
The Genetic Control of Stomatal Development in Barley: New Solutions for Enhanced Water-Use Efficiency in Drought-Prone Environments



Abstract
:1. Introduction
2. The Genetic Control of Stomatal Density Improves WUE

{kind=link}
{kind=link}
| Type of Stomatal Modification | Plant Response | Study Species | Citation |
|---|---|---|---|
| Increased density | Enhanced CO2 assimilation capacity. | Mosses | [64] |
| Increase in gs, no impact on photosynthetic efficiency or CO2 assimilation rate. | Arabidopsis | [65] | |
| No impact on photosynthetic efficiency. | Rice | [66] | |
| Clustered stomata | Impaired CO2 diffusion, no impact on photosynthetic efficiency. | Arabidopsis | [65] |
| Reduced aperture | Increased WUE, decrease in gs. | Arabidopsis | [67] |
| Reduced density | Increased WUE, decrease in gs, reduced decrease in grain yield under water restriction (29.68%) versus higher stomatal density varieties (33.57%). | Wheat | [68] |
| Increased WUE, enhanced survivorship and evaporative cooling in drought and high temperature (40 °C) versus controls, 27% increase in yield versus controls under drought conditions. | Rice | [48] | |
| Increased WUE, stable yield under water-restriction (<50% reduction in stomatal density only). | Wheat | [58] | |
| Increased WUE, Decrease in gs, minimal reduction in CO2 assimilation, barley lines with 88% reductions in stomatal density produce yields comparable to controls grown under well-watered conditions even under water-restricted conditions. | Barley | [62] | |
| Increased WUE, 80% higher survival rate under drought (versus control plants), reduced leaf water loss, no impact to photosynthetic efficiency under well-watered conditions, higher photosynthetic rate than controls under drought conditions. | Maize | [69] | |
| Increased WUE, no impact on photosynthetic efficiency, no difference in fruit yield versus controls under both well-watered and water restricted conditions, enhanced dehydration tolerance. | Tomato | [70,71] |
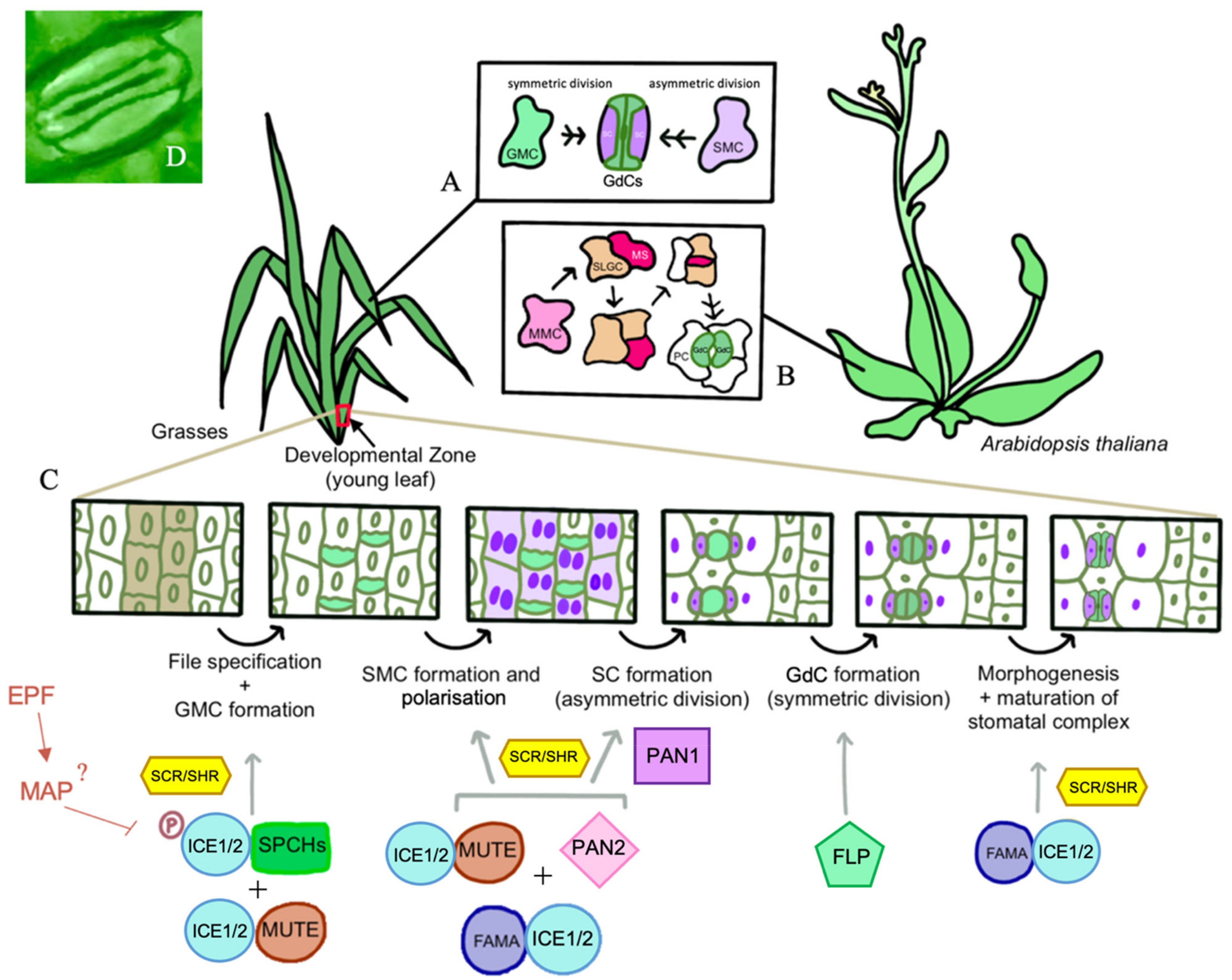
3. Complex Gene Networks Direct Stomatal Development
4. Stomatal Development in the Dicotyledons versus the Grasses
5. Climate Resilient Cereal Crops through Genetic Improvement Using Barley as an Example
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naumann, G.; Alfieri, L.; Wyser, K.; Mentaschi, L.; Betts, R.A.; Carrao, H.; Spinoni, J.; Vogt, J.; Feyen, L. Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 2018, 45, 3285–3296. [Google Scholar] [CrossRef]
- Kirono, D.G.C.; Round, V.; Heady, C.; Chiew, F.H.S.; Osbrough, S. Drought projections for Australia: Updated results and analysis of model simulations. Weather Clim. Extrem. 2020, 30, 100280. [Google Scholar] [CrossRef]
- Markonis, Y.; Kumar, R.; Hanel, M.; Rakovec, O.; Máca, P.; AghaKouchak, A. The rise of compound warm-season droughts in Europe. Sci. Adv. 2021, 7, 9668. [Google Scholar] [CrossRef]
- Stahl, K.; Kohn, I.; Blauhut, V.; Urquijo, J.; De Stefano, L.; Acácio, V.; Dias, S.; Stagge, J.H.; Tallaksen, L.M.; Kampragou, E.; et al. Impacts of European drought events: Insights from an international database of text-based reports. Nat. Hazards Earth Syst. Sci. 2016, 16, 801–819. [Google Scholar] [CrossRef] [Green Version]
- Ekwurzel, B.; Boneham, J.; Dalton, M.W.; Heede, R.; Mera, R.J.; Allen, M.R.; Frumhoff, P.C. The rise in global atmospheric CO2, surface temperature, and sea level from emissions traced to major carbon producers. Clim. Chang. 2017, 144, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Venturas, M.D.; Todd, H.N.; Trugman, A.T.; Anderegg, W.R.L.; Wang, Y.; Tai, X. The impact of rising CO2 and acclimation on the response of US forests to global warming. Proc. Natl. Acad. Sci. USA 2019, 116, 25734–25744. [Google Scholar] [CrossRef]
- Crawford, A.J.; McLachlan, D.H.; Hetherington, A.M.; Franklin, K.A. High temperature exposure increases plant cooling capacity. Curr. Biol. 2012, 22, 396–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Azorin-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 044001. [Google Scholar] [CrossRef]
