
VaCRK2 Mediates Gray Mold Resistance in Vitis amurensis by Activating the Jasmonate Signaling Pathway


1
Shandong Academy of Grape, Shandong Academy of Agricultural Sciences, No. 1–27, Shanda South Road, Jinan 250100, China
2
Institute of Agro-Food Sciences and Technology, Shandong Academy of Agricultural Sciences, No. 202, Gongye North Road, Jinan 250100, China
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(8), 1672; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081672
Submission received: 29 June 2021
/
Revised: 19 August 2021
/
Accepted: 19 August 2021
/
Published: 23 August 2021
(This article belongs to the Special Issue Advances in Pesticide Discovery and Application)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Cysteine-rich receptor-like kinases (CRKs) are ubiquitous plant receptor-like kinases, which play a significant role in plant disease resistance. Gray mold is an economically important disease of grapes caused by Botrytis cinerea. However, CRK genes and their function in gray mold disease resistance in grapes have not been elucidated. This study aimed to identify and characterize CRKs in grapes and determine their role in gray mold resistance. Four CRKs were identified in Vitis amurensis and named VaCRK1–VaCRK4 according to their genomic distribution. The four VaCRKs were ectopically expressed in Arabidopsis thaliana to study their function in defense response against B. cinerea. Heterologous expression of VaCRK2 in A. thaliana conferred resistance to B. cinerea. VaCRK2 expression in gray mold-resistant grape cultivar increased significantly after B. cinerea inoculation and methyl jasmonate treatment. Furthermore, the expression of jasmonic acid (JA) signaling pathway-related genes in VaCRK2 overexpression lines of A. thaliana was significantly increased after B. cinerea inoculation, leading to the upregulation of pathogenesis-related (PR) genes and reactive oxygen species (ROS) accumulation. Overall, these results suggest that VaCRK2 confers resistance to B. cinerea by activating PR gene expression and oxidative burst through the JA signaling pathway.
1. Introduction
Two layers of immune system response have evolved during the arms race between plants and phytopathogens, including bacteria, fungi and viruses [1]. One involves the recognition of specific molecular motifs conserved within a class of pathogens, also known as pathogen-associated molecular patterns (PAMP), through receptors located on the plant cell membrane, thereby activating the PAMP-triggered immunity (PTI) to resist pathogen invasion [2]. However, sometimes pathogens manage to bypass PTI by secreting effectors into the plant cell. In response, plants evolved another form of immune response involving resistance (R) proteins. These proteins perceive specific virulence effectors secreted by pathogens, thereby triggering the effector-triggered immunity (ETI), a more robust immune response against pathogen attack [3]. Recent studies have found that ETI can increase the expression of core PTI components, thereby amplifying the PTI and inducing a durable immune response [4]. For example, ETI enhances the expression of respiratory burst oxidase homolog D (RBOHD) protein, while PTI promotes the complete activation of RBOHD protein. This cooperation mechanism ensures that plants mount a rapid and precise response against pathogen infection while maintaining optimal growth and development [4].
PTI response is activated when receptors on the plant cell membrane interact with specific pathogen molecular patterns. Plants use a variety of plasma membrane-bound receptor proteins to sense exogenous immune signals [5]. These cell surface receptor proteins have different extracellular domains for sensing specific ligands. Receptor-like kinases (RLKs) are an important type of receptor proteins mainly composed of an N-terminal signal peptide, a variable extracellular domain, a transmembrane domain and a C-terminal intracellular kinase domain [6]. They can be divided into different subfamilies based on the nature of their variable extracellular domain, including leucine-rich repeats (LRRs), lectins (Lec), lysine motifs (LysMs) and cell wall-related kinases (WAKs) [7]. Generally, the N-terminus of RLKs, including signal peptide and variable extracellular domain, extends to the apoplast and can sense the signals released by pathogens. Meanwhile, the C-terminal kinase domain is located in the cytoplasm and transmits signals into the intracellular space to activate the plant cell immune response [8].
Cysteine-rich receptor-like kinases (CRKs) belong to an important class of RLKs that plays a vital function in plant disease resistance [9]. The extracellular domain of CRKs comprises two DUF26 motifs and a conserved C-X8-C-X2-C motif. The conserved Cys residues of the C-X8-C-X2-C motif facilitate the formation of the three-dimensional structure of proteins through disulfide bonds or zinc finger motifs, like in many DNA binding transcription factors [6]. Notably, disulfide bond and zinc finger can mediate protein–protein interaction, which is a key step in receptor activation [6]. The role of CRKs in plant defense response has been revealed in many plants [10,11,12,13,14]. In Arabidopsis thaliana, overexpression of CRK13 and CRK28 improves resistance to Pseudomonas syringae [10,11]. In wheat, TaCRK2 enhances resistance to Puccinia triticina and has a positive effect on pathogen-induced plant cell death [12]. Rice CRK10 mediates the resistance of rice to Xanthomonas oryzae pv. oryzae by interacting with transcription factor TGA2.1 [13]. In addition, HvCRK1 negatively regulates the basal resistance of barley to Blumeria graminis f. sp. hordei [14].
Gray mold disease is mainly caused by Botrytis cinerea, a common fungal pathogen infecting more than 1400 plant species, including tomato, strawberry, grape and pepper [15]. Gray mold is the primary disease during the growth, storage and transportation of grapes [15]. It causes 20–50% yield losses in grapes if not controlled [16]. Improving host disease resistance is the most economical and effective strategy for controlling B. cinerea. However, the process requires identifying resistance genes, which can then be incorporated into the grape breeding programs. At present, a few genes that can confer resistance or tolerance to B. cinerea have been identified in grape, including VvNPR1.1 [16], VaSTS19 [17], VlWRKY3 [18], VqSTS21 [19] and VqAP13 [20]. The introduction of VaSTS19 and VvNPR1.1 genes into A. thaliana improved the resistance to B. cinerea [16,17]. In contrast, overexpression of VlWRKY3, VqSTS21, and VqAP13 genes in A. thaliana increased susceptibility to B. cinerea [18,19,20]. The demand for gray mold resistance genes in grape breeding has continued to increase with the continuous evolution of B. cinerea. Thus, there is a need to mine new resistance genes against the pathogen.
Vitis amurensis is a grape species that exhibits excellent resistance to diverse diseases, including gray mold. It is, therefore, an important source of disease resistance genes [21]. To the best of our knowledge, there is no report on the disease resistance function of CRK genes in grapes. This study (1) identified CRK genes in the V. amurensis genome and validated their role in B. cinerea resistance in A. thaliana, (2) analyzed the expression patterns of the candidate gene in response to B. cinerea infection and hormone induction, (3) evaluated the relationship between hormones and plant defense response mediated by candidate gene and (4) identified the signaling pathway of candidate gene-mediated resistance to B. cinerea. This study illuminates the role of the grape CRK gene in resistance against B. cinerea and provides a reference for molecular breeding of grapes for resistance against gray mold disease.
2. Materials and Methods
2.1. Identification of the VaCRKs
Genomic data of V. amurensis were download from http://www.grapeworld.cn/am/download/ (accessed on 11 December 2020). HMMER program was used to match Stress-antifung protein and Pkinase protein based on the HMM profile of Stress-antifung (PF01657) domain and Pkinase (PF00069) domain in V. amurensis genome with an E-value cutoff of e−5 [22]. SignalP-5.0 (http://www.grapeworld.cn/am/download/ (accessed on 15 December 2020)) and TMHMM Server v. 2.0 (http://www.cbs.dtu.dk/services/TMHMM/ (accessed on 15 December 2020)) were used to verify the presence of signal peptides and transmembrane domain, respectively [23,24]. Putative genes were further verified based on the SMART website and InterproScan database [25,26].
