
Modeling the Effect of Temperature on Ginger and Turmeric Rhizome Sprouting



Environmental Horticulture Department, Institute of Food and Agricultural Sciences (IFAS), University of Florida, Gainesville, FL 32611-0670, USA
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(10), 1931; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101931
Submission received: 30 August 2021
/
Revised: 21 September 2021
/
Accepted: 22 September 2021
/
Published: 26 September 2021
(This article belongs to the Special Issue A New Decade of Horticultural and Medicinal Plants Cultivation)
Abstract
:Ginger and turmeric are tropical plant species with medicinal, beverage, and edible uses. Both species are typically propagated using seed rhizomes that often lack uniformity when sprouting, ultimately affecting the transplant growth and quality. Our objectives were to (1) develop a model to predict the effect of temperature on rhizome sprouting and transplant growth and (2) characterize the morphological factors affecting the sprouting of ginger and turmeric rhizomes. Two experiments were conducted where the rhizomes were placed in plastic bags with a moist substrate inside dark incubator chambers. Five temperature treatments (21, 25, 27, 30, and 32 °C) were used for calibrating the model, and six temperature treatments (14, 20, 25, 30, 35, and 40 °C) were used in the validation phase. The number of days for rhizomes to develop 1- and 5-cm sprouts were counted; after which, the total number of sprouts, total leaf length, and root quality were measured. A nonlinear regression analysis was used to develop temperature–response curves. Ginger and turmeric had optimal sprouting temperatures of 27.5 and 30.1 °C, respectively. Temperatures close to the optimal reduced the time to sprout and to subsequently reach the transplant stage. No sprouting was observed at 14 °C, and the minimum temperature to develop 5-cm sprouts was estimated at slightly above 17 °C in both species. Temperatures above 32 °C resulted in tissue damage and rhizome loss. The results from this study show the potential to produce uniform ginger and turmeric transplants using temperature treatments that accelerate sprouting.
1. Introduction
Ginger (Zingiber officinale Roscoe) and turmeric (Curcuma longa L.) consumption in the United States (U.S.) is rising partly due to an increased consumer interest in their medicinal properties [1,2,3]. Both crops are described as “superfoods” due to the high concentration of bioactive compounds and phytochemicals in their rhizomes. Ginger has more than 400 health-related compounds, including paradols, terpenoids, shogaols, and gingerols [4]. Over 230 compounds have been identified in turmeric, including various phenolics and terpenoids (curcuminoids) [5], with antiviral, antibacterial, antidiabetic, antioxidant, and anti-inflammatory properties [6,7,8]. The unique flavor, color, and preservative characteristics of these rhizomes also make them attractive supplements for food and beverage uses [9]. Currently, commercial production is primarily limited to Asian and South American countries, and a few small-scale farms in Hawaii and the southern U.S. [10]. Although interest in locally produced medicinal plant products is increasing, protocols for the propagation and production of these two crops in different U.S. climates are lacking [11,12].
Ginger and turmeric are typically propagated using tissue-cultured transplants that are low-yielding on the first cropping cycle [13] or from small pieces (10–100 g) of vegetative rhizomes, often referred to as “seed rhizomes” [14,15,16]. Seed rhizomes have certain disadvantages for commercial production, including nonuniform and slow sprouting, which affect the transplant growth and sometimes affect the final yield of plants in the field or containers [17]. The breaking of bud dormancy occurs in response to physiological and biochemical changes in the rhizome, including the reallocation of water, metabolites, and plant hormones and increases in the synthesis of sugars, all of which are heavily affected by various environmental factors [18].
Temperature and plant hormones such as cytokinins (benzyladenine, benzylaminopurine, and kinetin) and ethylene are common factors that regulate the sprouting of rhizomes, tubers, bulbs, and other storage organs in plants [14,15,19,20,21,22,23,24,25]. However, limited information is available on the effect of temperature on ginger and turmeric rhizome sprouting and the subsequent transplant growth. Kaushal et al. [26] reported that ginger rhizomes can be stored and kept dormant under temperature ranges between 12 and 14 °C, but sprouting occurs at temperatures above 16 °C. Paull and Cheng [27] reported that ginger rhizomes are affected by chilling injuries when kept at temperatures below 12 °C. The storage of turmeric rhizomes at 12–14 °C has been recommended to delay sprouting, whereas temperature ranges between 27 and 30 °C accelerate sprouting. Lee at al. [28] showed that, under 10–15 °C, turmeric rhizomes can be kept dormant, whereas 24 °C promotes sprouting. This information is helpful when developing temperature models, which are useful to predict developmental processes such as bud sprouting and the breaking of dormancy in a wide range of species [29,30]. These models could be used as a decision–support tool to inform growers about proper conditions to store ginger and turmeric rhizomes and to induce uniform rhizome sprouting [31,32,33].