- Climate Council of Australia. Compound Costs: How Climate Change is Damaging Australia’s Economy; Climate Council of Australia Limited: Melbourne, Australia, 2019; p. 2. [Google Scholar]
- Intergovernmental Panel on Climate Change. Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Sultan, B.; Defrance, D.; Izumi, T. Evidence of crop production losses in West Africa due to historical global warming in two crop models. Sci. Rep. 2019, 9, 12834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, T.P.; Perryman, A.H.; Osborne, T.M. Modelling impacts of climate change on global food security. Clim. Chang. 2016, 134, 429–440. [Google Scholar] [CrossRef]
- Jha, P.K.; Kumar, S.N.; Ines, A.V. Responses of soybean to water stress and supplemental irrigation in upper Indo-Gangetic plain: Field experiment and modelling approach. Field Crop. Res. 2018, 219, 76–86. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.; Bota, J. From leaf to whole-plant water-use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.; Soh, W.K.; Yiotis, C.; Batke, S.; Parnell, A.C.; Spicer, R.A.; Lawson, T.; Caballero, R.; Wright, I.J.; Purcell, C.; et al. Convergence in maximum stomatal conductance of C3 woody angiosperms in natural ecosystems across bioclimatic zones. Front. Plant Sci. 2019, 10, 558. [Google Scholar] [CrossRef] [Green Version]
- Roberts, B. What are stomates and how do they work? J. Arboric. 1990, 16, 331. [Google Scholar]
- Rudall, P.J.; Chen, E.D.; Cullen, E. Evolution and development of monocot stomata. Am. J. Bot. 2017, 104, 1122–1141. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Griffiths, R.; Ying, J.; McCourt, P.; Huang, Y. Development of drought-tolerant canola (Brassica napus L.) through genetic modulation of ABA-mediated stomatal responses. Crop Sci. 2009, 49, 1539–1554. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Wang, H.; Liu, S.; Jia, F.; Su, Y.; Li, S.; He, F.; Feng, C.; Niu, M.; et al. The receptor-like kinase ERECTA confers improved water-use efficiency and drought tolerance to poplar via modulating stomatal density. Int. J. Mol. Sci. 2021, 22, 7245. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Beerling, D.J.; Chaloner, W.G. The impact of atmospheric CO2 and temperature changes on stomatal density: Observation from Quercus robur Lammas leaves. Ann. Bot. 1993, 71, 231–235. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Influence of different light intensities on specific leaf weight, stomatal density photosynthesis and seed yield in soybean. Plant Physiol. Rep. 2020, 25, 277–283. [Google Scholar] [CrossRef]
- Pillitteri, L.J.; Dong, J. Stomatal development in Arabidopsis. Arab. Book 2013, 11, e0162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of stomatal Density and morphology on water-use efficiency in a changing world. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupu, A.; Nevo, E.; Zamorzaeva, I.; Korol, A. Ecological–genetic feedback in DNA repair in wild barley, Hordeum spontaneum. Genetica 2006, 127, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ren, X.; Sun, D.; Sun, G. Origin of worldwide cultivated barley revealed by NAM-1 gene and grain protein content. Front. Plant Sci. 2015, 6, 803. [Google Scholar] [CrossRef] [Green Version]
- Abbo, S.; Lev-Yadun, S.; Gopher, A. Plant domestication and crop evolution in the near east: On events and processes. Crit. Rev. Plant Sci. 2012, 31, 241–257. [Google Scholar] [CrossRef]
- Sato, K.; Yamane, M.; Yamaji, N.; Kanamori, H.; Tagiri, A.; Schwerdt, J.G.; Fincher, G.B.; Matsumoto, T.; Takeda, K.; Komatsuda, T. Alanine aminotransferase controls seed dormancy in barley. Nat. Commun. 2016, 7, 11625. [Google Scholar] [CrossRef]
- Langridge, P. Economic and Academic Importance of Barley. In The Barley Genome; Stein, N., Muehlbauer, G., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–10. [Google Scholar]
- Department of Health and Ageing Office of the Gene Technology Regulator. The Biology of Hordeum vulgare L. (Barley); Office of the Gene Technology Regulator: Canberra, Australia, 2008; pp. 1–10. [Google Scholar]
- Australian Bureau of Statistics. Value of Agricultural Commodities Produced, Australia; Australian Bureau of Statistics: Canberra, Australia, 2019.