2.2. Analysis of Subcellular Localization of VaCRK2
The primers of VaCRK2 gene were designed according to the full-length VAG0104573.1 sequence. The full-length VaCRK2 gene was amplified by PCR using cDNA of V. amurensis cultivar (cv. Shuangyou) as template. Purified PCR products were then inserted into pBin-GFP4 vector, fused with GFP gene, under the control of cauliflower mosaic virus 35S promoter [6]. The vectors p35S:GFP and p35S:VaCRK2 were transiently expressed in tobacco leaf epidermal cells using the Agrobacterium tumefaciens infiltration method [12]. Laser scanning confocal microscope (LSMT-PMT) was used to examine the subcellular localization of the proteins at 488 and 510 nm excitation and emission wavelengths, respectively.
2.3. Grape Plant Materials and Treatments
Grape species V. amurensis cv. Shuangyou and V. vinifera cv. Red Globe were used for the experiments. Notably, cv. Shuangyou is resistant to B. cinerea, whereas cv. Red Globe is susceptible. B. cinerea strain BcSD3 (highly virulent strain) was cultured in potato dextrose agar (PDA) medium to produce conidia. Conidia were eluted with sterile deionized water and adjusted to a concentration of 2 × 106 conidia/mL. Grape leaves were inoculated with BcSD3 and harvested at 0, 2, 6, 12, 24, 48, 72 and 120 h time points to assess resistance to B. cinerea. Grape leaves were sprayed with 10 mM methyl jasmonate (MeJA), 10 mM salicylic acid (SA), 10 mM ethephon (ETH) and 100 mM abscisic acid (ABA) to explore related signaling pathways. Leaves of control plants were sprayed with sterile distilled water. The leaves were collected at 0, 2, 6, 12, 24, 48 and 72 h time points. Six leaves per replicate, repeated three times, were harvested at each time point, frozen in liquid nitrogen and stored at −80 °C for subsequent analysis.
2.4. Generation and Analysis of Transgenic A. thaliana
The ORF sequence of VaCRK2 gene was integrated into vector pBI121 under the control of cauliflower mosaic virus 35S promoter. The vector was then transformed into Agrobacterium tumefaciens strain GV3101 using freeze–thaw method [18]. VaCRK2 gene was introduced into A. thaliana wildtype Col-0 using A. tumefaciens mediated transformation [27]. Putatively transformed events were analyzed using PCR and RT-PCR to identify T3 homozygous transgenic plants. Genomic DNA and cDNA from A. thaliana wildtype Col-0 were used as the control for PCR and RT-PCR detection [19].
2.5. Evaluation of Transgenic A. thaliana for Resistance against B. cinerea
A. thaliana wildtype Col-0 and VaCRK2 overexpression lines (OE1, OE2 and OE3) were used for the experiments. BcSD3 strain was cultured in PDA medium for five days. Conidia were eluted with sterile deionized water and adjusted to a concentration of 2 × 106 conidia/mL. Three-week-old A. thaliana plants were inoculated with B. cinerea conidia suspension. After inoculation, the plants were incubated at 22 °C under 16 h photoperiod with a humidity of 80–100%. A total of 24 leaves of six plants (four leaves per plant) were inoculated in one experiment, and the experiment was repeated thrice. The lesion diameter and fungal biomass were determined three days post-inoculation. Six leaves per replicate were harvested at each time point. The samples were frozen in liquid nitrogen and stored at −80 °C for subsequent analysis.
2.6. Oxidative Burst Assay in A. thaliana
A. thaliana wildtype Col-0 and VaCRK2 overexpression 1 line (OE1) were used for the experiments. Reactive oxygen species accumulation was determined by diaminobenzidine (DAB) staining. Three-week-old A. thaliana leaves were infiltrated with 10 μL conidia suspension of B. cinerea. The A. thaliana leaves were infiltrated with sterile water as a control. After 12 h, the leaves were immersed in DAB buffer and vacuum infiltrated at 25 °C in the dark. The reaction was terminated after 12 h, and the leaves were washed with sterile water, bleached using 75% ethanol, and then immersed in 30% glycerin. ImageJ software was used to determine the percentage of brown pixels in the leaves. Antioxidant enzymes, including peroxidase (POD) and catalase (CAT), were extracted from A. thaliana leaves and measured as described by Guo et al. [18].
2.7. Relative Gene Expression Analysis
PLANTeasy kit (YPH-Bio, Beijing, China) was used to extract total RNA. After extraction and quality verification, the RNA samples were reverse transcribed to cDNA using TIANSeq M-MLV (RNase H-) reverse transcriptase (Tiangen, Beijing, China) according to the manufacturer’s instructions. Then, RT-qPCR analysis was performed using the SYBR Premix Ex Taq kit (TaKaRa, Kusatsu, Japan) on the QuantStudio 6 Flex qPCR System (Applied Biosystems, Foster City, CA, USA). The 2−ΔΔCT method was used to determine relative gene expression [28]. V. vinifera EF1γ (CB977561), A. thaliana UBQ5 (NM_116090.3) and APT1 (NM_102509.4) were used as the internal reference genes. The primer sequences are listed in Table S1.
3. Results
3.1. VaCRK2 Enhances Resistance to B. cinerea in A. thaliana
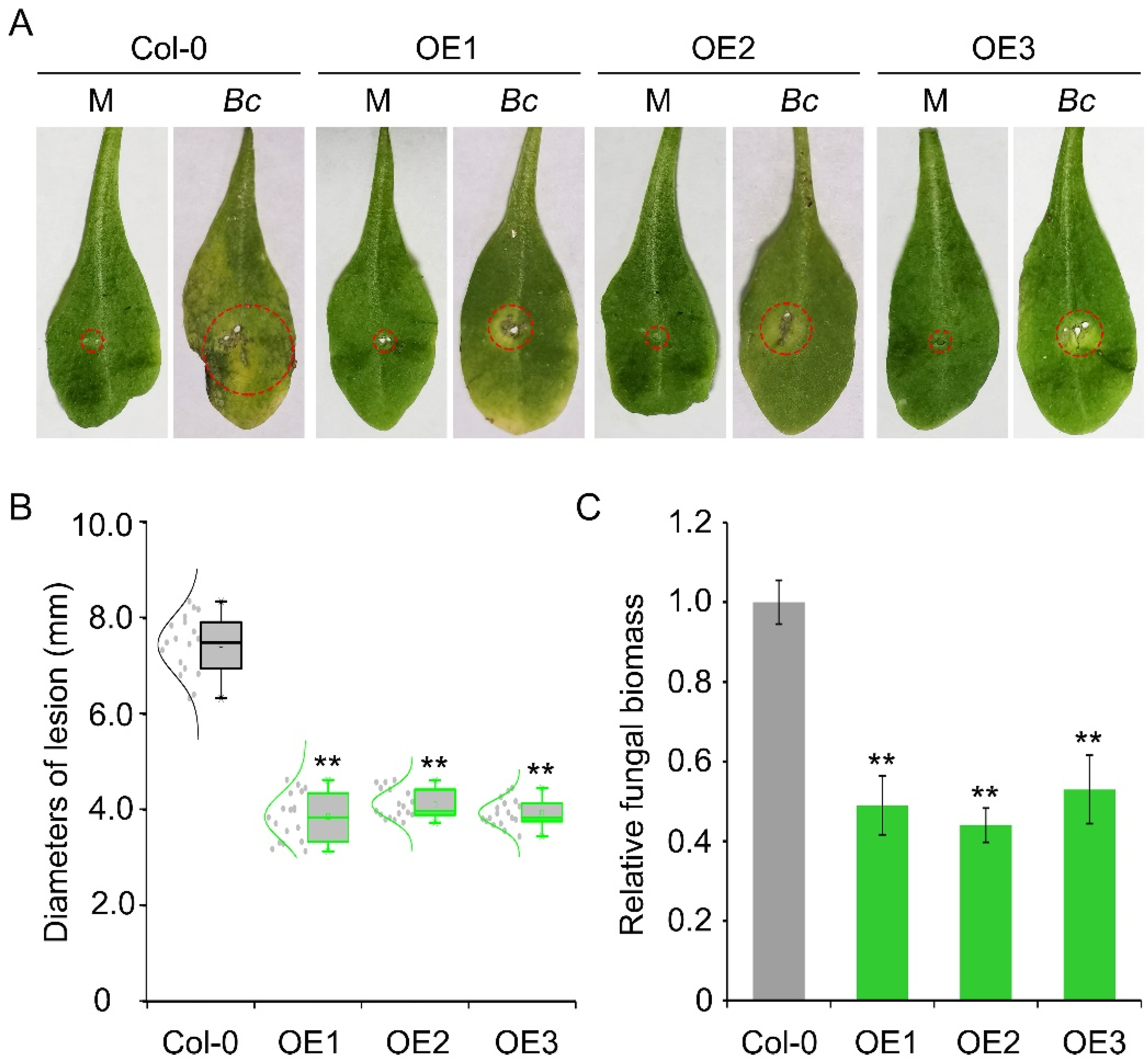
Four cysteine-rich receptor-like kinase (CRK) genes were identified in V. amurensis genome and named VaCRK1–VaCRK4 according to their position in the genome. The protein encoded by VaCRK genes comprises 200 to 420 amino acids in length. The predicted molecular weight is 48 to 66 KDa, and the isoelectric point ranges from 3.7 to 5.2 (Table S2). The four VaCRK genes were transformed into A. thaliana wildtype Col-0, and at least three overexpression lines were obtained following PCR and RT-PCR analyses. The disease assay results showed that the lesion area and fungal biomass of VaCRK2 overexpression lines were 56% and 40% lower than those of the wildtype Col-0, respectively, indicating that the VaCRK2 overexpression lines were more resistant to B. cinerea than the wildtype Col-0 (Figure 1). Notably, transgenic A. thaliana lines overexpressing the other three VaCRK genes VaCRK1, VaCRK3 and VaCRK4 showed no significant improvement in resistance to B. cinerea compared with the wildtype Col-0 (Figure S1).