The morphological characteristics of seed rhizomes (e.g., weight, thickness, and length) have also been shown to affect the growth and yield of ginger and turmeric plants [34]. In general, larger rhizomes produce a higher yield than smaller rhizomes, most likely because of the larger accumulation of reserve compounds and a greater number of buds that grow on to become shoots [34,35,36,37]. However, information is not available correlating the rhizome morphology and subsequent sprouting, which could provide further insight to explain how the propagation stage affects the growth and yield of these crops.
The objectives of this study were to (1) develop a model to predict the effect of temperature on rhizome sprouting and (2) characterize the morphological factors affecting the sprouting of ginger and turmeric rhizomes. Two experiments were conducted over time. The calibration experiment included temperature treatments between 21 and 32 °C (a range expected to include rapid sprouting conditions) in order to precisely identify an optimum sprouting temperature. A separate validation experiment generated a dataset with a broader range of temperatures in order to confirm the repeatability of the temperature response equation generated from the calibration experiment. We hypothesized that rhizomes of both species would be induced to sprout faster and would produce more and larger shoots under warmer temperatures, but the optimum temperatures would be species-specific. Furthermore, we hypothesized that larger and heavier rhizomes would sprout faster than smaller and lighter rhizomes.
2. Materials and Methods
2.1. Plant Material
Seed rhizomes of “Bubba Blue” and “Madonna” ginger (Hawaii Clean Seed LLC., Pahoa, HI, USA) and “Indira Yellow” and “Hawaiian Red” turmeric (Hawaii Clean Seed LLC.), which are varieties commonly grown by commercial growers in the U.S., were harvested on 18 March 2020 and 16 June 2020 for the validation and calibration experiments, respectively. Mother plants from which the rhizomes were harvested had previously been grown for 28 weeks in a polycarbonate greenhouse in Gainesville, FL, USA. The rhizomes were placed in empty black plastic trays (21 cm (h) × 27.8 cm (w) × 6.2 cm (d); T.O. Plastics, Inc. Clearwater, MN, USA) and stored in a dark air-conditioned room for approx. 91 d. The daily ambient temperature and relative humidity (RH) in the room were set to 14 °C and 60%, respectively. The environmental conditions were monitored with a temperature/RH data logger (HOBO MX1101; Onset, Bourne, MA, USA). After storage, rhizomes ranging from 2.0 to 4.5 cm in length and 50 to 150 g were used in the experiments.
2.2. Treatments
In both experiments, individual rhizomes were placed inside 16.5 cm × 14.9 cm transparent plastic bags (Ziploc®; SC Johnson & Son, Racine, WI, USA) filled with 300 mL of a horticultural grade substrate composed of (v/v) 79–87% peat moss, 10–14% perlite, and 3–7% vermiculite (Pro-Mix BX general purpose; Premier Tech Horticulture, Quakertown, PA, USA) previously moistened with 80 mL of tap water with 0.4-dS·m−1 electrical conductivity, 8.3 pH, and 31.2-mg·L‒1 calcium carbonate. Twelve replicate bags were placed inside dark incubator chambers (74.5-cm (h) × 59.5 cm (w) × 55.5 cm (d); IB-15G; Jeiotech, Daejeon, Republic of Korea), which were used as treatment compartments. Five temperature treatments (21, 25, 27, 30, and 32 °C) were evaluated in the calibration experiment, which started on 16 September 2020. Six temperature treatments (14, 20, 25, 30, 35, and 40 °C) were used in the validation experiment, which started on 17 June 2020. Both experiments lasted 20 weeks, and throughout each experiment, the temperature was measured using a temperature datalogger (HOBO MX1101; Onset) placed in the center of each incubator. The ambient temperatures measured in the five incubators were (daily average ± standard deviation) 20.5 ± 0.4, 24.9 ± 0.2, 27.1 ± 0.5, 29.7 ± 0.4, and 32.4 ± 0.3 °C for the calibration experiment and 14.1 ± 0.4, 20.4 ± 0.3, 25.0 ± 0.3, 29.8 ± 0.4, 34.6 ± 0.5, and 40.2 ± 0.3 °C for the validation experiment. The experimental design was completely randomized, with one rhizome per replicate bag as the experimental unit. Each treatment consisted of 12 replicate bags. To maintain the substrate moisture, 15 mL of tap water were added weekly to each replicate bag.