- Australian Export Grains Innovation Centre. Australian Barley: Quality, Safety and Reliability; Department of Primary Industries and Regional Development: Perth, Australia, 2018; pp. 1–4. [Google Scholar]
- Cossani, C.M.; Slafer, G.A.; Savin, R. Do barley and wheat (bread and durum) differ in grain weight stability through seasons and water–nitrogen treatments in a Mediterranean location? Field Crop. Res. 2011, 121, 240–247. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Zhang, G.; Wu, F. Exploration and Utilization of Drought-Tolerant Barley Germplasm. In Exploration, Identification and Utilization of Barley Germplasm; Li, C., Zhang, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 115–152. [Google Scholar]
- Liu, W.; Feng, X.; Cao, F.; Wu, D.; Zhang, G.; Vincze, E.; Wang, Y.; Chen, Z.; Wu, F. An ATP binding cassette transporter HvABCB25 confers aluminum detoxification in wild barley. J. Hazard. Mater. 2020, 401, 123371. [Google Scholar] [CrossRef]
- D’Alpoim Guedes, J.A.; Lu, H.; Hein, A.M.; Schmidt, A.H. Early evidence for the use of wheat and barley as staple crops on the margins of the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2015, 112, 5625–5630. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Monem, A.; El-Habbasha, S.; Hozayn, M. Mitigation salinity stress effects on barley (Hordeum vulgare L.) growth, yield and some physiological aspects. J. Appl. Sci. Res. 2013, 9, 2411–2421. [Google Scholar]
- Kumar, A.; Verma, R.P.S.; Singh, A.; Kumar Sharma, H.; Devi, G. Barley landraces: Ecological heritage for edaphic stress adaptations and sustainable production. Environ. Sustain. Indic. 2020, 6, 1035. [Google Scholar] [CrossRef]
- Mwando, E.; Angessa, T.T.; Han, Y.; Li, C. Salinity tolerance in barley during germination—Homologs and potential genes. J. Zhejiang Univ. Sci. B 2020, 21, 93–121. [Google Scholar] [CrossRef] [PubMed]
- Gürel, F.; Öztürk, Z.N.; Uçarlı, C.; Rosellini, D. Barley genes as tools to confer abiotic stress tolerance in crops. Front. Plant Sci. 2016, 7, 1137. [Google Scholar] [CrossRef] [PubMed]
- Vafadar Shamasbi, F.; Jamali, S.; Sadeghzadeh, B.; Abdollahi Mandoulakani, B. Genetic mapping of quantitative trait loci for yield-affecting traits in a barley doubled haploid population derived from clipper × sahara 3771. Front. Plant Sci. 2017, 8, 688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chater, C.C.; Caine, R.S.; Fleming, A.J.; Gray, J.E. Origins and evolution of stomatal development. Plant Physiol. 2017, 174, 624–638. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Chen, F.; Qi, S.; Zhang, L.; Wu, S. Loss or duplication of key regulatory genes coincides with environmental adaptation of the stomatal complex in Nymphaea colorata and Kalanchoe laxiflora. Hortic. Res. 2018, 5, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, K.M.; Rychel, A.L.; Torii, K.U. Out of the mouths of plants: The molecular basis of the evolution and diversity of stomatal development. Plant Cell 2010, 22, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [Green Version]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water-use efficiency. Plant Physiol. 2014, 164, 1556–1570. [Google Scholar] [CrossRef] [Green Version]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2018, 221, 371–384. [Google Scholar] [CrossRef]
- Mohammed, U.; Caine, R.S.; Atkinson, J.A.; Harrison, E.L.; Wells, D.; Chater, C.C.; Gray, J.E.; Swarup, R.; Murchie, E.H. Rice plants overexpressing OsEPF1 show reduced stomatal density and increased root cortical aerenchyma formation. Sci. Rep. 2019, 9, 5584. [Google Scholar] [CrossRef] [Green Version]
- Patz, J.A.; Frumkin, H.; Holloway, T.; Vimont, D.J.; Haines, A. Climate Change. J. Am. Med. Assoc. 2014, 312, 1565. [Google Scholar] [CrossRef]
- Grantz, D.A.; Zeiger, E. Stomatal responses to light and leaf-air water vapor pressure difference show similar kinetics in sugarcane and soybean. Plant Physiol. 1986, 81, 865–868. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomata and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, D.; Pradhan, G.C. Effect of transpiration-reducing chemicals on growth, flowering, and stomatal opening of tomato plants. Plant Physiol. 1972, 50, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Doheny-Adams, T.; Hunt, L.; Franks, P.J.; Beerling, D.J.; Gray, J.E. Genetic manipulation of stomatal density influences stomatal size, plant growth and tolerance to restricted water supply across a growth carbon dioxide gradient. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 547–555. [Google Scholar] [CrossRef]
- Dittberner, H.; Korte, A.; Mettler-Altmann, T.; Weber, A.P.M.; Monroe, G.; de Meaux, J. Natural variation in stomata size contributes to the local adaptation of water-use efficiency in Arabidopsis thaliana. Mol. Ecol. 2018, 27, 4052–4065. [Google Scholar] [CrossRef] [Green Version]