3.2. VaCRK2 Localizes to Plasma Membrane
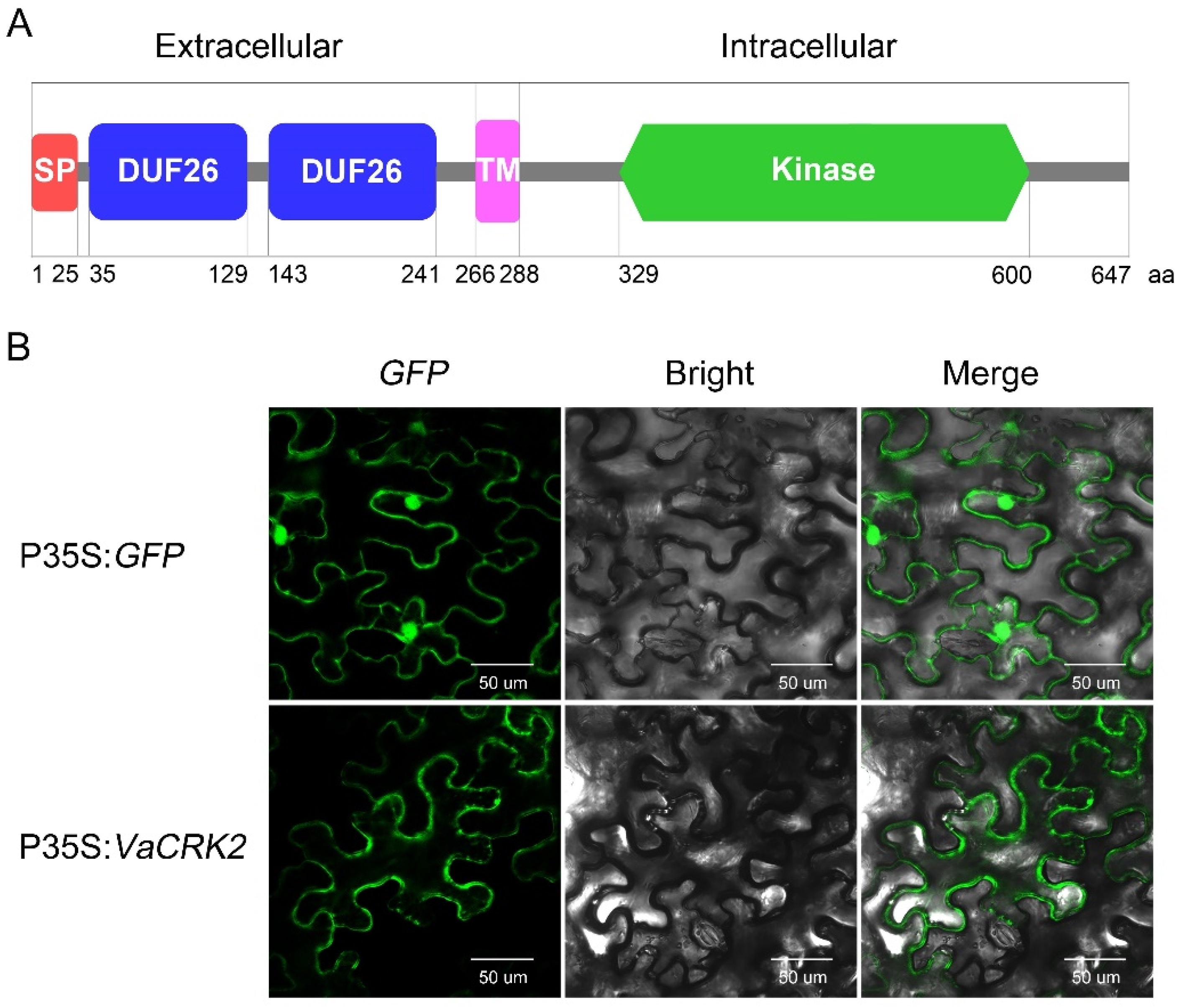
Conserved domain analysis of VaCRK2 peptide sequence using SMART identified a typical CRK family protein with signal peptide and TM domain, which were confirmed by SignalP5.0 (1–25 aa) and TMHMM2.0 (266–288 aa), respectively (Figure 2A). To further understand the function of VaCRK2, the subcellular localization was verified by transient expression of VaCRK2 and green fluorescent fusion protein (VaCRK2-GFP) in tobacco leaf epidermal cells. According to the results, the fluorescent signal of VaCRK2-GFP fusion protein was clearly localized to the cell membrane, in contrast to the fluorescence signal of GFP proteins, which was prevalent throughout the foliar cells in tobacco, indicating that VaCRK2 was localized on the cell membrane (Figure 2B).