2.3. Data Collected
Before starting each experiment, the number of buds per rhizome were counted, and the rhizomes were weighed using a portable scale (PC 4400; Mettler-Toledo International, Greifensee, Switzerland). The length, width, and thickness of each rhizome were measured with a digital caliper (Ultra-Cal Mark III; Fred V. Fowler Co, Newton, MA, USA). All bags were checked every other day to record the number of days until the rhizomes developed 1- and 5-cm sprouts. To compare the growth and development in response to temperature, the number and height of the sprouts, root number, primary root length, and root quality were measured three weeks after the start of each experiment. In the subjective 0–5 scale used for root quality determination, 0 = no roots, 1 = ≤ three root, 2 = ≤ six roots, 3 = ≤ eight roots, 4 = ≤ 10 roots, and 5 = ≤ 15 roots.
Once the rhizomes developed a 5-cm sprout, they were moved out of the incubator chambers and immediately transplanted into 15-cm plastic containers (VTD 15 US; Poppelmann Plastics US, Claremont, NC, USA) filled with the same substrate previously described. The containers were randomly placed on benches (4.6 m × 1.8 m) inside a polycarbonate greenhouse compartment in Gainesville, FL, USA. The number of days until the transplants that reached a commercial transplant stage (20 and 30 cm heights for ginger and turmeric, respectively) were counted. The air temperature, RH, and daily light integral (DLI) were monitored with temperature and RH probes (HMP60-L; Campbell Scientific, Logan, UT, USA) and quantum sensors (SQ512; Apogee Instruments Inc., Logan, UT, USA) interfaced to a datalogger (CR1000; Campbell Scientific) and multiplexer (AM16/32B; Campbell Scientific), respectively. Each bench had one temperature/RH probe and one quantum sensor placed above the canopy height, and the measurements were made every 30 s and recorded at 60-min intervals. The day and night air temperatures and solar DLI measured in the greenhouse were 25.5 ± 2.6 °C, 22.6 ± 2.2 °C, and 11.4 ± 4.3 mol·m‒2·d‒1 in the calibration experiment and 24.7 ± 1.9 °C, 21.5 ± 1.6 °C, and 9.8 ± 3.6 mol·m‒2·d‒1 in the validation experiment, respectively. All the plants were grown under natural sunlight, and the night interruption lighting was delivered by red + white + far-red light-emitting diode lamps (Arize Greenhouse Pro; GE Lighting, Cleveland, OH, USA), providing a total photon flux density of 4.5 µmol·m‒2·s‒1 from 10 p.m. to 2 a.m. The transplants were irrigated with tap water as needed.
2.4. Temperature Modeling
Temperature–response curves were developed to model the effect of temperature on rhizome sprouting. The curves were fitted to the measured data in the calibration and validation experiments using the Landsberg function, which is an asymmetric, peak-shaped curve [38,39]:
where
and
R = A (T − Tₘᵢₙ)(Tₘₐₓ − T)ᴮ
A = Rₘₐₓ/(Tₒₚₜ − Tₘᵢₙ)(Tₘₐₓ − Tₒₚₜ)ᴮ
B = (Tₘₐₓ − Tₒₚₜ)/(Tₒₚₜ − Tₘᵢₙ)
All four parameters have biological meaning, where Tₘᵢₙ is the minimum temperature (°C) for sprouting, Tₘₐₓ is the maximum temperature (°C) for sprouting, Rₘₐₓ is the maximum rate (days) for sprouting, and Tₒₚₜ is the optimum sprouting temperature, where the developmental rate (R) equals Rₘₐₓ. Below Tmin and above Tₘₐₓ, R equals zero, and the calculated B value affects the skew.