- Von Caemmerer, S.; Evans, J.R. Temperature responses of mesophyll conductance differ greatly between species. Plant Cell Environ. 2014, 38, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U.F.; McMillan, A.M.S. Warming and elevated CO2 have opposing influences on transpiration. Which is more important? Curr. For. Rep. 2018, 4, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Dunn, J.; Hunt, L.; Afsharinafar, M.; Meselmani, M.A.; Mitchell, A.; Howells, R.; Wallington, E.; Fleming, A.J.; Gray, J.E. Reduced stomatal density in bread wheat leads to increased water-use efficiency. J. Exp. Bot. 2019, 70, 4737–4748. [Google Scholar] [CrossRef] [PubMed]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Climatol. 2017, 38, 1718–1736. [Google Scholar] [CrossRef] [Green Version]
- Coupel-Ledru, A.; Lebon, E.; Christophe, A.; Gallo, A.; Gago, P.; Pantin, F.; Doligez, A.; Simonneau, T. Reduced nighttime transpiration is a relevant breeding target for high water-use efficiency in grapevine. Proc. Natl. Acad. Sci. USA 2016, 113, 8963–8968. [Google Scholar] [CrossRef] [Green Version]
- Büssis, D.; von Groll, U.; Fisahn, J.; Altmann, T. Stomatal aperture can compensate altered stomatal density in Arabidopsis thaliana at growth light conditions. Funct. Plant Biol. 2006, 33, 1037. [Google Scholar] [CrossRef]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing stomatal density in barley improves drought tolerance without impacting on yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef] [Green Version]
- Franks, P.J.; Doheny-Adams, T.W.; Britton-Harper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef]
- Kubásek, J.; Hájek, T.; Duckett, J.; Pressel, S.; Šantrůček, J. Moss stomata do not respond to light and CO2 concentration but facilitate carbon uptake by sporophytes: A gas exchange, stomatal aperture, and 13 C labelling study. New Phytol. 2021, 230, 1815–1828. [Google Scholar] [CrossRef]
- Vráblová, M.; Vrábl, D.; Hronková, M.; Kubásek, J.; Šantrůček, J. Stomatal function, density and pattern, and CO2 assimilation in Arabidopsis thaliana tmm1 and sdd1-1 mutants. Plant Biol. 2017, 19, 689–701. [Google Scholar] [CrossRef]
- Schuler, M.L.; Sedelnikova, O.V.; Walker, B.J.; Westhoff, P.; Langdale, J.A. SHORTROOT-mediated increase in stomatal density has no impact on photosynthetic efficiency. Plant Physiol. 2017, 176, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Long, Y.; Feng, X.; Zhu, X.; Sai, N.; Chirkova, L.; Betts, A.; Herrmann, J.; Edwards, E.J.; Okamoto, M.; et al. GABA signalling modulates stomatal opening to enhance plant water use efficiency and drought resilience. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Li, Y.; Zhang, S. Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. Crop J. 2017, 5, 231–239. [Google Scholar] [CrossRef]
- Liu, Y.; Qin, L.; Han, L.; Xiang, Y.; Zhao, D. Overexpression of maize SDD1 (ZmSDD1) improves drought resistance in Zea mays L. by reducing stomatal density. Plant Cell Tissue Organ Cult. 2015, 122, 147–159. [Google Scholar] [CrossRef]
- Li, S.; Zhang, J.; Liu, L.; Wang, Z.; Li, Y.; Guo, L.; Li, Y.; Zhang, X.; Ren, S.; Zhao, B.; et al. SlTLFP8 reduces water loss to improve water-use efficiency by modulating cell size and stomatal density via endoreduplication. Plant Cell Environ. 2020, 43, 2666–2679. [Google Scholar] [CrossRef]
- Morales-Navarro, S.; Pérez-Díaz, R.; Ortega, A.; de Marcos, A.; Mena, M.; Fenoll, C.; González-Villanueva, E.; Ruiz-Lara, S. Overexpression of a SDD1-like gene From wild tomato decreases stomatal density and enhances dehydration avoidance in Arabidopsis and cultivated tomato. Front. Plant Sci. 2018, 9, 940. [Google Scholar] [CrossRef]
- Davenport, D.C.; Fisher, M.A.; Hagan, R.M. Some counteractive effects of antitranspirants. Plant Physiol. 1972, 49, 722–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinn, G.; Brummett, D.L. Leaf age, decline in photosynthesis, and changes in abscisic acid, indole-3-acetic acid, and cytokinin in cotton leaves. Field Crop. Res. 1993, 32, 269–275. [Google Scholar] [CrossRef]
- Negin, B.; Yaaran, A.; Kelly, G.; Zait, Y.; Moshelion, M. Mesophyll abscisic acid restrains early growth and flowering but does not directly suppress photosynthesis. Plant Physiol. 2019, 180, 910–925. [Google Scholar] [CrossRef] [Green Version]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Seiler, C.; Harshavardhan, V.T.; Reddy, P.S.; Hensel, G.; Kumlehn, J.; Eschen-Lippold, L.; Rajesh, K.; Korzun, V.; Wobus, U.; Lee, J.; et al. Abscisic acid flux alterations result in differential abscisic acid signaling responses and impact assimilation efficiency in barley under terminal drought stress. Plant Physiol. 2014, 164, 1677–1696. [Google Scholar] [CrossRef]
- Ruggiero, A.; Punzo, P.; Landi, S.; Costa, A.; Van Oosten, M.J.; Grillo, S. Improving plant water-use efficiency through molecular genetics. Horticulturae 2017, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Yang, L.; Liu, X.; Tang, R.; Wang, Y.; Ge, H.; Wu, M.; Zhang, J.; Zhao, F.; Luan, S.; et al. Overexpression of poplar pyrabactin resistance-like abscisic acid receptors promotes abscisic acid sensitivity and drought resistance in transgenic Arabidopsis. PLoS ONE 2016, 11, e0168040. [Google Scholar] [CrossRef]
- Christiansen, M.W.; Matthewman, C.; Podzimska-Sroka, D.; O’Shea, C.; Lindemose, S.; Møllegaard, N.E.; Holme, I.B.; Hebelstrup, K.; Skriver, K.; Gregersen, P.L. Barley plants over-expressing the NAC transcription factor geneHvNAC005 show stunting and delay in development combined with early senescence. J. Exp. Bot. 2016, 67, 5259–5273. [Google Scholar] [CrossRef] [Green Version]
