
Nutrient Dynamics in Sandy Soil and Leaf Lettuce Following the Application of Urea and Urea-Hydrogen Peroxide Impregnated Co-Pyrolyzed Animal Manure and Bone Meal


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nutrient Dynamics Parameters Assessed
2.2. Preparation of the Enriched Biochars and Experimental Set-Up
2.3. Labotory Methods Used for the Assessment of NEBs, Soil and Plant Tissues
2.4. Statistical Analysis
3. Results
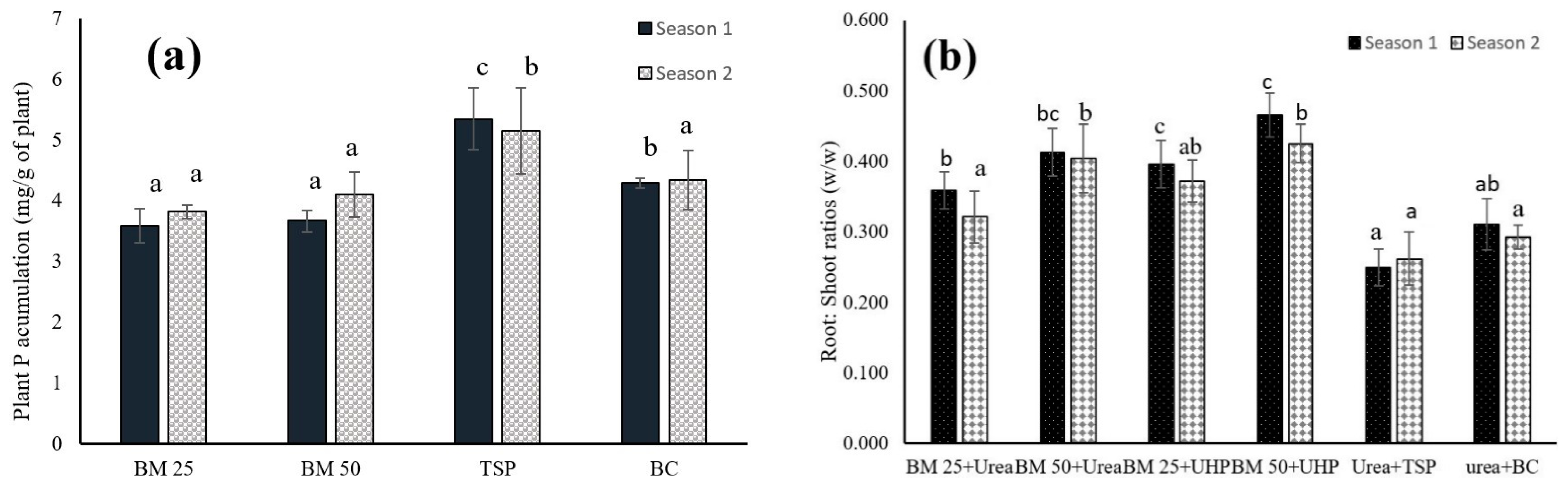
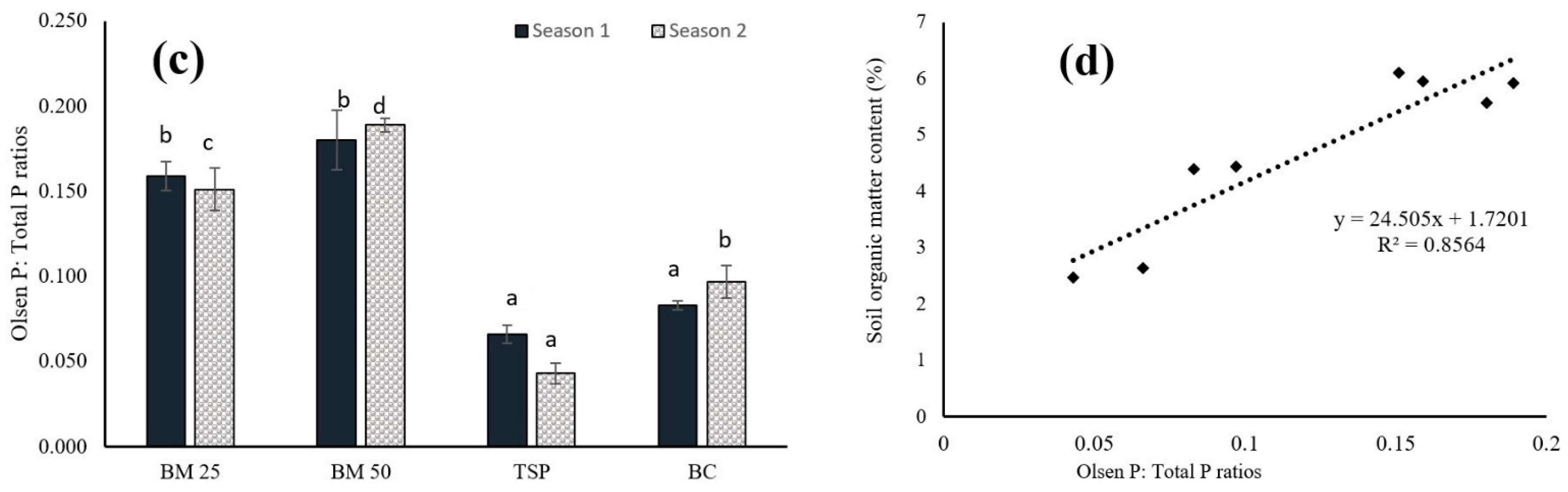
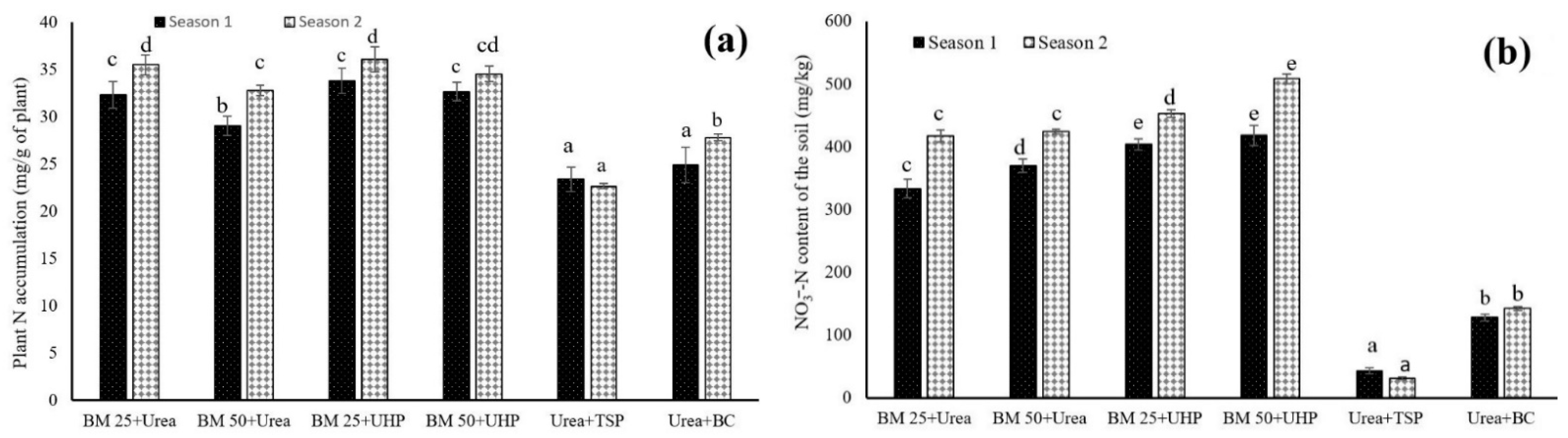
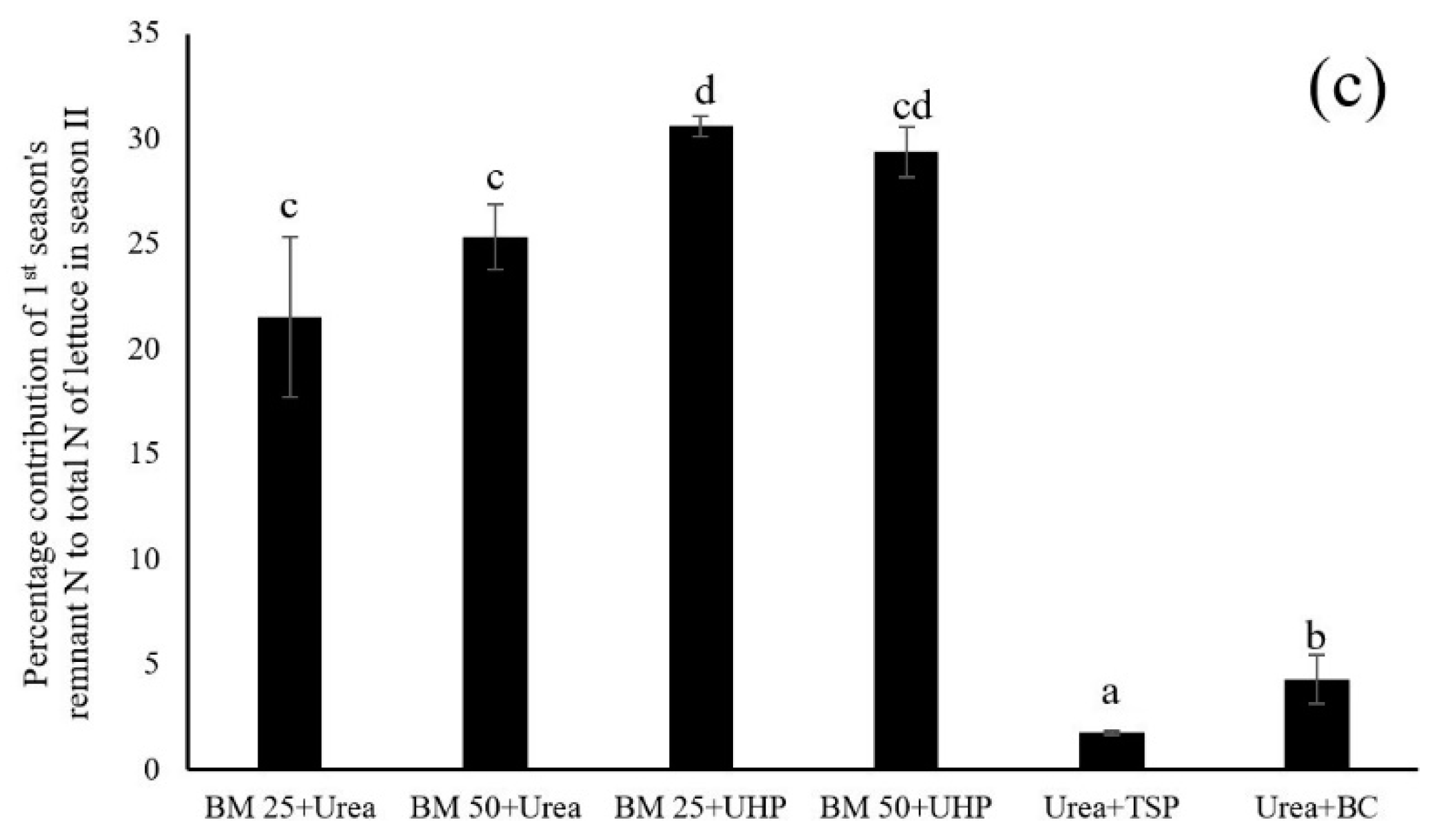
3.1. Dynamics of N and P in Both the Leaf Lettuce and Soil
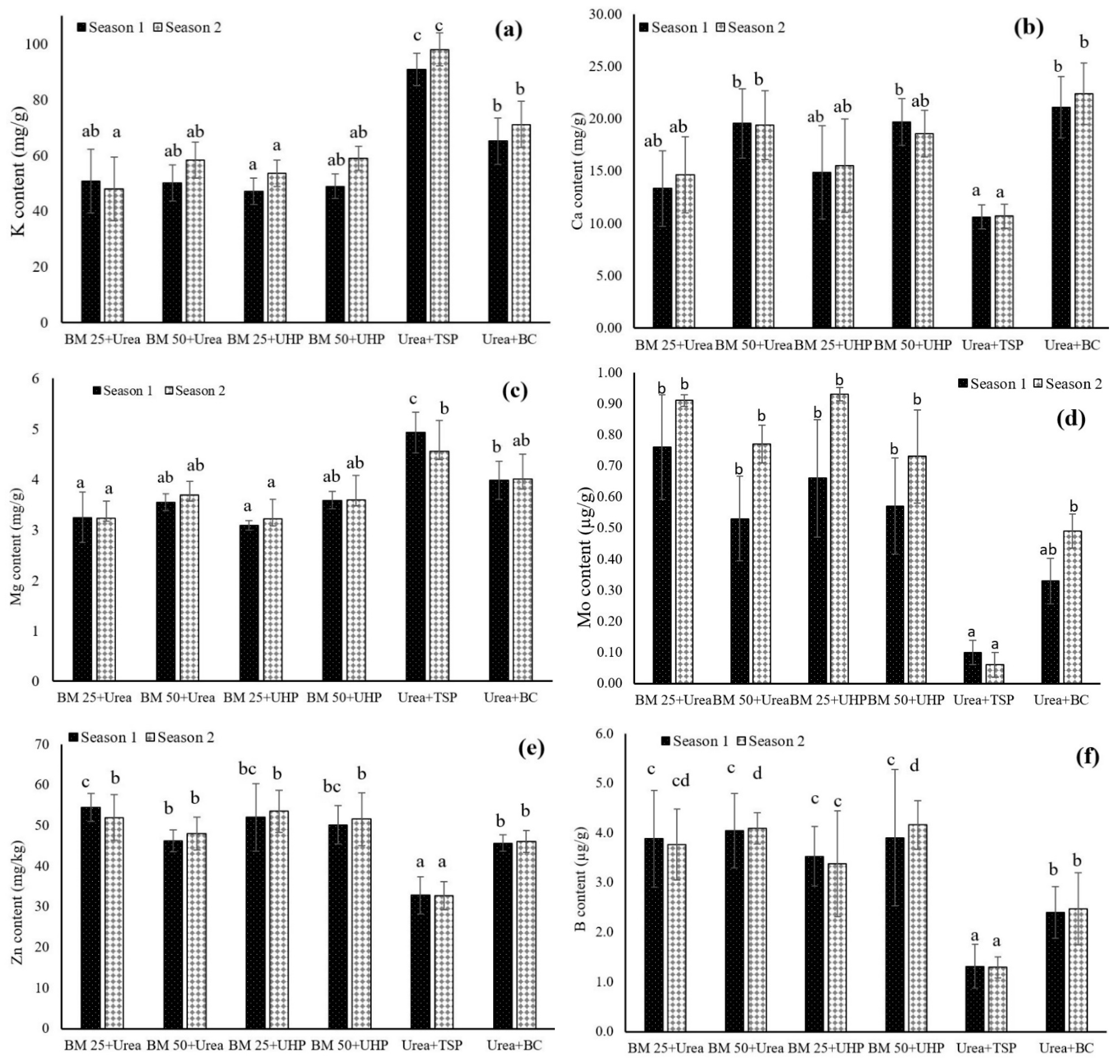
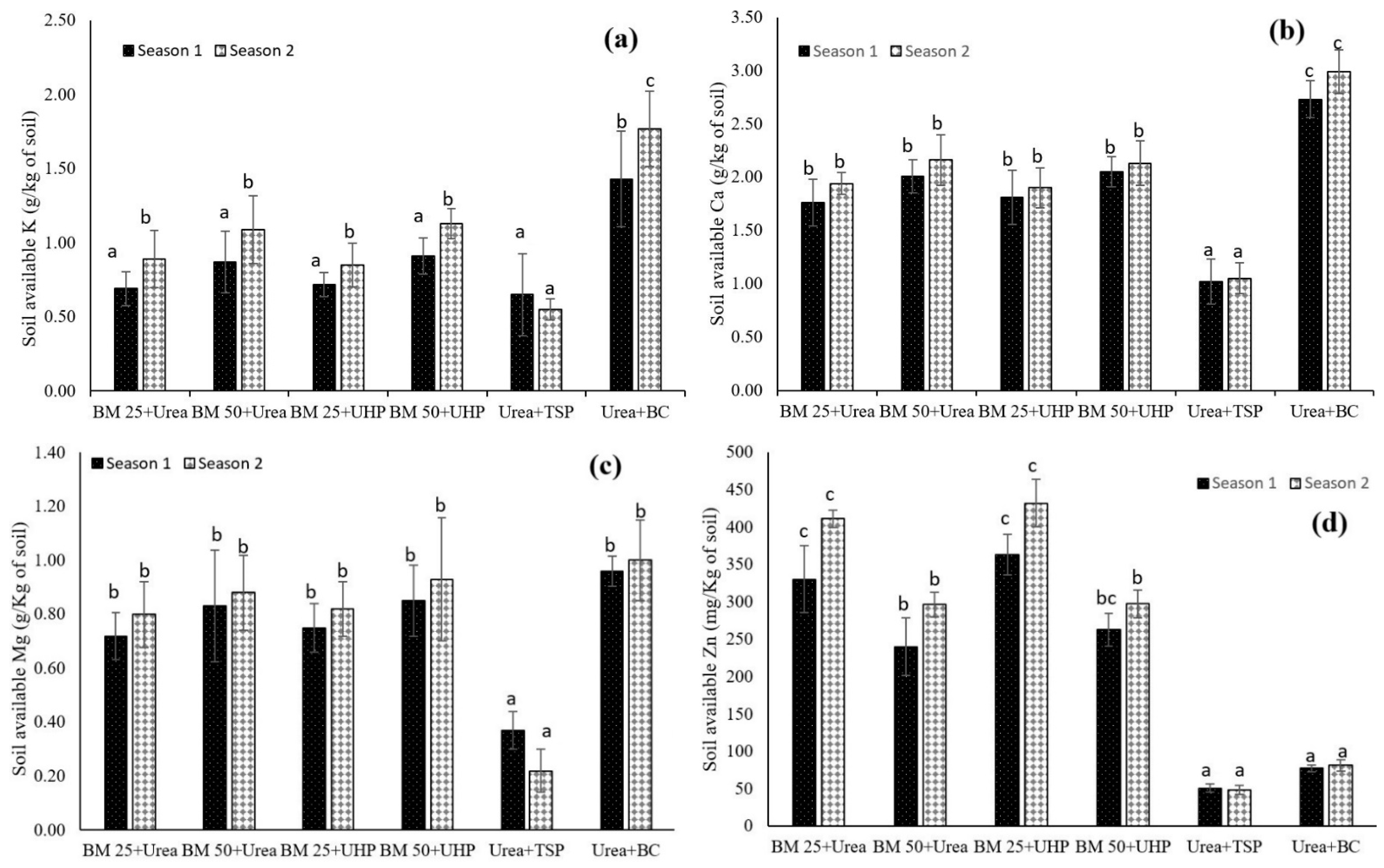
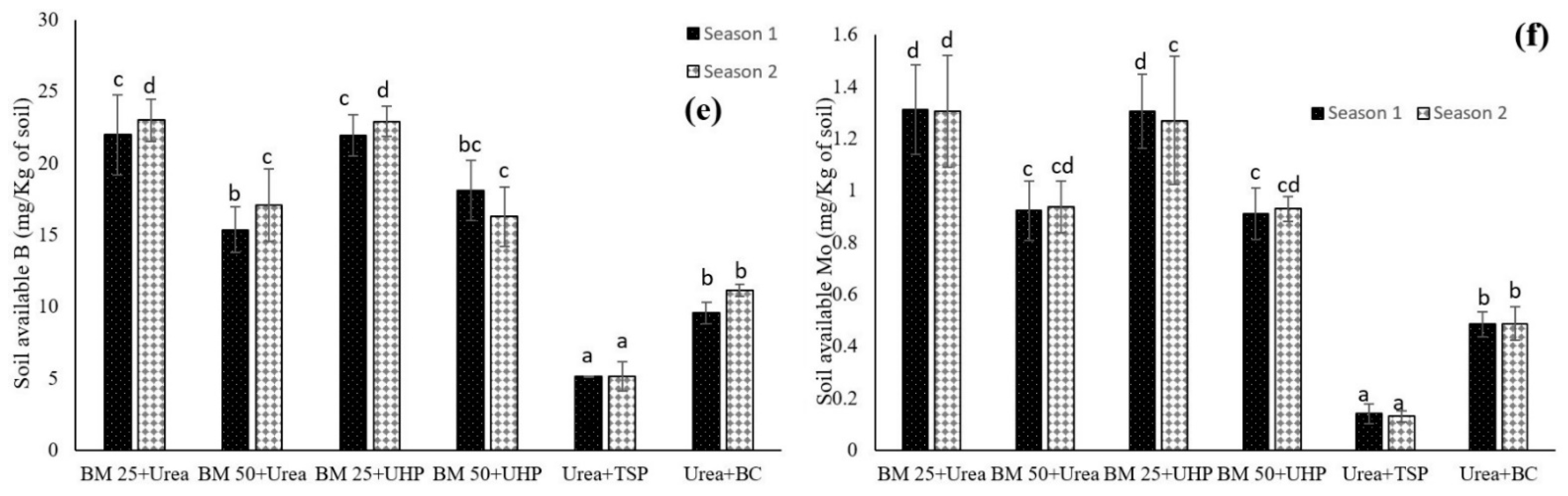
3.2. Dynamics of Other Macronutrients and Micronutrients in Both the Leaf Lettuce and Soil
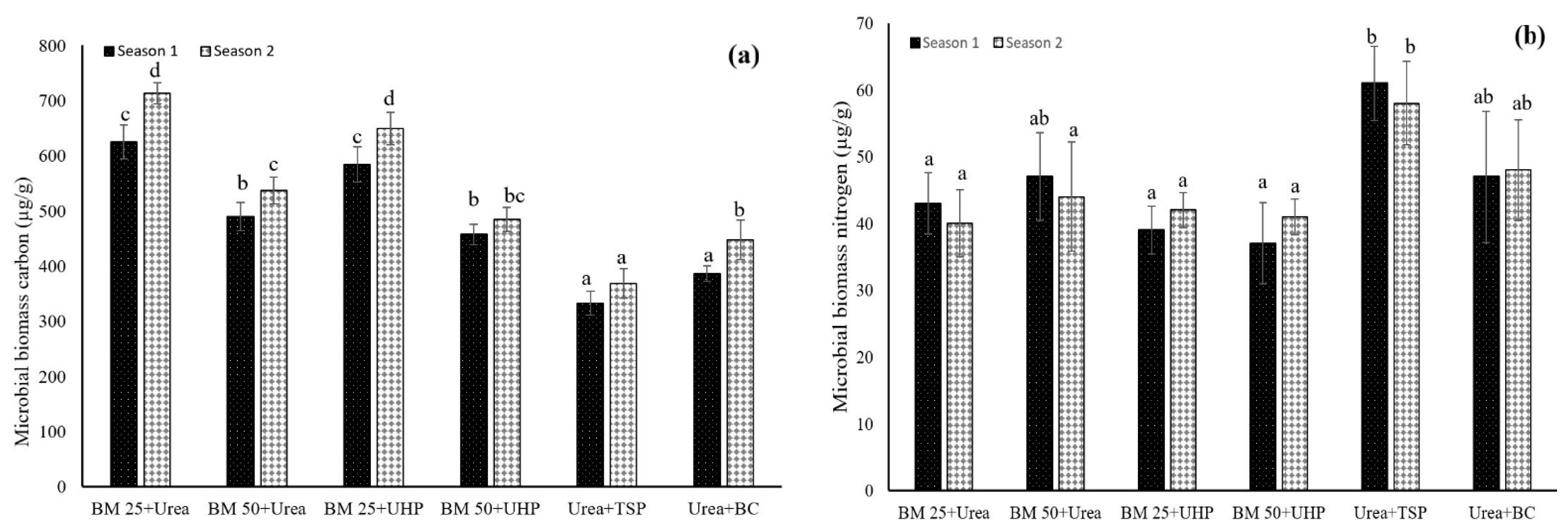