The estimates for Tₘᵢₙ, Tₘₐₓ, Tₒₚₜ, and Rₘₐₓ based on the measured plant temperature were obtained using the nonlinear regression analysis procedure (PROC NLIN) of SAS (Version 9.2; SAS Institute, Cary, NC, USA) based on the inverse of days to 5-cm sprouting from the calibration dataset, including only the temperature treatments that induced 5-cm sprouting. The Tₘᵢₙ, Tₘₐₓ, Tₒₚₜ, and Rₘₐₓ parameters estimated from the calibration experiment were then used to validate the model with the validation dataset. The statistically fit curve was compared to the mean temperature and 95% confidence intervals for each temperature treatment, where overlapping 95% confidence intervals and mean estimates from the Landsberg function curve would indicate that the temperature model was valid for predicting rhizome sprouting.
2.5. Other Data Analyses
To identify the effects of temperature on the other aspects of rhizome growth and development, data from the calibration and validation experiments were tested for the assumptions of normality and homoscedasticity, and when they were not met, logarithmic transformation was used. The data were subjected to an analysis of variance (ANOVA) using R version 3.6.1 [40]. The least-square treatment means were compared using Tukey’s honestly significant difference (HSD) test (p = 0.05) in the Agricolae package of R [41]. The morphological data were also subjected to ANOVA and Tukey’s HSD tests (p = 0.05) using R. Pearson’s correlations among the morphological data were calculated, and an analysis of covariance (ANCOVA) was used to test if sprouting data from the calibration experiment were influenced by morphological parameters, where 5-cm sprouting was the dependent variable, temperature treatments were the independent variables, and the morphological parameters of the seed rhizomes were the covariates.
3. Results and Discussion
3.1. Model Calibration
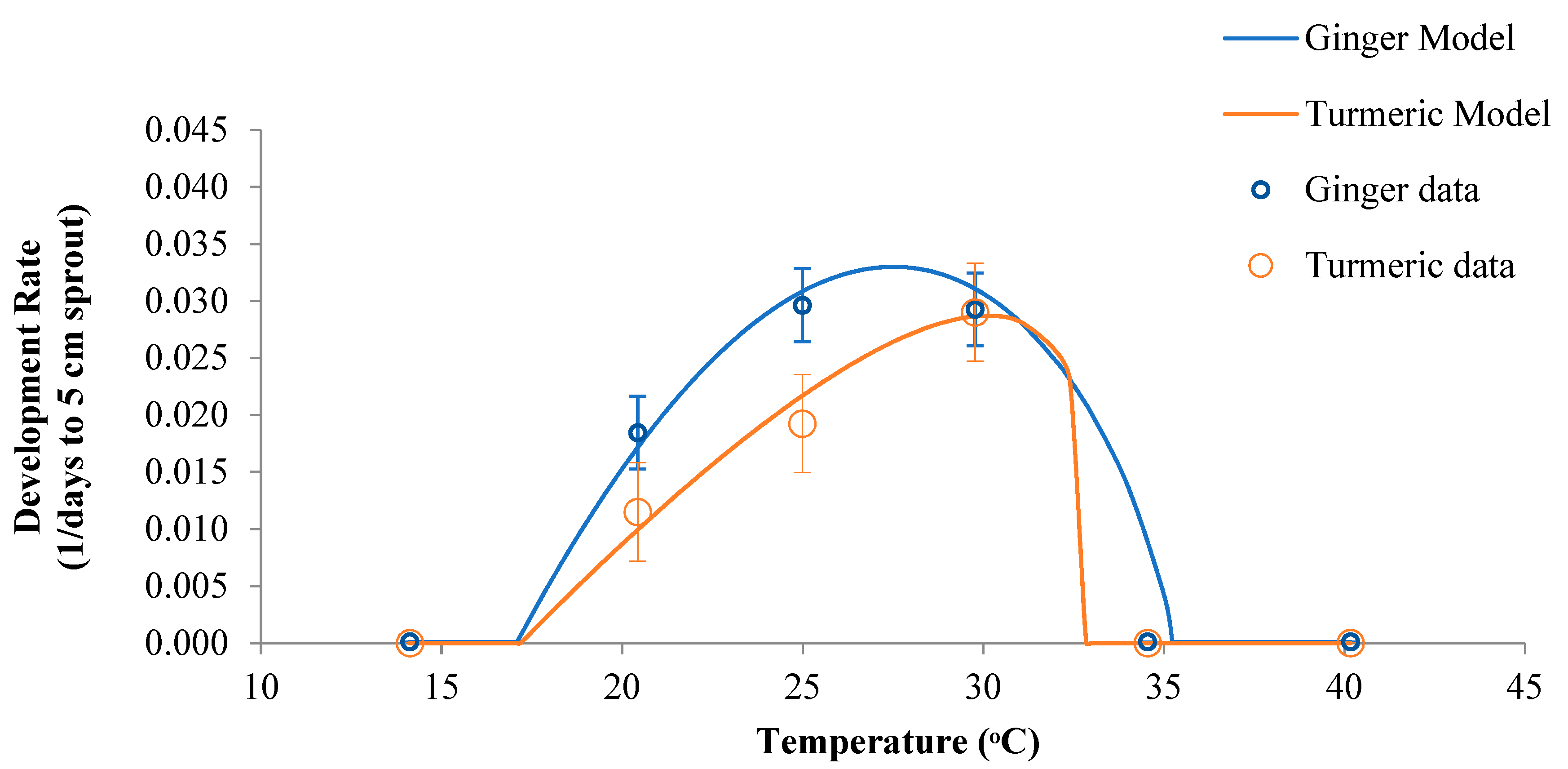
The nonlinear regression of Equation (1) with calibration data for 1/day to a 5-cm sprout resulted in the parameter estimates described in Table 1. The 95% confidence intervals were less than 10% of the parameter estimates, indicating that the model was not overparameterized for the dataset. Comparing the observed and estimated data for the individual rhizomes resulted in an R2 of 0.728 for ginger and 0.703 for turmeric. The 95% confidence intervals around the mean estimates for each temperature overlapped with the Landsberg function curve, indicating that the function was an adequate empirical representation of the sprouting behavior of ginger and turmeric rhizomes as a function of the temperature (Figure 1).