- Faralli, M.; Grove, I.G.; Hare, M.C.; Boyle, R.D.; Williams, K.S.; Corke, F.M.K.; Kettlewell, P.S. Canopy application of film antitranspirants over the reproductive phase enhances yield and yield-related physiological traits of water-stressed oilseed rape (Brassica napus). Crop Pasture Sci. 2016, 67, 751. [Google Scholar] [CrossRef]
- Lau, O.S.; Bergmann, D.C. Stomatal development: A plant’s perspective on cell polarity, cell fate transitions and intercellular communication. Development 2012, 139, 3683–3692. [Google Scholar] [CrossRef] [Green Version]
- MacAlister, C.A.; Ohashi-Ito, K.; Bergmann, D.C. Transcription factor control of asymmetric cell divisions that establish the stomatal lineage. Nature 2006, 445, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Zoulias, N.; Harrison, E.L.; Casson, S.A.; Gray, J.E. Molecular control of stomatal development. Biochem. J. 2018, 475, 441–454. [Google Scholar] [CrossRef] [Green Version]
- De Marcos, A.; Houbaert, A.; Triviño, M.; Delgado, D.; Martín-Trillo, M.; Russinova, E.; Fenoll, C.; Mena, M. A mutation in the bHLH domain of the SPCH transcription factor uncovers a BR-dependent mechanism for stomatal development. Plant Physiol. 2017, 174, 823–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jewaria, P.K.; Hara, T.; Tanaka, H.; Kondo, T.; Betsuyaku, S.; Sawa, S.; Sakagami, Y.; Aimoto, S.; Kakimoto, T. Differential effects of the peptides stomagen, EPF1 and EPF2 on activation of MAP kinase MPK6 and the SPCH protein level. Plant Cell Physiol. 2013, 54, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Hnilova, M.; Maes, M.; Lin, Y.L.; Putarjunan, A.; Han, S.; Avila, J.; Torii, K.U. Competitive binding of antagonistic peptides fine-tunes stomatal patterning. Nature 2015, 522, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Chen, X.; Mang, H.; Liu, C.; Yu, X.; Gao, X.; Torii, K.U.; He, P.; Shan, L. Differential function of Arabidopsis SERK family receptor-like kinases in stomatal patterning. Curr. Biol. 2015, 25, 2361–2372. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.; Michniewicz, M.; Bergmann, D.C.; Wang, Z. Brassinosteroid regulates stomatal development by GSK3-mediated inhibition of a MAPK pathway. Nature 2012, 482, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Gudesblat, G.E.; Schneider-Pizoń, J.; Betti, C.; Mayerhofer, J.; Vanhoutte, I.; van Dongen, W.; Boeren, S.; Zhiponova, M.; de Vries, S.; Jonak, C.; et al. SPEECHLESS integrates brassinosteroid and stomata signalling pathways. Nat. Cell Biol. 2012, 14, 548–554. [Google Scholar] [CrossRef]
- Serna, L. What causes opposing actions of brassinosteroids on stomatal development? Plant Physiol. 2013, 162, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Hunt, L.; Gray, J.E. The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development. Curr. Biol. 2009, 19, 864–869. [Google Scholar] [CrossRef] [Green Version]
- Vatén, A.; Soyars, C.L.; Tarr, P.T.; Nimchuk, Z.L.; Bergmann, D.C. Modulation of asymmetric division diversity through cytokinin and SPEECHLESS regulatory interactions in the Arabidopsis stomatal lineage. Dev. Cell 2018, 47, 53–66.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillitteri, L.J.; Torii, K.U. Breaking the silence: Three bHLH proteins direct cell-fate decisions during stomatal development. BioEssays 2007, 29, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.A.; Bergmann, D.C. Functional specialization of stomatal bHLHs through modification of DNA-binding and phosphoregulation potential. Proc. Natl. Acad. Sci. USA 2014, 111, 15585–15590. [Google Scholar] [CrossRef] [Green Version]
- Han, S.K.; Qi, X.; Sugihara, K.; Dang, J.H.; Endo, T.A.; Miller, K.L.; Kim, E.; Miura, T.; Torii, K.U. MUTE directly orchestrates cell-state switch and the single symmetric division to create stomata. Dev. Cell 2018, 45, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, M.; Ueda, H.; Shimada, T.; Hara-Nishimura, I. FAMA: A molecular link between stomata and myrosin cells. Trends Plant Sci. 2016, 21, 861–871. [Google Scholar] [CrossRef]
- Han, S.; Torii, K.U. Lineage-specific stem cells, signals and asymmetries during stomatal development. Development 2016, 143, 1259–1270. [Google Scholar] [CrossRef] [Green Version]
- Lau, O.S.; Davies, K.A.; Chang, J.; Adrian, J.; Rowe, M.H.; Ballenger, C.E.; Bergmann, D.C. Direct roles of SPEECHLESS in the specification of stomatal self-renewing cells. Science 2014, 345, 1605–1609. [Google Scholar] [CrossRef] [Green Version]
- Hara, K.; Yokoo, T.; Kajita, R.; Onishi, T.; Yahata, S.; Peterson, K.; Torii, K.; Kakimoto, T. Epidermal cell density is autoregulated via a secretory peptide, EPIDERMAL PATTERNING FACTOR 2 in Arabidopsis leaves. Plant Cell Physiol. 2009, 50, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Horst, R.J.; Fujita, H.; Lee, J.S.; Rychel, A.L.; Garrick, J.M.; Kawaguchi, M.; Peterson, K.M.; Torii, K.U. Molecular framework of a regulatory circuit initiating two-dimensional spatial patterning of stomatal lineage. PLoS Genet. 2015, 11, e1005374. [Google Scholar] [CrossRef] [Green Version]