3.3. VaCRK2 Positively Responds to B. cinerea and MeJA Treatment
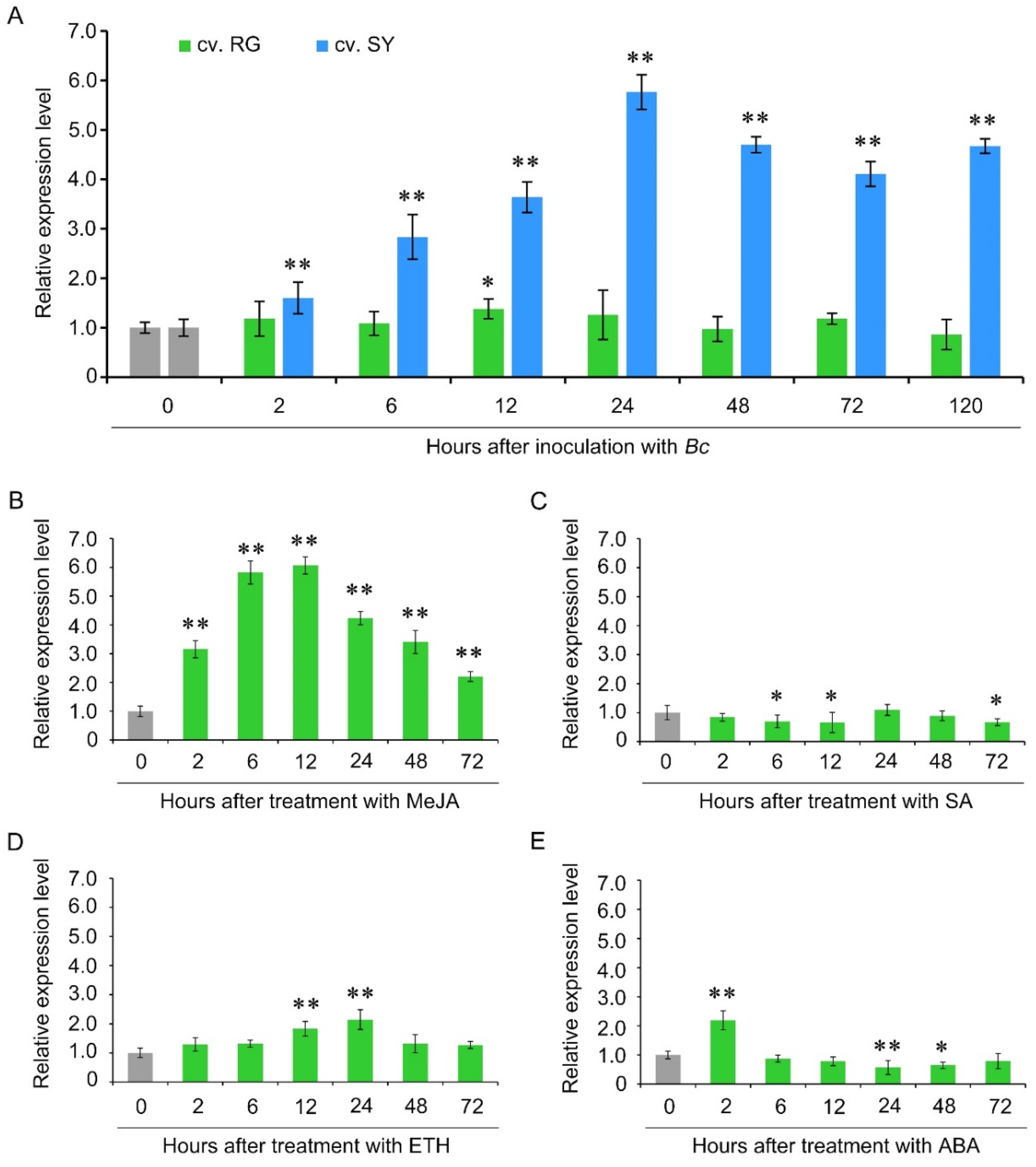
To determine the relationship between VaCRK2 expression and B. cinerea resistance, we compared the expression patterns of VaCRK2 between the resistant and susceptible grape cultivars following B. cinerea inoculation. VaCRK2 expression was significantly upregulated in resistant cv. Shuangyou from 2 to 24 h post-inoculation, after which the expression was reduced (Figure 3A). In contrast, no significant change in VaCRK2 expression was observed in susceptible cv. Red Globe during the entire experimental period, indicating that VaCRK2 may mediate B. cinerea resistance in grapes. To explore VaCRK2-related signaling pathways, we analyzed the expression patterns of VaCRK2 after MeJA, SA, ETH and ABA treatments. VaCRK2 was significantly upregulated in resistant cv. Shuangyou after treatment with methyl MeJA within 120 h (Figure 3B) and less affected by treatment with ET (Figure 3D), but it was downregulated by treatment with SA or ABA at several points (Figure 3C,E), which suggested that the VaCRK2 response of the resistant cultivar to B. cinerea is mediated by JA signaling pathway. Overall, these findings show that B. cinerea infection and exogenous MeJA induce VaCRK2 expression in gray mold-resistant cv. Shuangyou.
3.4. VaCRK2 Mediates B. cinerea Resistance Via the JA Signaling Pathway
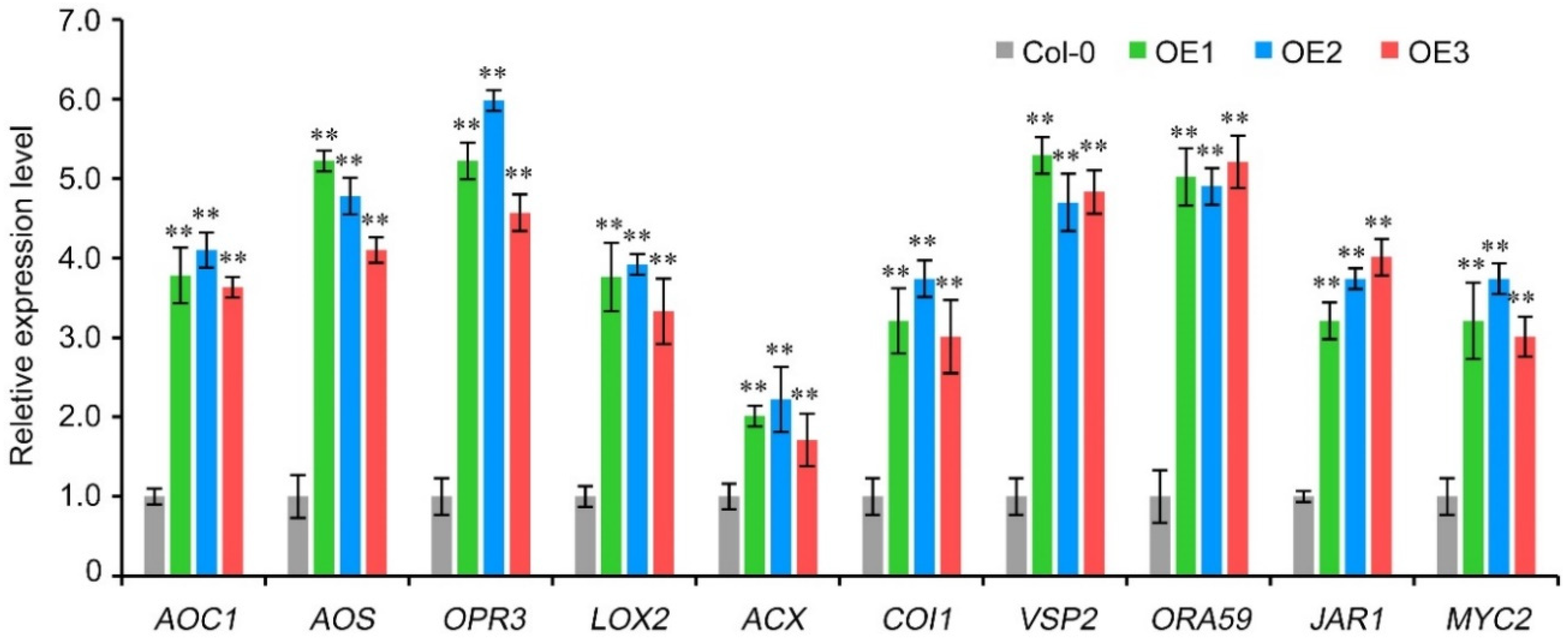
Jasmonic acid (JA) signaling pathway plays a vital role in plant disease resistance, especially against saprophytic fungal diseases [29]. Therefore, in order to verify whether VaCRK2 mediates resistance to B. cinerea through JA signaling pathway, we firstly detected the expression pattern of AtCRK2, a homolog of VaCRK2 in A. thaliana, and the results showed that the expression of AtCRK2 gene was not induced in A. thaliana wildtype Col-0 inoculated with B. cinerea (Figure S2). Secondly, we overexpressed VaCRK2 gene in JA-deficient A. thaliana mutant jar1 (SALK_030821); compared with the mutant jar1, the overexpressed line could not improve the resistance to B. cinerea (Figure S3). Thirdly, we examined the expression patterns of genes related to JA signaling pathway in A. thaliana wildtype Col-0 and VaCRK2 overexpression lines after inoculation with B. cinerea; the results showed that 11 JA signaling pathway-related genes, including AOC1, AOS, OPR3, LOX2, ACX, COI1, VSP2, ORA59, JAR1 and MYC2, were upregulated in both wildtype Col-0 and VaCRK2 overexpression lines but significantly more upregulated in VaCRK2 overexpression lines than in the wildtype Col-0 (Figure 4, Figures S4 and S5). Altogether, these results indicate that VaCRK2 interacts with the JA signaling pathway to mediate B. cinerea resistance in A. thaliana.
