending on the quality of applied sowing material. Biuletyn Inst. Hodowli Aklim. Roślin 2009, 251, 83–93. (In Polish) [Google Scholar]
- Grzesik, M.; Janas, R.; Górniak, K.; Romanowska-Duda, Z. Biological and physical methods of seed production and processing. J. Agric. Eng. Res. 2012, 57, 147–152. [Google Scholar]
- Jakubowski, T. Evaluation of the impact of pre-sowing microwave stimulation of bean seeds on the germination process. Agric. Eng. 2015, 2, 45–56. [Google Scholar]
- Jalali, A.H.; Salehi, F. Sugar beet Yield as affected by seed priming and weed control. Arch. Agron. Soil Sci. 2013, 59, 281–288. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Dominguez, P.A.; Cruz, O.A.; Ivanov, R.; Caballo, C.Z.; Zepeda, B.R. Laser in agriculture. Int. Agrophys. 2010, 24, 407–422. [Google Scholar]
- Szajsner, H. The analysis of laser radiation treatment effects on grains of selected genotypes of cereals—Analiza efektów działania promieniowania laserowego na ziarniaki wybranych genotypów roślin zbożowych. Zesz. Nauk. Uniw. Przyr. We Wrocławiu. Rozpr. 2009, 256, 1–98. (In Polish) [Google Scholar]
- Gładyszewska, B. Estimation of a laser biostimulation dose. Int. Agrophys. 2011, 25, 403–405. [Google Scholar]
- Szajsner, H.; Koszelnik-Leszek, A.; Nowosad, K. Influence of laser stimulation seeds on germination and initial growth of seeding Silene vulgaris. Pol. Nat. Sci. 2014, 29, 5–16. [Google Scholar]
- Prokop, M.; Pietruszewski, S.; Kornarzyński, K. The preliminary investigation of magnetic and electric fields influence on germination, crops, and mechanical features of radish and skall radish roots. Acta Agrophys. 2002, 62, 83–95. [Google Scholar]
- Samuilov, F.D.; Garifullina, R.L. Effect of laser irradiation on microviscosity of aqueous medium in imbibing maize seeds as studied with a spain probe method. Russ. J. Plant Physiol. 2007, 54, 128–131. [Google Scholar] [CrossRef]
- Khristyuk, V.T. Effect of a Low-Frequency Electromagnetic Field on Growth and Malt quality of brewer’s barley. Russ. Agric. Sci. 2009, 35, 429–431. [Google Scholar] [CrossRef]
- Shine, M.B.; Guruprasad, K.N.; Anand, A. Enhancement of germination, growth, and photosynthesis in soybean by pretreatment of seeds with magnetic field. Bioelectromagnetics 2011, 32, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Kumari, A.; Thakur, M.; Koul, A. Hydrogen peroxide signaling integrates with phytohormones during the germination of magnetoprimed tomato seeds. Sci. Rep. 2019, 9, 8814. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Anand, A.; Nagarajan, S. Biochemical and biophysical changes associated with magnetopriming in germinating cucumber seeds. Plant Physiol. Biochem. 2012, 57, 67–73. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Guruprasad, K. Acceleration of germination and early growth characteristics of soybean and maize after pre-treatment of seeds with static magnetic field. Int. J. Trop. Agric. 2015, 33, 985–992. [Google Scholar]
- Sarraf, M.; Kataria, S.; Taimourya, H.; Santos, L.O.; Menegatti, R.D.; Jain, M.; Ihtisham, M.; Liu, S. Magnetic Field (MF) Applications in Plants: An Overview. Review. Plants 2020, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, T.; Dumlupinar, R.; Erdal, S. Acceleration of germination and early growth of wheat and bean seedlings grown under various magnetic field and osmotic conditions. Bioelectromagnetics 2010, 31, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Podleśny, J.; Pietruszewski, S.; Podleśna, A. Influence of magnetic stimulation of seeds on the formation of morphological features and yielding of the pea. Int. Agrophys. 2005, 19, 1–8. [Google Scholar]
- Vashisth, A.; Nagarajan, S. Exposure of seeds to static magnetic field enhances germination and early growth characteristics in chickpea (Cicer arietinum L.). Bioelectromagnetics 2008, 29, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Jankiewicz, L.S. Regulators of Plant Growth and Development—Properties and Action—Regulatory Wzrostu I Rozwoju Roślin—Właściwości I Działanie; PWN: Warszawa, Polish, 1997; pp. 1–282. (In Polish) [Google Scholar]
- Ross, J.J.; O’Neill, D.P.; Smith, J.J.; Kerckhoffs, L.H.; Elliott, R.C. Evidence that auxin promotes gibberellin A1 biosynthesis in pea. Plant J. 2000, 21, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Olszewski, N.; Sun, T.P.; Gubler, F. Gibberellin signaling:biosynthesis, catabolism, and response pathways. Plant Cell 2002, 14, S61–S80. [Google Scholar] [CrossRef] [Green Version]
- Podleśny, J.; Podleśna, A. The effect of different levels of soil moisture on development and yielding of two different genotypes of white lupine (Lupinus albus L.). Biul. Inst. Hod. I Aklim. Roślin 2003, 228, 315–322. [Google Scholar]
- Jarzyniak, K. The role of cytokinins in Legume-Rhizobium symbiosis. Postępy Biochem. 2020, 66, 229–238. (In Polish) [Google Scholar] [CrossRef] [PubMed]
- Pietruszewski, S.; Kornarzyński, K. Magnetic biostimulation of wheat seeds. Int. Agroph. 1999, 13, 497–501. [Google Scholar]