Ginger and turmeric had similar estimates of the minimum temperature below which sprouting would not occur (Tₘᵢₙ), with corresponding temperatures of 17.1 or 17.2 °C, respectively (Table 1). These estimates are above the recommended storage temperature for ginger and turmeric rhizomes, which was described as 14 °C by Policegoudra et al. [42]. The Tₒₚₜ was higher for turmeric (30.1 °C) than ginger (27.5 °C), and the 95% confidence intervals did not overlap between species. However, the Tₘₐₓ estimates overlapped, with corresponding temperatures of 35.2 °C for ginger and 32.8 °C for turmeric rhizomes. The estimated Tₘₐₓ for turmeric was only slightly higher than the highest temperature used in this study at 32.3 °C. Therefore, Tₘₐₓ may have been underestimated by the Landsberg function for turmeric.
The Rₘₐₓ estimates, which represent the maximum developmental rate, were 0.0342 d–1 for ginger and 0.0318 d–1 for turmeric, and the 95% confidence intervals overlapped (Table 1). These parameters can be inverted to estimate a minimum time to sprout, which resulted in 29 and 31 d for ginger and turmeric rhizomes, respectively. These values corresponded with the observed sprouting durations reported in Table 1. The optimum sprouting temperatures increased the physiological activity, protein and enzyme synthesis, and accelerate rhizome metabolism [43]. In contrast, storing rhizomes at higher or lower temperatures than the Tₒₚₜ could lengthen the propagation cycle when using ginger and turmeric seed rhizomes.
3.2. Model Validation
Comparing the observed and estimated data for individual rhizomes resulted in an R2 of 0.774 for ginger and 0.795 for turmeric. When the parameter estimates for Tₘᵢₙ, Tₘₐₓ, Tₒₚₜ, and Rₘₐₓ from the calibration dataset were used to predict the temperature response with the broader temperature range in the validation experiment, the 95% confidence intervals around the mean estimates for each temperature overlapped with the Landsberg function curve (Figure 2). This overlap supports the validity of the temperature model, explaining the effect of temperature on ginger and turmeric rhizome sprouting.
According to our model, the optimum temperatures for sprouting for ginger and turmeric are 27.5 and 30.1 °C, respectively, indicating that the optimum sprouting of turmeric occurs at slightly warmer temperatures than ginger. No sprouting was observed under 14 or 40 °C. The rhizomes at both 35 and 40 °C showed signs of tissue damage. These findings suggest that temperatures ≤ 14 °C are not sufficient to break the dormancy of ginger and turmeric rhizomes, which are consistent with the previously recommended storage temperatures of 12–14 °C [44].
Similar to our findings, Chen at al. [45] and Lee et al. [28] reported limited sprouting in ginger and turmeric rhizomes under 15 °C. Ishimine et al. [43] showed that the optimal temperature range for turmeric rhizome sprouting was between 25 and 35 °C and that temperatures below 10 °C or above 40 °C inhibit turmeric rhizome sprouting. Chen at al. [45] reported the highest rhizome sprouting under 25 °C. Our results also indicated that temperatures ≥35 °C are too high for sprouting, leading to tissue damage and rhizome loss.