- Caine, R.S.; Chater, C.C.; Kamisugi, Y.; Cuming, A.C.; Beerling, D.J.; Gray, J.E.; Fleming, A.J. An ancestral stomatal patterning module revealed in the non-vascular land plant Physcomitrella patens. Development 2016, 143, 3306–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chater, C.C.; Caine, R.S.; Tomek, M.; Wallace, S.; Kamisugi, Y.; Cuming, A.C.; Lang, D.; MacAlister, C.A.; Casson, S.; Bergmann, D.C.; et al. Origin and function of stomata in the moss Physcomitrella patens. Nat. Plants 2016, 2, 16179. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Ohashi-Ito, K.; Bergmann, D.C. Orthologs of Arabidopsis thaliana stomatal bHLH genes and regulation of stomatal development in grasses. Development 2009, 136, 2265–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, C.; Caine, R.S.; Harrison, E.L.; Sloan, J.; Gray, J.E. Stomatal development: Focusing on the grasses. Curr. Opin. Plant Biol. 2018, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nunes, T.D.G.; Zhang, D.; Raissig, M.T. Form, development and function of grass stomata. Plant J. 2019, 101, 780–799. [Google Scholar] [CrossRef]
- Rudall, P.J.; Hilton, J.; Bateman, R.M. Several developmental and morphogenetic factors govern the evolution of stomatal patterning in land plants. New Phytol. 2013, 200, 598–614. [Google Scholar] [CrossRef]
- Raissig, M.T.; Abrash, E.; Bettadapur, A.; Vogel, J.P.; Bergmann, D.C. Grasses use an alternatively wired bHLH transcription factor network to establish stomatal identity. Proc. Natl. Acad. Sci. USA 2016, 113, 8326–8331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Chen, L.; Yu, Q.; Zhou, W.; Gou, X.; Li, J.; Hou, S. Multiple transcriptional factors control stomata development in rice. New Phytol. 2019, 223, 220–232. [Google Scholar] [CrossRef]
- Raissig, M.T.; Matos, J.L.; Anleu Gil, M.X.; Kornfeld, A.; Bettadapur, A.; Abrash, E.; Allison, H.R.; Badgley, G.; Vogel, J.P.; Berry, J.A.; et al. Mobile MUTE specifies subsidiary cells to build physiologically improved grass stomata. Science 2017, 355, 1215–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Biswal, A.K.; Dionora, J.; Perdigon, K.M.; Balahadia, C.P.; Mazumdar, S.; Chater, C.; Lin, H.; Coe, R.A.; Kretzschmar, T.; et al. CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep. 2017, 36, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Serna, L. The role of grass MUTE orthologues during stomatal development. Front. Plant Sci. 2020, 11, 55. [Google Scholar] [CrossRef]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Schulman, A.H.; Tanskanen, J.; Spannagl, M.; Twardziok, S.; Mascher, M.; Springer, N.M.; Li, Q.; Waugh, R.; Li, C.; et al. The repetitive landscape of the 5100 Mbp barley genome. Mob. DNA 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Swati, K.C.; Tiwari, J.P.; Kumar, A.; Goel, P. Genetic architecture of barley (Hordeum vulgare L.) genotypes for grain yield and yield attributing traits. Wheat Barley Res. 2018, 10, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef]
- The International Barley Genome Sequencing Consortium. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–716. [Google Scholar] [CrossRef]
- Fan, Y.; Zhou, G.; Shabala, S.; Chen, Z.; Cai, S.; Li, C.; Zhou, M. Genome-wide association study reveals a new QTL for salinity tolerance in barley (Hordeum vulgare L.). Front. Plant Sci. 2016, 7, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazzouri, K.M.; Khraiwesh, B.; Amiri, K.M.A.; Pauli, D.; Blake, T.; Shahid, M.; Mullath, S.K.; Nelson, D.; Mansour, A.L.; Salehi-Ashtiani, K.; et al. Mapping of HKT1;5 gene in barley using GWAS approach and its implication in salt tolerance mechanism. Front. Plant Sci. 2018, 9, 156. [Google Scholar] [CrossRef] [Green Version]
- Monat, C.; Padmarasu, S.; Lux, T.; Wicker, T.; Gundlach, H.; Himmelbach, A.; Ens, J.; Li, C.; Muehlbauer, G.J.; Schulman, A.H.; et al. TRITEX: Chromosome-scale sequence assembly of Triticeae genomes with open-source tools. Genome Biol. 2019, 20, 284. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sun, G.; Ren, X.; Du, B.; Cheng, Y.; Wang, Y.; Li, C.; Sun, D. Dissecting the genetic basis of grain size and weight in barley (Hordeum vulgare L.) by QTL and comparative genetic analyses. Front. Plant Sci. 2019, 10, 469. [Google Scholar] [CrossRef]
- Gous, P.W.; Hickey, L.; Christopher, J.T.; Franckowiak, J.; Fox, G.P. Discovery of QTL for stay-green and heat-stress in barley (Hordeum vulgare) grown under simulated abiotic stress conditions. Euphytica 2015, 207, 305–317. [Google Scholar] [CrossRef]
- Gudys, K.; Guzy-Wrobelska, J.; Janiak, A.; Dziurka, M.A.; Ostrowska, A.; Hura, K.; Jurczyk, B.; Żmuda, K.; Grzybkowska, D.; Śróbka, J.; et al. Prioritization of candidate genes in QTL regions for physiological and biochemical traits underlying drought response in barley (Hordeum vulgare L.). Front. Plant Sci. 2018, 9, 769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwando, E.; Han, Y.; Angessa, T.T.; Zhou, G.; Hill, C.B.; Zhang, X.; Li, C. Genome-wide association study of salinity tolerance during germination in Barley (Hordeum vulgare L.). Front. Plant Sci. 2020, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, P.; Yang, Z.; Xu, C. Genetic mapping of quantitative trait loci in crops. Crop J. 2017, 5, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Long, Z.; Jia, Y.; Tan, C.; Zhang, X.; Angessa, T.; Broughton, S.; Westcott, S.; Dai, F.; Zhang, G.; Sun, D.; et al. Genetic mapping and evolutionary analyses of the black grain trait in barley. Front. Plant Sci. 2019, 9, 1921. [Google Scholar] [CrossRef] [PubMed]