- Somogy-Nelson, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Hardin, J.M.; Stutte, C.A. Analysis of plant hormones using high-performance liquid chromatography. J. Chromatogr. 1981, 208, 124–128. [Google Scholar] [CrossRef]
- Hirosava, E. Auxins induce α-amylase activity in pea cotyledons. Plant Physiol. 1989, 916, 484–486. [Google Scholar]
- Podleśna, A.; Bojarszczuk, J.; Podleśny, J. Effect of pre-sowing magnetic field treatment on some biochemical and physiological processes in faba bean (Vicia faba L. spp. Minor). J. Plant. Growth Regul. 2019, 38, 1153–1160. [Google Scholar]
- Podleśny, J. Studies on influence of laser light on seeds, growth, development and yielding of the white lupine (Lupinus album L.) plants. Monogr. I Rozpr. Nauk. 2002, 3, 1–59. (In Polish) [Google Scholar]
- Wood, A.J.; Roper, J. A simple and nondestructive technique for measuring plant growth and development. Am. Biol. Teach. 2000, 62, 215–217. [Google Scholar] [CrossRef]
- Pittman, U.J. Magnetism and plant growth. I. Effect on germination and early growth of cereal seeds. Can. J. Plant. Sci. 1963, 43, 513–518. [Google Scholar] [CrossRef]
- Galland, P.; Pazur, A. Magnetoreception in plants. J. Plant Res. 2005, 118, 371–389. [Google Scholar] [CrossRef]
- Alikamanoglu, S.; Sen, A. Stimulation of growth and some biochemical parameters by magnetic field in wheat (Triticum aestivum L.) tissue cultures. Afric. J. Biotech. 2011, 10, 10957–10963. [Google Scholar]
- Selim, A.H.; El-Nady, M.F. Physio-anatomical responses of drought stressed tomato plants to magnetic field. Acta Astronaut. 2011, 69, 387–396. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kumari, B.D.R. Pulsed magnetic field: A contemporary approach offers to enhance plant growth and yield of soybean. Plant Physiol. Biochem. 2012, 51, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Selim, D.A.F.H.; Nassar, R.M.A.; Boghdady, M.S.; Bonfill, M. Physiological and anatomical studies of two wheat cultivars irrigated with magnetic water under drought stress conditions. Plant Physiol. Biochem. 2019, 135, 480–488. [Google Scholar] [CrossRef]
- Iqbal, M.; Haq, Z.; Jamil, Y.; Ahmad, M.R. Effect of pre-sowing magnetic treatment on growth and chlorophyll contents of pea. Int. Agrophys. 2012, 26, 211–214. [Google Scholar] [CrossRef]
- Carbonell, M.V.; Florez, M.; Martinez, E.; Maqueda, R.; Amaya, J.M. Study of stationary magnetic fields on initial growth of pea (Pisum sativum L.) seeds. Seed Sci. Technol. 2011, 39, 673–679. [Google Scholar] [CrossRef]
- Podleśny, J.; Sowiński, M. The effect of seeds stimulation by magnetic field on growth, development and dynamics of biomass accumulation in pea (Pisum sativum L.)—Wpływ traktowania nasion polem magnetycznym na wzrost, rozwój i dynamikę gromadzenia masy grochu siewnego (Pisum sativum L.). Inż. Rol. 2005, 4, 103–110. (In Polish) [Google Scholar]
- Podleśny, J.; Pietruszewski, S. The effect of magnetic stimulation of seeds on growth and cropping of seed pea grown at varying soil moisture content—Wpływ stymulacji magnetycznej nasion na wzrost i plonowanie grochu siewnego uprawianego przy różnej wilgotności gleby. Inż. Rol. 2007, 8, 207–212. (In Polish) [Google Scholar]
- Selim, A.; Zayed, M.A.; Zayed, M. Magnetic field treated water effects on germination, growth and physio-chemical aspects of some economic plants. Acta Bot. Hung. 2013, 55, 99–116. [Google Scholar] [CrossRef]
- Piacentini, M.P.; Fraternale, D.; Piatti, E.; Ricci, D.; Vetrano Dachà, M.; Accorsi, A. Senescence delay and change of antioxidant enzyme levels in Cumcumis sativus L. etiolated seedling by ELF magnetic fields. Plant Sci. 2001, 161, 45–53. [Google Scholar] [CrossRef]
- Dhavi, F. Why Magnetic Fields are Used to Enhance a Plant’s Growth and Productivity? Ann. Res. Rev. Biol. 2014, 4, 886–896. [Google Scholar]
- Atak, Ҁ.; Emiriğlu, O.; Alikamanoğlu, S.; Rzakoulieva, A. Stimulation of regeneration by magnetic field in soybean (Glycine max. L. Merrill) tissue cultures. J. Mol. Biol. 2003, 2, 113–119. [Google Scholar]
- De Souza, A.; García, D.; Sueiro, L.; Licea, L.; Porras, E. Pre-sowing magnetic treatment of tomato seeds: Effects on the growth and yield of plants cultivated late in the season. Span. J. Agric. Res. 2005, 3, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.J. Plant Hormones: Biosynthesis, Signal Transduction, Action! Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podleśny, J.; Podleśna, A.; Gładyszewska, B.; Bojarszczuk, J. Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings. Agronomy 2021, 11, 494. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030494
Podleśny J, Podleśna A, Gładyszewska B, Bojarszczuk J. Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings. Agronomy. 2021; 11(3):494. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030494
Chicago/Turabian StylePodleśny, Janusz, Anna Podleśna, Bożena Gładyszewska, and Jolanta Bojarszczuk. 2021. "Effect of Pre-Sowing Magnetic Field Treatment on Enzymes and Phytohormones in Pea (Pisum sativum L.) Seeds and Seedlings" Agronomy 11, no. 3: 494. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11030494