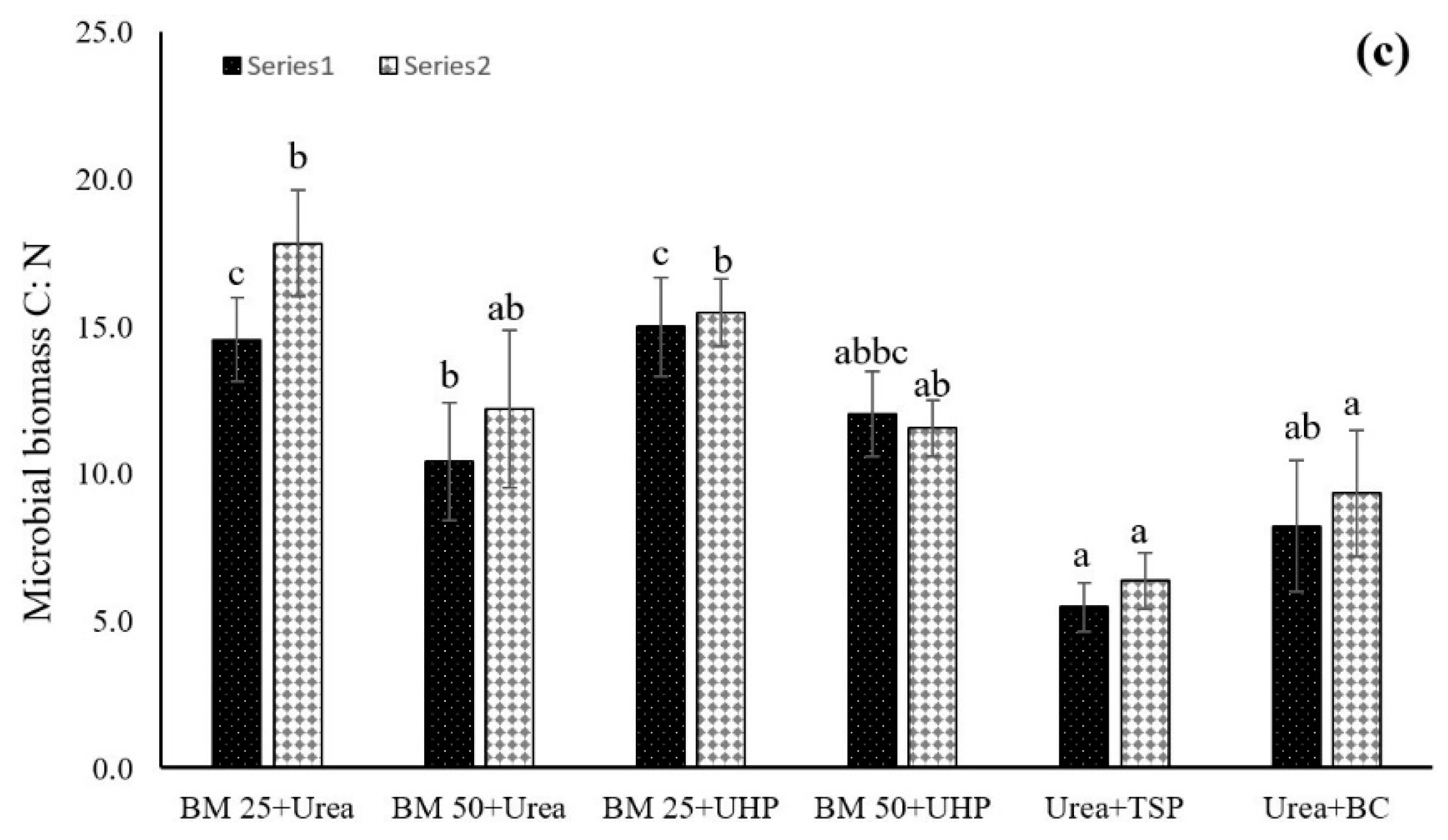
3.3. Dynamics of Soil Microbial Biomass C, N, and C:N Ratios
4. Discussions
4.1. Dynamics of Nitrogen and Phosphorus in Both the Leaf Lettuce and Soil
4.2. Dynamics of Other Macronutrients and Micronutrients in Both the Leaf Lettuce and Soil
4.3. Dynamics of Soil Microbial Biomass Carbon, Nitrogen, and C:N Ratios
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Osman, K.T. Sandy Soils. In Management of Soil Problems; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–65. [Google Scholar] [CrossRef]
- Vinh, N. Coastal sandy soils and constraints for crops in Binh Thuan Province, Southern Central Vietnam. In Proceedings of the International Management of Tropical Sandy Soils for Sustainable Agriculture Symposium, Khon Khaen, Thailand, 28 November–2 December 2005. [Google Scholar]