3.5. VaCRK2 Overexpression Affects PR Gene Expression and ROS Accumulation
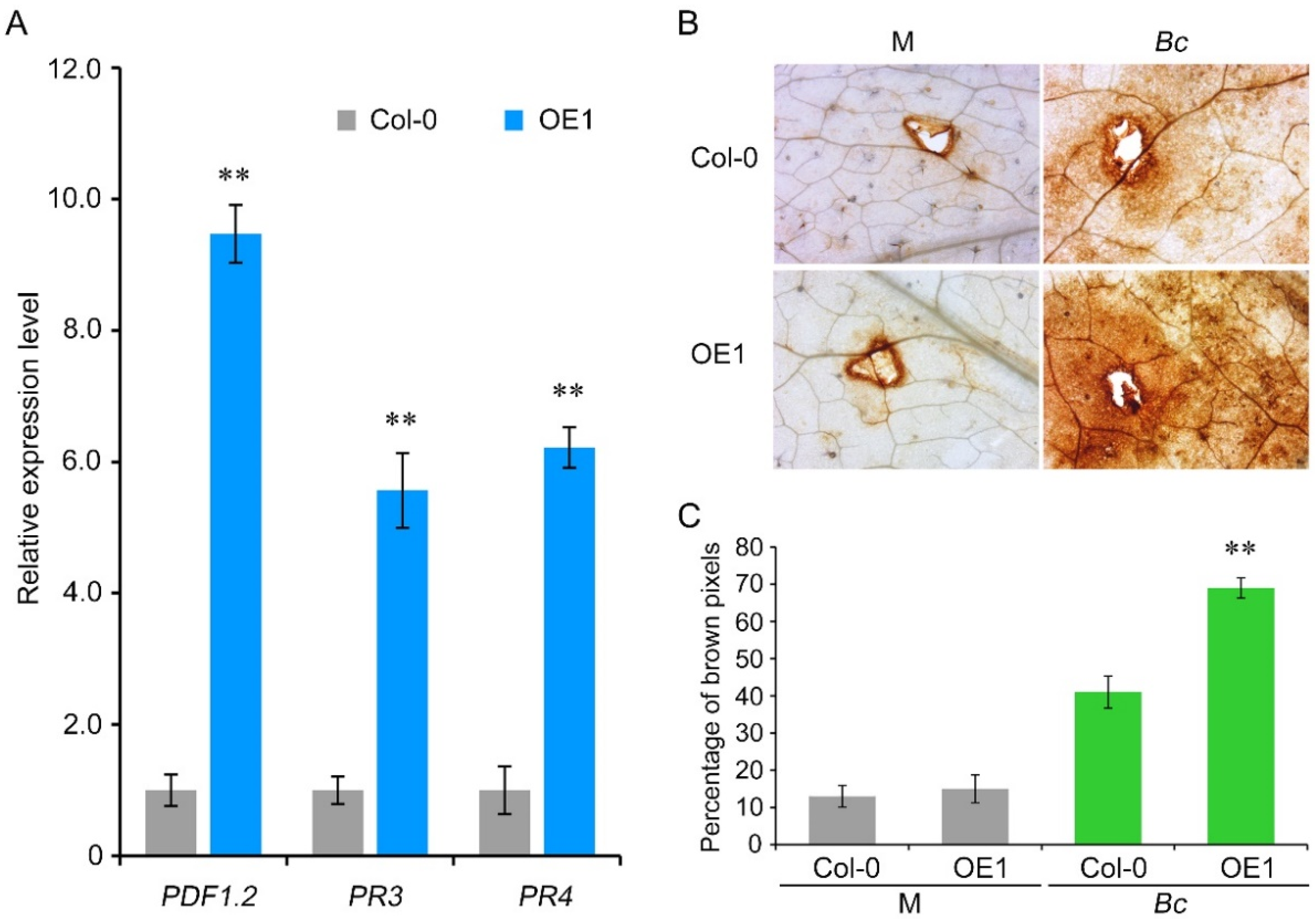
Pathogenesis-related (PR) gene expression and ROS accumulation in A. thaliana wildtype Col-0 and VaCRK2 overexpression lines were evaluated after B. cinerea inoculation to further verify that VaCRK2 facilities defense response against B. cinerea. The expression levels of JA signaling pathway defense-related genes (PDF1.2, PR3 and PR4) were significantly higher in the VaCRK2 overexpression lines than in the wildtype Col-0 after inoculation with B. cinerea (Figure 5A, Figures S6 and S7). Furthermore, ROS accumulation in the leaves of VaCRK2 overexpression lines was significantly higher than in wildtype Col-0 relative to sterile water treatment (Figure 5B,C). Peroxidase and catalase activity detection results showed that there was no significant difference in peroxidase and catalase activities between wildtype Col-0 and overexpression line of A. thaliana under control conditions. After inoculation with B. cinerea, the peroxidase and catalase activities in wildtype Col-0 and overexpression line of A. thaliana were increased, and the overexpression line showed significantly higher changes than wildtype Col-0 (Figure S8). Altogether, these results suggest that VaCRK2 mediates resistance to B. cinerea by upregulating PR genes and inducing oxidative burst in A. thaliana.
4. Discussion
Cysteine-rich receptor-like kinases (CRKs) have been shown to play an essential role in mediating disease resistance in several plants, including Arabidopsis [10,11,30], rice [13], barley [14], wheat [31] and Medicago truncatula [32]. However, CRK genes and their function in disease resistance have not been demonstrated in grapes. This study characterized CRK genes in V. amurensis (VaCRK) and evaluated their role in mediating B. cinerea resistance. The VaCRK genes identified in this study have a signal peptide and two DUF26 motifs at the N-terminus, a TM domain in the middle and a kinase domain at the C-terminus, which are typical domains of a CRK gene. Based on the CRK gene identification criteria, five candidate genes were identified in V. amurensis. However, VAG0113336.1 and VAG0128137.1 genes were the same and were combined into one gene, VaCRK3 (Table S2). Among the four CRK genes identified in V. amurensis, VaCRK2 was the only one shown to confer resistance to B. cinerea effectively. VaCRK2 is located on the cell membrane and significantly increased the resistance of A. thaliana overexpression lines to B. cinerea (Figure 1 and Figure 2). Studies have shown that ETI promotes PTI by inducing the expression of PTI signal components, while PTI ensures the normal resistance function of ETI through diverse signals [33]. Enhanced expression of immune system-related genes, especially PTI genes, is crucial in mediating plant disease resistance [34]. VaCRK2 expression was significantly increased in resistant cv. Shuangyou at several time points after B. cinerea inoculation, but not in susceptible cv. Red Globe (Figure 3A), suggesting that VaCRK2 potentially mediates B. cinerea resistance and its expression is cultivar-dependent. At the same time, it is speculated that this result may be because the host increased the transcription level of VaCRK2 gene through immune regulation after sensing the signal of B. cinerea, especially in the early stage, to resist the infection of B. cinerea [16,17]. Perhaps because of this, different grape varieties have resistant and susceptible phenotypes to pathogens, which is consistent with the disease resistance mediated by CRKs in other plants [6].