Studies have shown that the maximum development rate (Rₘₐₓ in the Landsberg function) in modified stems or storage organs such as rhizomes can be affected by the temperature and RH, soil moisture content, plant genetics, various physiological and hormonal responses, rhizome morphology and age, and storage time, among other factors [45,46,47]. It is likely that, by evaluating several of these factors and their interactions, the optimum conditions for the parameters other than temperature could be identified for ginger or turmeric rhizome sprouting. Further optimizing Rₘₐₓ could result in lower values than the ones reported in our study. However, our findings provided a good starting point to better understand the temperature response for storage and sprouting ginger and turmeric seed rhizomes.
In terms of decision support to evaluate the effect of nonoptimum temperatures, the model can be generalized to other batches of rhizomes by dividing the Rmax by R in Equation (1) in order to standardize the curve to a minimum sprouting time of 100%. For example, ginger at the optimum temperature of 27.5 °C (Topt from Table 1) would require a standard 100% minimum time to sprout. At 25 °C, the value for ginger would be 107% of the minimum time (requiring only 7% more time to sprout compared with at 27.5 °C). However, ginger plants at 20 °C would require 216% of the minimum time (more than double the number of days to sprout at 20 °C compared with 27.5 °C).
3.3. Other Temperature Effects on Rhizome Growth and Development
For both species, the most extreme temperature treatments (21 and 32 °C in the calibration experiment and 14 and 40 °C in the validation experiment) generally resulted in a decrease in root and shoot growth compared with the intermediate temperatures (Table 2). In the calibration experiment, ginger rhizomes under 25, 27, and 30 °C had a higher rooting index (from 4.0 to 4.5), produced 261–464% more roots per plant, and took less time to produce a 1-cm sprout (from 5 to 12 fewer days), 5-cm sprout (from 10 to 19 fewer days), and to reach a transplant stage (from 5 to 7 fewer days), respectively, compared with rhizomes under 21 and 32 °C (Table 1). In addition, sprouts of ginger rhizomes under 27 and 30 °C were 96–120% taller than those under any other treatment.

{kind=link}
{kind=link}
Table 2.
Seed rhizome growth parameters measured from “Bubba Blue” and “Madonna” ginger sprouted under different temperatures inside dark incubators and then grown in a greenhouse in two experimental phases.
Table 2.
Seed rhizome growth parameters measured from “Bubba Blue” and “Madonna” ginger sprouted under different temperatures inside dark incubators and then grown in a greenhouse in two experimental phases.
| Experiment | Sprouts per Plant (No.) | Average Sprout Height (mm) | Roots per Plant (No.) | Primary Root Length (mm) | Rooting Index (1 to 5) | Days to 1-cm Sprout | Days to 5-cm Sprout | Transplant Stage in Greenhouse (days) |
|---|---|---|---|---|---|---|---|---|
| Calibration | ||||||||
| 21 °C | 7.6 | 5.1 b z | 2.8 b | 7.6 c | 0.8 b | 26.4 a | 51.8 a | 18.3 a |
| 25 °C | 7.9 | 8.2 b | 10.1 a | 67.7 b | 4.5 a | 13.4 c | 32.5 c | 14.8 b |
| 27 °C | 7.7 | 10.0 a | 12.5 a | 69.2 b | 4.3 a | 13.6 c | 28.9 c | 12.9 b |
| 30 °C | 9.0 | 11.2 a | 14.1 a | 88.4 a | 4.0 a | 14.2 c | 31.7 c | 15.1 b |
| 32 °C | 8.0 | 6.6 b | 2.5 b | 16.0 c | 1.2 b | 19.6 b | 42.2 b | 19.6 a |
| Treatment (T) | NS | ** | *** | *** | ** | *** | *** | ** |
| Variety (V) | NS | NS | NS | NS | NS | NS | NS | NS |
| T × V | NS | NS | NS | NS | NS | NS | NS | NS |
| Validation | ||||||||
| 14 °C | - | - | - | - | - | 65.0 a | - | - |
| 20 °C | 5.0 bc | 3.2 b | 1.7 c | 3.2 b | 0.3 b | 27.7 b | 54.2 a | 17.2 |
| 25 °C | 6.5 b | 4.7 b | 7.2 b | 58.3 a | 3.5 a | 12.2 c | 33.8 b | 14.1 |
| 30 °C | 9.8 a | 8.8 a | 10.5 a | 71.6 a | 3.3 a | 16.1 c | 34.2 b | 16.7 |
| 35 °C | 3.9 c | 1.9 c | - | - | - | - | - | - |
| 40 °C | - | - | - | - | - | - | - | - |
| T | *** | *** | *** | ** | *** | *** | *** | ** |
| V | NS | NS | NS | NS | NS | NS | NS | NS |
| T × V | NS | NS | NS | NS | NS | NS | NS | NS |
z Means within columns followed by the same letter are not different based on Tukey’s honestly significant different test at p ≤ 0.05 (n = 12). ***, **, and NS indicate statistical significance at the 0.001 and 0.01 p ≤ levels or not significant, respectively. - indicates no data.