- Lu, C.; Tian, H. Global nitrogen and phosphorus fertilizer use for agriculture production in the past half century: Shifted hot spots and nutrient imbalance. Earth Syst. Sci. Data 2017, 9, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.; Hussain, Q.; Khan, K.S.; Alwabel, M.I.; Hayat, R.; Akmal, M.; Ijaz, S.S.; Alvi, S.; Rehman, O.U. Carbon-Based Slow-Release Fertilizers for Efficient Nutrient Management: Synthesis, Applications, and Future Research Needs. J. Soil Sci. Plant Nutr. 2021, 21, 1–26. [Google Scholar] [CrossRef]
- Lammel, J. Cost of the different options available to the farmers: Current situation and prospects. In IFA International Workshop on Enhanced-Efficiency Fertilizers, Frankfurt; International Fertilizer Industry Association: Paris, France, 2005. [Google Scholar]
- Azeem, B.; KuShaari, K.; Man, Z.B.; Basit, A.; Thanh, T.H. Review on materials & methods to produce controlled release coated urea fertilizer. J. Control. Release 2014, 181, 11–21. [Google Scholar] [CrossRef]
- Luyima, D.; Sung, J.; Lee, J.-H.; Woo, S.-A.; Park, S.-J.; Oh, T.-K. Sorption of urea hydrogen peroxide by co-pyrolysed bone meal and cow dung slowed-down phosphorus and nitrogen releases but boosted agronomic efficiency. Appl. Biol. Chem. 2020, 63, 1–11. [Google Scholar] [CrossRef]
- Hubbe, M.; Nazhad, M.; Sanchez, C. Cellulosic biomass and organic waste into high value soil amendments: A review. BioResources 2010, 5, 2808–2854. [Google Scholar] [CrossRef]
- Xie, L.; Liu, M.; Ni, B.; Zhang, X.; Wang, Y. Slow-release nitrogen and boron fertilizer from a functional superabsorbent formulation based on wheat straw and attapulgite. Chem. Eng. J. 2011, 167, 342–348. [Google Scholar] [CrossRef]
- Liu, J.; Su, Y.; Li, Q.; Yue, Q.; Gao, B. Preparation of wheat straw based superabsorbent resins and their applications as adsorbents for ammonium and phosphate removal. Bioresour. Technol. 2013, 143, 32–39. [Google Scholar] [CrossRef]