- Choo, T. Genetic resources of Tibetan barley in China. Crop Sci. 2002, 42, 1759–1760. [Google Scholar] [CrossRef]
- Han, Y.; Broughton, S.; Liu, L.; Zhang, X.; Zeng, J.; He, X.; Li, C. Highly efficient and genotype-independent barley gene editing based on anther culture. Plant Commun. 2020, 2, 100082. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Ni, S.; Zhang, G. Transcriptome and metabolome analysis reveals regulatory networks and key genes controlling barley malting quality in responses to drought stress. Plant Physiol. Biochem. 2020, 152, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pham, A.; Maurer, A.; Pillen, K.; Brien, C.; Dowling, K.; Berger, B.; Eglinton, J.K.; March, T.J. Genome-wide association of barley plant growth under drought stress using a nested association mapping population. BMC Plant Biol. 2019, 19, 134. [Google Scholar] [CrossRef] [Green Version]
- Wehner, G.G.; Balko, C.C.; Enders, M.M.; Humbeck, K.K.; Ordon, F.F. Identification of genomic regions involved in tolerance to drought stress and drought stress induced leaf senescence in juvenile barley. BMC Plant Biol. 2015, 15, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, D.; Sánchez-Bermejo, E.; de Marcos, A.; Martín-Jimenez, C.; Fenoll, C.; Alonso-Blanco, C.; Mena, M. A genetic dissection of natural variation for stomatal abundance traits in Arabidopsis. Front. Plant Sci. 2019, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, Y.; Zhao, P.; Xu, P.; Yu, G.; Zhang, L.; Xiong, Y.; Xiang, C. AtEDT1/HDG11 regulates stomatal density and water-use efficiency via ERECTA and E2Fa. New Phytol. 2019, 223, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dai, X.; Xu, S.; Zhao, P.X. GPU empowered pipelines for calculating genome-wide kinship matrices with ultra-high dimensional genetic variants and facilitating 1D and 2D GWAS. NAR Genom. Bioinform. 2019, 2, lqz009. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Yan, J. Crop genome-wide association study: A harvest of biological relevance. Plant J. 2018, 97, 8–18. [Google Scholar] [CrossRef]
- Jabbari, M.; Fakheri, B.A.; Aghnoum, R.; Nezhad, N.M.; Ataei, R. GWAS analysis in spring barley (Hordeum vulgare L.) for morphological traits exposed to drought. PLoS ONE 2018, 13, e0204952. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Sun, G.; Ren, X.; Li, C.; Sun, D. Identification of QTL underlying physiological and morphological traits of flag leaf in barley. BMC Genet. 2015, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- Janiak, A.; Kwasniewski, M.; Sowa, M.; Gajek, K.; Żmuda, K.; Kościelniak, J.; Szarejko, I. No time to waste: Transcriptome study reveals that drought tolerance in barley may be attributed to stressed-like expression patterns that exist before the occurrence of stress. Front. Plant Sci. 2018, 8, 2212. [Google Scholar] [CrossRef]
- Adrian, J.; Chang, J.; Ballenger, C.E.; Bargmann, B.O.R.; Alassimone, J.; Davies, K.A.; Lau, O.S.; Matos, J.L.; Hachez, C.; Lanctot, A.; et al. Transcriptome dynamics of the stomatal lineage: Birth, amplification, and termination of a self-renewing population. Dev. Cell 2015, 33, 107–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmielewska, K.; Rodziewicz, P.; Swarcewicz, B.; Sawikowska, A.; Krajewski, P.; Marczak, Ł.; Ciesiołka, D.; Kuczyńska, A.; Mikołajczak, K.; Ogrodowicz, P.; et al. Analysis of drought-induced proteomic and metabolomic changes in barley (Hordeum vulgare L.) leaves and roots unravels some aspects of biochemical mechanisms Involved in drought tolerance. Front. Plant Sci. 2016, 7, 1108. [Google Scholar] [CrossRef] [PubMed]
- Gallusci, P.; Dai, Z.; Génard, M.; Gauffretau, A.; Leblanc-Fournier, N.; Richard-Molard, C.; Vile, D.; Brunel-Muguet, S. Epigenetics for plant improvement: Current knowledge and modeling avenues. Trends Plant Sci. 2017, 22, 610–623. [Google Scholar] [CrossRef]
- Robertsen, C.D.; Hjortshøj, R.L.; Janss, L.L. Genomic selection in cereal breeding. Agronomy 2019, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, N.; Douches, D.; Dhingra, A.; Glenn, K.C.; Herzig, P.R.; Stowe, E.C.; Swarup, S. The role of conventional plant breeding in ensuring safe levels of naturally occurring toxins in food crops. Trends Food Sci. Technol. 2020, 100, 51–66. [Google Scholar] [CrossRef]
- National Research Council (US) Committee on Identifying and Assessing Unintended Effects of Genetically Engineered Foods on Human Health. Safety of Genetically Engineered Foods: Approaches to Assessing Unintended Health Effects; National Academies Press: Washington, DC, USA, 2004; p. 3, Unintended Effects from Breeding. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK215778/ (accessed on 25 June 2021).