JA is an essential signaling molecule in plants and facilitates various physiological reactions, including seed germination, damage response and biological stress response [29]. In this study, exogenous MeJA application induced the expression of VaCRK2 gene (Figure 3B), indicating that B. cinerea resistance mediated by VaCRK2 may be related to the JA signaling pathway. VaCRK2 gene expression was upregulated at 12 and 24 h after ETH treatment and downregulated at 6, 12 and 72 h after SA treatment (Figure 3C,D), which may be because ETH and JA act synergistically, and SA has negative feedback regulation on JA [29]. The JA signaling pathway activates diverse defense-related proteins and secondary metabolites in response to necrotrophic fungal invasion [29]. B. cinerea, the causative agent of gray mold disease, is a typical necrotrophic fungus infecting several plant species. Several genes related to JA signaling pathway have been cloned from different plants. These include JA self-synthesis genes, such as LOX2, AOS and OPR3, and defense-related genes, such as PDF1.2, PR3 and PR4 [35]. In this study, several genes related to JA signaling pathway were upregulated in A. thaliana VaCRK2 overexpression lines compared with A. thaliana wildtype Col-0 after B. cinerea inoculation (Figure 4, Figures S2 and S8). The JA signaling pathway has been associated with gray mold resistance in different plants, including A. thaliana, cucumber and tomato [36,37,38]. For example, A. thaliana phyB and SDG8 genes participate in the resistance defense against B. cinerea by regulating JA biosynthesis [39,40]. In addition, SlMYB75 plays an active role in the resistance of tomato to B. cinerea by regulating the JA signaling pathway [38]. Therefore, we speculated that the JA signaling pathway may be essential in VaCRK2-mediated resistance against B. cinerea.
PR gene expression is upregulated in many plants after inoculation with phytopathogens, including fungi, bacteria or viruses, to attack pathogens and inhibit pathogen infection [41]. Oxidative burst plays a vital role in plant disease resistance by directly neutralizing pathogens and amplifying signals of other immune events [4]. Notably, ROS accumulation can directly kill the invading pathogens and activate cell wall cross-linking to restrict further pathogen invasion [42]. Plant resistance proteins, including CRK proteins, recognize specific pathogenic effectors and initiate various defense responses, such as PR gene overexpression and ROS accumulation [43,44]. For instance, overexpression of CRK5, CRK6, CRK36 and CRK45 in A. thaliana significantly enhances the resistance to Pseudomonas syringae pv. tomato DC3000 by upregulating PR genes and enhancing ROS accumulation [9,30,45]. CRK6 and CRK7 genes play an essential function in extracellular ROS signal transduction in A. thaliana [46]. Various defense responses, including hormone induction, PR gene overexpression and ROS accumulation, are key to grape resistance against gray mold disease [16,47]. In this study, JA signaling pathway defense-related PR gene expression and ROS accumulation in A. thaliana VaCRK2 overexpression lines were significantly higher than in wildtype Col-0 after B. cinerea inoculation (Figure 5, Figures S3 and S6), indicating that VaCRK2 mediates gray mold resistance through JA signaling pathway defense-related PR gene expression and ROS accumulation.
5. Conclusions
This study identified and characterized CRK genes in V. amurensis and validated their role in B. cinerea resistance. The overexpression of VaCRK2 significantly increased the resistance to B. cinerea in A. thaliana, suggesting that VaCRK2 may play a crucial role in gray mold resistance. Furthermore, VaCRK2 expression significantly induced the JA signaling pathway-related genes, PR gene expression and ROS accumulation after B. cinerea inoculation, indicating that VaCRK2 mediates B. cinerea resistance by interacting with the JA signaling pathway and activating oxidative burst. This study provides new insights into the role of CRKs in plant disease resistance and will inform future research on the molecular breeding of grapes for resistance against gray mold disease.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agronomy11081672/s1, Figure S1. Performance of A. thaliana wildtype Col-0 and VaCRK1, VaCRK3, and VaCRK4 overexpression lines following inoculation with B. cinerea. Figure S2. Expression analysis of AtCRK2 in A. thaliana wildtype Col-0 after inoculation with B. cinerea. Figure S3. VaCRK2 cannot mediates resistance to B. cinerea in A. thaliana mutant jar1. Figure S4. JA signaling pathway-related genes expression pattern in A. thaliana wildtype Col-0 and VaCRK2 overexpression lines (OE1, OE2, and OE3) after inoculation with B. cinerea. Figure S5. JA signaling pathway-related genes expression pattern in A. thaliana wildtype Col-0 and VaCRK2 overexpression line1 (OE1) before and after inoculation with B. cinerea. Figure S6. Expression analysis of JA signaling pathway PR genes (PDF1.2, PR3, and PR4) in A. thaliana wildtype Col-0 and VaCRK2 overexpression line 1 (OE1) after inoculation with B. cinerea. Figure S7. Expression analysis of JA signaling pathway PR genes (PDF1.2, PR3, and PR4) in wildtype Col-0 and VaCRK2 overexpression lines (OE1, OE2, and OE3) after inoculation with B. cinerea. Figure S8. Detection of the peroxidase (POD) and catalase (CAT) enzyme activity in A. thaliana wildtype Col-0 and VaCRK2 overexpression line 1 (OE1) after inoculation with B. cinerea. Table S1. Information of the PCR primers used in this study. Table S2. Information of candidate VaCRK genes in Vitis amurensis.
Author Contributions
Writing—original draft preparation, T.L.; methodology, H.G.; data curation, X.T.; conceptualization, D.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31901866).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicble.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection to deployment. Science 2013, 341, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Césari, S.; Kanzaki, H.; Fujiwara, T.; Bernoux, M.; Chalvon, V.; Kawano, Y.; Shimamoto, K.; Dodds, P.; Terauchi, R.; Kroj, T. The NB-LRR proteins RGA4 and RGA5 interact functionally and physically to confer disease resistance. EMBO J. 2014, 33, 1941–1959. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ahn, H.K.; Ding, P.T.; Jones, J.D.G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [PubMed]
- He, Y.X.; Zhou, J.G.; Shan, L.B.; Meng, X.Z. Plant cell surface receptor-mediated signaling—A common theme amid diversity. J. Cell Sci. 2018, 131, jcs209353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.G.; Zhang, D.D.; Zhou, L.; Kong, Z.Q.; Hussaini, A.S.; Wang, D.; Li, J.J.; Short, D.P.G.; Dhar, N.; Klosterman, S.J.; et al. Genome-wide identification and functional analyses of the CRK gene family in cotton reveals GbCRK18 confers Verticillium wilt resistance in Gossypium barbadense. Front Plant Sci. 2018, 9, 1266. [Google Scholar] [CrossRef]
- Tör, M.; Lotze, M.T.; Holton, N. Receptor-mediated signalling in plants: Molecular patterns and programmes. J. Exp. Bot. 2009, 60, 3645–3654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, S.; Waszczak, C.; Hunter, K.; Wrzaczek, M. Bound by fate: The role of reactive oxygen species in receptor-like kinase signaling. Plant Cell 2017, 29, 638–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Du, L.; Chen, Z. Sensitization of defense responses and activation of programmed cell death by a pathogen-induced receptor-like protein kinase in Arabidopsis. Plant Mol. Biol. 2003, 53, 61–74. [Google Scholar] [CrossRef]
- Acharya, B.R.; Raina, S.; Maqbool, S.B.; Jagadeeswaran, G.; Mosher, S.L.; Appel, H.M.; Schultz, J.C.; Klessig, D.F.; Raina, R. Overexpression of CRK13, an Arabidopsis cysteine rich receptor like kinase, results in enhanced resistance to Pseudomonas syringae. Plant J. 2007, 50, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Yadeta, K.A.; Elmore, J.M.; Creer, A.Y.; Feng, B.; Franco, J.Y.; Rufian, J.S.; He, P.; Phinney, B.; Coaker, G. A cysteine-rich protein kinase associates with a membrane immune complex and the cysteine residues are required for cell death. Plant Physiol. 2017, 173, 771–787. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Sun, J.W.; Liu, N.; Sun, X.Z.; Liu, C.J.; Wu, L.Z.; Liu, G.; Zeng, F.L.; Hou, C.Y.; Han, S.F.; et al. A novel cysteine-rich receptor-like kinase gene, TaCRK2, contributes to leaf rust resistance in wheat. Mol. Plant Pathol. 2020, 21, 732–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chern, M.; Xu, Q.F.; Bart, R.S.; Bai, W.; Ruan, D.L.; Sze-To, W.H.; Canlas, P.E.; Jain, R.; Chen, X.W.; Ronald, P.C. A genetic screen identifies a requirement for cysteine-rich-receptor-like kinases in rice NH1 (OsNPR1)-mediated immunity. PLoS Genet. 2016, 12, e1006049. [Google Scholar]