- Calabi-Floody, M.; Medina, J.; Rumpel, C.; Condron, L.M.; Hernandez, M.; Dumont, M.; De la Luz Mora, M. Smart Fertilizers as a Strategy for Sustainable Agriculture. Adv. Agron. 2018, 147, 119–157. [Google Scholar] [CrossRef]
- Luyima, D.; Lee, J.-H.; Sung, J.; Oh, T.-K. Co-pyrolysed animal manure and bone meal-based urea hydrogen peroxide (UHP) fertilisers are an effective technique of combating ammonia emissions. J. Mater. Cycles Waste Manag. 2020, 22, 1887–1898. [Google Scholar] [CrossRef]
- Zhao, L.; Cao, X.; Zheng, W.; Scott, J.W.; Sharma, B.K.; Chen, X. Co pyrolysis of biomass with phosphate fertilizers to improve biochar carbon retention, slow nutrient release, and stabilize heavy metals in soil. ACS Sustain. Chem. Eng. 2016, 4, 1630–1636. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Espie, P.; Ridgway, H. Bioactive carbon improves nitrogen fertiliser efficiency and ecological sustainability. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Luyima, D.; Egyir, M.; Lee, J.-H.; Yoo, J.-H.; Oh, T.-K. A review of the potentiality of biochar technology to abate emissions of particulate matter originating from agriculture. Int. J. Environ. Sci. Technol. 2021, 1–18. [Google Scholar] [CrossRef]
- Fairuz, D.M.; Se, S.-M.; Shaaban, A.; Hashim, M. Urea Impregnated Biochar to Minimize Nutrients Loss in Paddy Soils. Int. J. Automot. Mech. Eng. 2014, 10, 2016–2024. [Google Scholar] [CrossRef]
- Atkinson, J.H.; Giles, R.G.; Desjardins, J.G. Trace Element Content of Farmyard Manure. Can. J. Agric. Sci. 1954, 34, 76–80. [Google Scholar] [CrossRef]
- Möller, K. Assessment of Alternative Phosphorus Fertilizers for Organic Farming: Meat and Bone Meal. IMPROVE-P Factsheet 2015. Available online: https://orgprints.org/29505/1/moeller2015-factsheet-Meat-and-bone-meal.pdf (accessed on 20 May 2021).