- Yu, Q.; Chen, L.; Zhou, W.; An, Y.; Luo, T.; Wu, Z.; Wang, Y.; Xi, Y.; Yan, L.; Hou, S. RSD1 is essential for stomatal patterning and files in rice. Front. Plant Sci. 2020, 11, 600021. [Google Scholar] [CrossRef]
- Kamiya, N.; Itoh, J.; Morikami, A.; Nagato, Y.; Matsuoka, M. The SCARECROW gene’s role in asymmetric cell divisions in rice plants. Plant J. 2003, 36, 45–54. [Google Scholar] [CrossRef]
- Wang, H.; Guo, S.; Qiao, X.; Guo, J.; Li, Z.; Zhou, Y.; Bai, S.; Gao, Z.; Wang, D.; Wang, P.; et al. BZU2/ZmMUTE controls symmetrical division of guard mother cell and specifies neighbor cell fate in maize. PLoS Genet. 2019, 15, e1008377. [Google Scholar] [CrossRef]
- Abrash, E.; Anleu Gil, M.X.A.; Matos, J.L.; Bergmann, D.C. Conservation and divergence of YODA MAPKKK function in regulation of grass epidermal patterning. Development 2018, 145, dev165860. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Species | Gene Name | Function | Citation |
|---|---|---|---|
| Rice | RSD1 (RICE STOMATAL DEVELOPMENT DEFECT 1) | Inhibits ectopic asymmetric division and clustering, stomatal file specification, regulator of stomatal development, required for normal expression of related genes in the stomatal development pathway. | [144] |
| Rice, Maize | SDD1 (STOMATAL DENSITY AND DISTRIBUTION 1) | Inhibits ectopic asymmetric division and clustering, negative regulator of stomatal density. | [69,144] |
| Rice | SCR1/2 (SCARECROW 1/2) | Controls cell entry into the stomatal lineage (meristemoid mother cells to meristemoids), regulates subsidiary cell formation, expression regulated by OsSPCH and OsMUTE (Rice). | [108,145] |
| Rice, Maize | SHR1/2 (SHORTROOT 1/2) | Stomatal file positioning, controls cell entry into the stomatal lineage, regulates subsidiary cell formation, regulator of stomatal density. | [66] |
| Rice, Maize, Bradypodium | MUTE | Guard mother cell formation, migrates to subsidiary mother cells to induce subsidiary cell formation, regulates expression of multiple genes in the stomatal development pathway. | [108,109,146] |
| Rice, Bradypodium | SPCH1/2 (SPEECHLESS) | Controls cell entry into the stomatal lineage, stomatal file development, SPCH2 knockout mutants exhibit a greater decrease in stomatal density versus SPCH1 knockout mutants. | [103,107,108] |
| Rice | FLP (FOUR LIPS) | Negative regulator of guard mother cell symmetric division, maintains orientation of symmetric division in guard mother cells. | [108] |
| Rice | FAMA | Regulation of subsidiary mother cell asymmetric division, ensures correct formation or mature guard cell complexes. | [107,108] |
| Rice, Bradypodium | ICE1/2 (INDUCER OF CBF EXPRESSION 1/2) aka SCREAM1/2 | Heterodimeric partner of SPCH, MUTE and FAMA, regulation of cell entry into the stomatal lineage, guard mother cell formation, guard cell maturation. | [107,108] |
| Rice, Wheat, Barley | EPF1/2 (EPIDERMAL PATTERNING FACTOR1/2) | Negative regulator of stomatal development, inhibits SPCH activity. | [48,58,62] |
| Bradypodium | YODA (YDA) | Negative regulator of stomatal development, regulates asymmetric cell divisions, inhibits stomatal clustering, maintains normal stomatal spacing/patterning, inhibits SPCH activity. | [147] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, B.C.; He, T.; Li, C. The Genetic Control of Stomatal Development in Barley: New Solutions for Enhanced Water-Use Efficiency in Drought-Prone Environments. Agronomy 2021, 11, 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081670
Robertson BC, He T, Li C. The Genetic Control of Stomatal Development in Barley: New Solutions for Enhanced Water-Use Efficiency in Drought-Prone Environments. Agronomy. 2021; 11(8):1670. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081670
Chicago/Turabian StyleRobertson, Brittany Clare, Tianhua He, and Chengdao Li. 2021. "The Genetic Control of Stomatal Development in Barley: New Solutions for Enhanced Water-Use Efficiency in Drought-Prone Environments" Agronomy 11, no. 8: 1670. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081670