- Rayapuram, C.; Jensen, M.K.; Maiser, F.; Shanir, J.V.; Hornshøj, H.; Rung, J.H.; Gregersen, P.L.; Schweizer, P.; Collinge, D.B. Regulation of basal resistance by a powdery mildew-induced cysteine-rich receptor-like protein kinase in barley. Mol. Plant Pathol. 2012, 13, 135–147. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Veloso, J.; Staats, M.; van Kan, J.A.L. Comparative genomics of plant pathogenic Botrytis species with distinct host specificity. BMC Genom. 2019, 20, 203. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.T.; Liao, Y.X.; Kan, J.Q.; Han, L.; Zheng, Y.H. Response of direct or priming defense against Botrytis cinerea to methyl jasmonate treatment at different concentrations in grape berries. Int. J. Food Microbiol. 2015, 194, 32–39. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Wang, D.J.; Wang, F.; Huang, L.; Tian, X.M.; Nocker, S.V.; Gao, H.; Wang, X.P. Expression of the grape VaSTS19 gene in Arabidopsis improves resistance to powdery mildew and Botrytis cinerea but increases susceptibility to Pseudomonas syringe pv tomato DC3000. Int. J. Mol. Sci. 2017, 18, 2000. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.R.; Qiao, H.B.; Zhao, J.; Wang, X.H.; Tu, M.X.; Guo, C.L.; Wan, R.; Li, Z.; Wang, X.P. The grape VlWRKY3 gene promotes abiotic and biotic stress tolerance in transgenic Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 545. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhang, S.L.; Singer, S.D.; Yin, X.J.; Yang, J.H.; Wang, Y.J.; Wang, X.P. Expression of the grape VqSTS21 gene in Arabidopsis confers resistance to osmotic stress and biotrophic pathogens but not Botrytis cinerea. Front. Plant Sci. 2016, 7, 1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.R.; Tu, M.X.; Wang, X.H.; Zhao, J.; Wan, R.; Li, Z.; Wang, Y.J.; Wang, X.P. Ectopic expression of a grape aspartic protease gene, AP13, in Arabidopsis thaliana improves resistance to powdery mildew but increases susceptibility to Botrytis cinerea. Plant Sci. 2016, 248, 17–27. [Google Scholar] [CrossRef]
- Wang, Y.; Xin, H.P.; Fan, P.G.; Zhang, J.S.; Liu, Y.B.; Dong, Y.; Wang, Z.M.; Yang, Y.Z.; Zhang, Q.; Ming, R.; et al. The genome of Shanputao (Vitis amurensis) provides a new insight into cold tolerance of grapevine. Plant J. 2021, 105, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Accelerated profile HMM searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, T.N.; Brunak, S.; Von, H.G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von, H.G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.; Jones, P.; Mitchell, A.; Apweiler, R.; Attwood, T.K.; Bateman, A.; Bernard, T.; Binns, D.; Bork, P.; Burge, S.; et al. InterPro in 2011: New developments in the family and domain prediction database. Nucleic Acids Res. 2012, 40, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Memelink, J. Regulation of gene expression by jasmonate hormones. Phytochemistry 2009, 70, 1560–1570. [Google Scholar] [CrossRef]
- Yeh, Y.H.; Chang, Y.H.; Huang, P.Y.; Huang, J.B.; Zimmerli, L. Enhanced Arabidopsis pattern-triggered immunity by overexpression of cysteine-rich receptor-like kinases. Front. Plant Sci. 2015, 6, 322. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Wei, R.; Lin, Q.; Li, J.R.; Wei, X.N.; Zhang, Z.Y. Isolation and characterization of a novel wheat cysteine-rich receptor-like kinase gene induced by Rhizoctonia cerealis. Sci. Rep. 2013, 3, 3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berrabah, F.; Bourcy, M.; Eschstruth, A.; Cayrel, A.; Guefrachi, I.; Mergaert, P.; Wen, J.Q.; Jean, V.; Mysore, K.S.; Gourion, B.; et al. A nonRD receptor-like kinase prevents nodule early senescence and defense-like reactions during symbiosis. New Phytol. 2014, 203, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.H.; Jiang, Z.Y.; Bi, G.Z.; Nomura, K.; Liu, M.H.; Wang, Y.P.; Cai, B.Y.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [PubMed]
- Bao, F.; Ding, A.Q.; Cheng, T.R.; Wang, J.; Zhang, Q.X. Genome-wide analysis of members of the WRKY gene family and their cold stress response in Prunus mume. Genes 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickman, R.; Van Verk, M.C.; Van Dijken, A.J.H.; Mendes, M.P.; Vroegop-Vos, I.A.; Caarls, L.; Steenbergen, M.; Nagel, I.V.D.; Wesselink, G.J.; Jironkin, A.; et al. Architecture and dynamics of the jasmonic acid gene regulatory network. Plant Cell 2017, 29, 2086–2105. [Google Scholar] [CrossRef] [Green Version]
- Balagué, C.; Gouget, A.; Bouchez, O.; Souriac, C.; Haget, N.; Boutet-Mercey, S.; Govers, F.; Roby, D.; Canut, H. The Arabidopsis thaliana lectin receptor kinase LecRK-I.9 is required for full resistance to Pseudomonas syringae and affects jasmonate signaling. Mol. Plant Pathol. 2017, 18, 937–948. [Google Scholar] [CrossRef]
- Liu, M.Y.; Zhang, Q.X.; Wang, C.; Meng, T.Q.; Wang, L.N.; Chen, C.H.; Ren, Z.H. CsWRKY10 mediates defence responses to Botrytis cinerea infection in Cucumis sativus. Plant Sci. 2020, 300, 110640. [Google Scholar] [CrossRef]
- Liu, M.Y.; Zhang, Z.; Xu, Z.X.; Wang, L.N.; Chen, C.H.; Ren, Z.H. Overexpression of SlMYB75 enhances resistance to Botrytis cinerea and prolongs fruit storage life in tomato. Plant Cell Rep. 2021, 40, 43–58. [Google Scholar] [CrossRef]
- Berr, A.; McCallum, E.J.; Alioua, A.; Heintz, D.; Heitz, T.; Shen, W.H. Arabidopsis histone methyltransferase SET DOMAIN GROUP8 mediates induction of the jasmonate/ethylene pathway genes in plant defense response to necrotrophic fungi. Plant Physiol. 2010, 154, 1403–1414. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.Y.; Wu, S.G.; Jing, Y.F.; Chen, L.G.; Yu, D.Q. Phytochrome B regulates jasmonic acid-mediated defense response against Botrytis cinerea in Arabidopsis. Plant Divers. 2021, in press. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [Green Version]
- Caplan, J.; Padmanabhan, M.; Dinesh-Kumar, S.P. Plant NB-LRR immune receptors: From recognition to transcriptional reprogramming. Cell Host Microbe. 2008, 3, 126–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Young, B.J.; Innes, R.W. Plant NBS-LRR proteins in pathogen sensing and host defense. Nat. Immunol. 2006, 7, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Han, X.M.; Shi, R.; Yang, G.Y.; Qi, L.W.; Wang, R.G.; Li, G.J. Arabidopsis cysteine-rich receptor-like kinase 45 positively regulates disease resistance to Pseudomonas syringae. Plant Physiol. Biochem. 2013, 73, 383–391. [Google Scholar] [CrossRef]
- Idänheimo, N.; Gauthier, A.; Salojärvi, J.; Siligato, R.; Brosché, M.; Kollist, H.; Mähönen, A.P.; Kangasjärvi, J.; Wrzaczek, M. The Arabidopsis thaliana cysteine-rich receptor-like kinases CRK6 and CRK7 protect against apoplastic oxidative stress. Biochem. Biophys. Res. Commun. 2014, 445, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.U.; Hanif, M.; Wan, R.; Hou, X.Q.; Ahmad, B.; Wang, X.P. Screening Vitis genotypes for responses to Botrytis cinerea and evaluation of antioxidant enzymes, reactive oxygen species and jasmonic acid in resistant and susceptible hosts. Molecules 2019, 24, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
VaCRK2 mediates resistance to Botrytis cinerea in Arabidopsis thaliana. (A) Phenotype of A. thaliana wildtype Col-0 and three VaCRK2 overexpression lines (OE1, OE2 and OE3) after inoculation with B. cinerea. (B) Relative disease index of A. thaliana wildtype Col-0 and three VaCRK2 OE lines. (C) RT-qPCR analysis of relative fungal biomass in A. thaliana wildtype Col-0 and three VaCRK2 OE lines. Error bars represent the standard error of the mean of three independent replicates, asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests). Mock (M), Botrytis cinerea (Bc).