- Güereña, D.; Lehmann, J.; Hanley, K.; Enders, A.; Hyland, C.; Riha, S. Nitrogen dynamics following field application of biochar in a temperate North American maize-based production system. Plant Soil 2012, 365, 239–254. [Google Scholar] [CrossRef]
- Biederman, L.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2012, 5, 202–214. [Google Scholar] [CrossRef]
- Karimi, A.; Moezzi, A.; Chorom, M.; Enayatizamir, N. Application of Biochar Changed the Status of Nutrients and Biological Activity in a Calcareous Soil. J. Soil Sci. Plant Nutr. 2019, 20, 450–459. [Google Scholar] [CrossRef]
- Kalra, Y.P. Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Wolf, B. The determination of boron in soil extracts, plant materials, composts, manures, water and nutrient solutions. Commun. Soil Sci. Plant Anal. 1971, 2, 363–374. [Google Scholar] [CrossRef]
- Jones, J.B., Jr. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Fontes, R.L.; Dallpai, D.L.; Braga, J.M.; Alvarez, V.V.H. Determination of molybdenum in soil test extracts with potassium iodide plus hydrogen peroxide reaction. Commun. Soil Sci. Plant. Anal. 2000, 31, 2671–2683. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method forthe deter-mination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Gasparatos, D.; Haidouti, C. A comparison of wet oxidation methods for determination of total phosphorus in soils. J. Plant Nutr. Soil Sci. 2001, 164, 435–439. [Google Scholar] [CrossRef]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.; Pommerening, B.; Chaussod, R.; Brookes, P. Measurement of soil microbial biomass C by fumigation-extraction—an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Zhang, Q.-Z.; Dijkstra, F.A.; Liu, X.-R.; Wang, Y.-D.; Huang, J.; Lu, N. Effects of Biochar on Soil Microbial Biomass after Four Years of Consecutive Application in the North China Plain. PLoS ONE 2014, 9, e102062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.-H.; Luyima, D.; Lee, J.-H.; Park, S.-Y.; Yang, J.-W.; An, J.-Y.; Yun, Y.-U.; Oh, T.-K. Effects of brewer’s spent grain biochar on the growth and quality of leaf lettuce (Lactuca sativa L. var. crispa.). Appl. Biol. Chem. 2021, 64, 1–10. [Google Scholar] [CrossRef]
- Siebers, N.F.; Godlinski, F.; Leinweber, P. The phosphorus fertilizer value of bone char for potatoes, wheat and onions: First results. Landbauforsch Volk. 2012, 62, 59–64. [Google Scholar]
- Zwetsloot, M.J.; Lehmann, J.; Bauerle, T.; Vanek, S.; Hestrin, R.; Nigussie, A. Phosphorus availability from bone char in a P-fixing soil influenced by root-mycorrhizae-biochar interactions. Plant Soil 2016, 408, 95–105. [Google Scholar] [CrossRef]
- Lustosa Filho, J.F.; Penido, E.S.; Castro, P.P.; Silva, C.A.; Melo, L.C.A. Co Pyroly sis of poultry litter and phosphate and magnesium generates alternative slow release fertilizer suitable for tropical soils. ACS Sustain. Chem. Eng. 2017, 5, 9043–9052. [Google Scholar] [CrossRef]
- Ismail, A.M.; Heuer, S.; Thomson, M.; Wissuwa, M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef]
- Li, J.; Xie, Y.; Dai, A.; Liu, L.; Li, Z. Root and shoot traits responses to phosphorus deficiency and QTL analysis at seedling stage using introgression lines of rice. J. Genet. Genom. 2009, 36, 173–183. [Google Scholar] [CrossRef]
- Wacker-Fester, K.; Uptmoor, R.; Pfahler, V.; Dehmer, K.J.; Bachmann-Pfabe, S.; Kavka, M. Genotype-Specific Differences in Phosphorus Efficiency of Potato (Solanum tuberosum L.). Front. Plant Sci. 2019, 10, 1029. [Google Scholar] [CrossRef] [Green Version]
- Kucey, R.; Janzen, H.; Leggett, M. Microbially Mediated Increases in Plant-Available Phosphorus. Adv. Agron. 1989, 42, 199–228. [Google Scholar] [CrossRef]
- Øgaard, A.F. Relationships between the Ratio of Plant-Available Phosphorus (P-AL) to Total Phosphorus and Soil Properties. Acta Agric. Scand. Sect. B Plant Soil Sci. 1994, 44, 136–141. [Google Scholar] [CrossRef]
- Shen, P.; He, X.-H.; Xu, M.-G.; Zhang, H.-M.; Peng, C.; Gao, H.-J.; Liu, H.; Xu, Y.-M.; Qin, S.; Xiao, H.-J. Soil Organic Carbon Accumulation Increases Percentage of Soil Olsen-P to Total P at Two 15-Year Mono-Cropping Systems in Northern China. J. Integr. Agric. 2014, 13, 597–603. [Google Scholar] [CrossRef]
- Utomo, W.H.; Islami, T.; Wisnubroto, E.; Soelistyari, H.T. Biochar as a carrier for nitrogen plant nutrition: 3. Effect of enriched biochar on rice (Oryza sativa L.) yield and soil qualities. Int. J. Appl. Eng. Res. 2017, 12, 10426–10432. [Google Scholar]
- Shi, W.; Ju, Y.; Bian, R.; Li, L.; Joseph, S.; Mitchell, D.R.; Munroe, P.; Taherymoosavi, S.; Pan, G. Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci. Total Environ. 2019, 701, 134424. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C.C.; Rahaman, A.; Robles-Aguilar, A.A.; Latif, S.; Intani, K.; Müller, J.; Jablonowski, N.D. Nutrient Loaded Biochar Doubled Biomass Production in Juvenile Maize Plants (Zea mays L.). Agronomy 2020, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Clough, T.J.; Condron, L.M.; Kammann, C.; Müller, C. A Review of Biochar and Soil Nitrogen Dynamics. Agronomy 2013, 3, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Haider, G.; Steffens, D.; Moser, G.; Müller, C.; Kammann, C.I. Biochar reduced nitrate leaching and improved soil moisture content without yield improvements in a four-year field study. Agric. Ecosyst. Environ. 2017, 237, 80–94. [Google Scholar] [CrossRef]
- Hagemann, N.; Kammann, C.I.; Schmidt, H.-P.; Kappler, A.; Behrens, S. Nitrate capture and slow release in biochar amended compost and soil. PLoS ONE 2017, 12, e0171214. [Google Scholar] [CrossRef]
- Deenik, J.L.; McClellan, T.; Uehara, G.; Antal, M.J.; Campbell, S. Charcoal Volatile Matter Content Influences Plant Growth and Soil Nitrogen Transformations. Soil Sci. Soc. Am. J. 2010, 74, 1259–1270. [Google Scholar] [CrossRef]
- O’Toole, A.; De Zarruk, K.K.; Steffens, M.; Rasse, D.P. Characterization, Stability, and Plant Effects of Kiln-Produced Wheat Straw Biochar. J. Environ. Qual. 2013, 42, 429–436. [Google Scholar] [CrossRef]
- Ianni, A.; Martino, C.; Innosa, D.; Bennato, F.; Grotta, L.; Martino, G. Zinc supplementation of lactating dairy cows: Effects on chemical-nutritional quality and volatile profile of Caciocavallo cheese. Asian Australas. J. Anim. Sci. 2020, 33, 825–835. [Google Scholar] [CrossRef] [Green Version]
- Olszyk, D.; Shiroyama, T.; Novak, J.; Cantrell, K.; Sigua, G.; Watts, D.; Johnson, M.G. Biochar affects growth and shoot nitrogen in four crops for two soils. Agrosyst. Geosci. Environ. 2020, 3. [Google Scholar] [CrossRef]
- Gartler, J.; Robinson, B.; Burton, K.; Clucas, L. Carbonaceous soil amendments to biofortify crop plants with zinc. Sci. Total Environ. 2013, 465, 308–313. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Taşkin, M.B.; Sahin, O.; Kaya, E.C.; Atakol, A. Effect of phosphorus-enriched biochar and poultry manure on growth and mineral composition of lettuce (Lactuca sativaL. cv.) grown in alkaline soil. Soil Use Manag. 2014, 30, 182–188. [Google Scholar] [CrossRef]
- Rees, F.; Germain, C.; Sterckeman, T.; Morel, J.-L. Plant growth and metal uptake by a non-hyperaccumulating species (Lolium perenne) and a Cd-Zn hyperaccumulator (Noccaea caerulescens) in contaminated soils amended with biochar. Plant. Soil 2015, 395, 57–73. [Google Scholar] [CrossRef]
- Syuhada, A.; Shamshuddin, J.; Fauziah, C.; Rosenani, A.; Arifin, A. Biochar as soil amendment: Impact on chemical properties and corn nutrient uptake in a Podzol. Can. J. Soil Sci. 2016, 96, 400–412. [Google Scholar] [CrossRef]
- Sorrenti, G.; Masiello, C.; Toselli, M. Biochar interferes with kiwifruit Fe-nutrition in calcareous soil. Geoderma 2016, 272, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Ramzani, P.M.A.; Shan, L.; Anjum, S.; Khan, W.-U.; Ronggui, H.; Iqbal, M.; Virk, Z.A.; Kausar, S. Improved quinoa growth, physiological response, and seed nutritional quality in three soils having different stresses by the application of acidified biochar and compost. Plant. Physiol. Biochem. 2017, 116, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Gaskin, J.W.; Speir, R.A.; Harris, K.; Das, K.C.; Lee, R.D.; Morris, L.A.; Fisher, D.S. Effect of Peanut Hull and Pine Chip Biochar on Soil Nutrients, Corn Nutrient Status, and Yield. Agron. J. 2010, 102, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Moradi, S.; Rasouli-Sadaghiani, M.H.; Sepehr, E.; Khodaverdiloo, H.; Barin, M. Soil nutrients status affected by simple and enriched biochar application under salinity conditions. Environ. Monit. Assess. 2019, 191, 257. [Google Scholar] [CrossRef]
- Kizito, S.; Luo, H.; Lu, J.; Bah, H.; Dong, R.; Wu, S. Role of Nutrient-Enriched Biochar as a Soil Amendment during Maize Growth: Exploring Practical Alternatives to Recycle Agricultural Residuals and to Reduce Chemical Fertilizer Demand. Sustainability 2019, 11, 3211. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Bai, Y.; Lv, M.; Tian, G.; Zhang, X.; Li, L.; Jiang, Y.; Ge, S. Soil Fertility, Microbial Biomass, and Microbial Functional Diversity Responses to Four Years Fertilization in an Apple Orchard in North China. Hortic. Plant. J. 2020, 6, 223–230. [Google Scholar] [CrossRef]
- Castaldi, S.; Riondino, M.; Baronti, S.; Esposito, F.; Marzaioli, R.; Rutigliano, F.; Vaccari, F.; Miglietta, F. Impact of biochar application to a Mediterranean wheat crop on soil microbial activity and greenhouse gas fluxes. Chemosphere 2011, 85, 1464–1471. [Google Scholar] [CrossRef]
- Zavalloni, C.; Alberti, G.; Biasiol, S.; Vedove, G.D.; Fornasier, F.; Liu, J.; Peressotti, A. Microbial mineralization of biochar and wheat straw mixture in soil: A short-term study. Appl. Soil Ecol. 2011, 50, 45–51. [Google Scholar] [CrossRef]
- Luo, Y.; Durenkamp, M.; De Nobili, M.; Lin, Q.; Devonshire, B.; Brookes, P. Microbial biomass growth, following incorporation of biochars produced at 350 °C or 700 °C, in a silty-clay loam soil of high and low pH. Soil Biol. Biochem. 2013, 57, 513–523. [Google Scholar] [CrossRef]
- Du, Z.; Xiao, Y.; Qi, X.; Liu, Y.; Fan, X.; Li, Z. Peanut-Shell Biochar and Biogas Slurry Improve Soil Properties in the North China Plain: A Four-Year Field Study. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Chen, J.; Pan, G.; Liu, X.; Zhang, X.; Li, L.; Bian, R.; Cheng, K.; Jinwei, Z. Biochar decreased microbial metabolic quotient and shifted community composition four years after a single incorporation in a slightly acid rice paddy from southwest China. Sci. Total Environ. 2016, 571, 206–217. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luyima, D.; Egyir, M.; Yun, Y.-U.; Park, S.-J.; Oh, T.-K. Nutrient Dynamics in Sandy Soil and Leaf Lettuce Following the Application of Urea and Urea-Hydrogen Peroxide Impregnated Co-Pyrolyzed Animal Manure and Bone Meal. Agronomy 2021, 11, 1664. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081664
Luyima D, Egyir M, Yun Y-U, Park S-J, Oh T-K. Nutrient Dynamics in Sandy Soil and Leaf Lettuce Following the Application of Urea and Urea-Hydrogen Peroxide Impregnated Co-Pyrolyzed Animal Manure and Bone Meal. Agronomy. 2021; 11(8):1664. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081664
Chicago/Turabian StyleLuyima, Deogratius, Michael Egyir, Yeo-Uk Yun, Seong-Jin Park, and Taek-Keun Oh. 2021. "Nutrient Dynamics in Sandy Soil and Leaf Lettuce Following the Application of Urea and Urea-Hydrogen Peroxide Impregnated Co-Pyrolyzed Animal Manure and Bone Meal" Agronomy 11, no. 8: 1664. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081664