Figure 1.
VaCRK2 mediates resistance to Botrytis cinerea in Arabidopsis thaliana. (A) Phenotype of A. thaliana wildtype Col-0 and three VaCRK2 overexpression lines (OE1, OE2 and OE3) after inoculation with B. cinerea. (B) Relative disease index of A. thaliana wildtype Col-0 and three VaCRK2 OE lines. (C) RT-qPCR analysis of relative fungal biomass in A. thaliana wildtype Col-0 and three VaCRK2 OE lines. Error bars represent the standard error of the mean of three independent replicates, asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests). Mock (M), Botrytis cinerea (Bc).

Figure 2.
VaCRK2 protein structure and its subcellular localization. (A) Schematic representation of VaCRK2 protein structure. A signal peptide (SP) from 1 to 25 amino acid (aa), two Stress-antifung (DUF26) domains from 35 to 129 aa and 143 to 241 aa, a transmembrane (TM) domain from 266 to 288 aa and a kinase domain from 347 to 596 aa. (B) Subcellular localization of VaCRK2 protein. p35S:VaCRK2 and p35S:GFP vectors were introduced into tobacco epidermal cells by Agrobacterium infiltration.
Figure 2.
VaCRK2 protein structure and its subcellular localization. (A) Schematic representation of VaCRK2 protein structure. A signal peptide (SP) from 1 to 25 amino acid (aa), two Stress-antifung (DUF26) domains from 35 to 129 aa and 143 to 241 aa, a transmembrane (TM) domain from 266 to 288 aa and a kinase domain from 347 to 596 aa. (B) Subcellular localization of VaCRK2 protein. p35S:VaCRK2 and p35S:GFP vectors were introduced into tobacco epidermal cells by Agrobacterium infiltration.

Figure 3.
Expression patterns of VaCRK2 under different treatments. (A) Expression analysis of VaCRK2 in resistant Vitis amurensis cv. Shuangyou (SY) and susceptible V. vinifera cv. Red Globe (RG) after inoculation with B. cinerea. Expression analysis of VaCRK2 in cv. Shuangyou in response to (B) methyl jasmonic acid (MeJA), (C) salicylic acid (SA), (D) ethephon (ETH) and (E) abscisic acid (ABA). Error bars represent the standard error of the mean of three independent replicates, asterisks (*) and (**) indicate significant difference at p < 0.05 and p < 0.01, respectively (Student’s t-tests). Botrytis cinerea (Bc).
Figure 3.
Expression patterns of VaCRK2 under different treatments. (A) Expression analysis of VaCRK2 in resistant Vitis amurensis cv. Shuangyou (SY) and susceptible V. vinifera cv. Red Globe (RG) after inoculation with B. cinerea. Expression analysis of VaCRK2 in cv. Shuangyou in response to (B) methyl jasmonic acid (MeJA), (C) salicylic acid (SA), (D) ethephon (ETH) and (E) abscisic acid (ABA). Error bars represent the standard error of the mean of three independent replicates, asterisks (*) and (**) indicate significant difference at p < 0.05 and p < 0.01, respectively (Student’s t-tests). Botrytis cinerea (Bc).

Figure 4.
VaCRK2 regulates the jasmonic acid signaling pathway-related genes to confer resistance to Botrytis cinerea in A. thaliana. Wildtype (Col-0) and VaCRK2 overexpression lines (OE1, OE2 and OE3) were inoculated with B. cinerea, and leaf samples were collected for RT-qPCR analysis 24 h post-inoculation. Error bars represent the standard error of three independent replicates, asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests).
Figure 4.
VaCRK2 regulates the jasmonic acid signaling pathway-related genes to confer resistance to Botrytis cinerea in A. thaliana. Wildtype (Col-0) and VaCRK2 overexpression lines (OE1, OE2 and OE3) were inoculated with B. cinerea, and leaf samples were collected for RT-qPCR analysis 24 h post-inoculation. Error bars represent the standard error of three independent replicates, asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests).

Figure 5.
VaCRK2 mediates Botrytis cinerea resistance in Arabidopsis thaliana by activating pathogenesis-related (PR) genes and oxidative burst. (A) Expression analysis of JA signaling pathway PR genes (PDF1.2, PR3 and PR4) in wildtype Col-0 and VaCRK2 overexpression line 1 (OE1) after inoculation with B. cinerea. (B) Phenotype of wildtype Col-0 and VaCRK2 OE1 after inoculation with B. cinerea followed by diaminobenzidine (DAB) staining. (C) The browning intensity of wildtype Col-0 and VaCRK2 OE1 leaves after inoculation with B. cinerea followed by DAB staining. Error bars represent the standard error of the mean of three independent biological replicates, and asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests). Mock (M), Botrytis cinerea (Bc).
Figure 5.
VaCRK2 mediates Botrytis cinerea resistance in Arabidopsis thaliana by activating pathogenesis-related (PR) genes and oxidative burst. (A) Expression analysis of JA signaling pathway PR genes (PDF1.2, PR3 and PR4) in wildtype Col-0 and VaCRK2 overexpression line 1 (OE1) after inoculation with B. cinerea. (B) Phenotype of wildtype Col-0 and VaCRK2 OE1 after inoculation with B. cinerea followed by diaminobenzidine (DAB) staining. (C) The browning intensity of wildtype Col-0 and VaCRK2 OE1 leaves after inoculation with B. cinerea followed by DAB staining. Error bars represent the standard error of the mean of three independent biological replicates, and asterisks (**) indicate significant difference at p < 0.01 (Student’s t-tests). Mock (M), Botrytis cinerea (Bc).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, T.; Gao, H.; Tang, X.; Gong, D. VaCRK2 Mediates Gray Mold Resistance in Vitis amurensis by Activating the Jasmonate Signaling Pathway. Agronomy 2021, 11, 1672. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081672
AMA Style
Li T, Gao H, Tang X, Gong D. VaCRK2 Mediates Gray Mold Resistance in Vitis amurensis by Activating the Jasmonate Signaling Pathway. Agronomy. 2021; 11(8):1672. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081672
Chicago/Turabian StyleLi, Tinggang, Huanhuan Gao, Xiaoning Tang, and Dongying Gong. 2021. "VaCRK2 Mediates Gray Mold Resistance in Vitis amurensis by Activating the Jasmonate Signaling Pathway" Agronomy 11, no. 8: 1672. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081672
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.