Turmeric rhizomes under 25, 27, and 30 °C in the calibration experiment produced 11–70% more roots per plant and had a higher rooting index (from 2.1 to 3.2, respectively) than those under 21 and 32 °C (Table 3). Sprouts of turmeric rhizomes under 27, 30, and 32 °C were 18–116% taller than those under 21 and 25 °C, and the time to produce 1-cm sprouts was reduced with the temperature, ranging from 8 to 38 fewer days in the three warmer temperatures. Similarly, the days to produce 5-cm sprouts and to reach the transplant stage were 14–51, and 6–15, respectively, which were fewer in turmeric rhizomes under 25, 27, and 30 °C compared with those under 21 and 32 °C. In both species, there were no treatment differences in the number of sprouts produced per rhizome.
Rhizomes from both species under 14 and 40 °C did not produce roots or sprouts in the validation experiment (Table 2 and Table 3). Consistent with the results from the calibration experiment, ginger rhizomes under 25 and 30 °C had a higher rooting index (from 3.3 to 3.5), produced 323–517% more roots per plant, had 55.1–68.4-mm longer roots, and took less time to produce 1-cm and 5-cm sprouts (up to 53 and 20 fewer days, respectively) compared with rhizomes under 20 °C (Table 2). The number and height of ginger sprouts under 30 °C were 3.3–4.9 higher and 4.1–6.9 mm taller, respectively, than those under 20, 25, and 35 °C. In ginger, the time to reach the transplant stage was similar in rhizomes under 20, 25, and 30 °C. Similar to our findings, Chen at al. [44] showed that ginger transplants from rhizomes sprouted under 25 °C produced more shoots and leaves, taller stems, and were more vigorous than those under 15 and 30 °C.
In the validation experiment, turmeric rhizomes under 25 and 30 °C had a higher rooting index (from 2.3 to 2.8), produced 5.6–6.2 more roots, and had 23.3–26.5-mm longer roots compared with those under 35 °C (Table 3). Less time to produce 1-cm sprouts (23 fewer days), 5-cm sprouts (35 fewer days), and to reach the transplant stage (19 fewer days) was measured in turmeric rhizomes under 30 °C compared with those under 20 °C. Moreover, the number and length of turmeric sprouts under 25 and 30 °C were 1.7–5.2 higher and 1.1–3.7 mm taller, respectively, than those under 14, 20, and 35 °C.
3.4. Rhizome Morphology and Sprouting
For both species, there were no differences between the varieties in the morphological measurements of seed rhizomes, regardless of the experiment (Table 4). In the calibration experiment, most of the Pearson’s correlation coefficients (r) showed weak but significant positive correlations between the morphologic parameters from rhizomes in both species (Table 4). For example, the ginger weight and length (r = 0.44), weight and width (r = 0.65), and weight and thickness (r = 0.32) were positively correlated. In turmeric, the rhizome weight and width (r = 0.62) and length and width (r = 0.42) were also positively correlated. The highest correlation was observed for the turmeric rhizome weight and length (r = 0.76), which, as expected, indicates that the longer rhizomes were heavier. Considering the large variability in rhizome morphology, predicting one or more characteristics of rhizome morphology based on others is unlikely. Based on the above, an ANCOVA was performed including all the morphological parameters as covariates. Except for temperature, no other parameter had a significant effect on rhizome sprouting, regardless of species (Table 5).
Sprouting was not affected by the morphological characteristics of the rhizomes used in our experiments (Table 5). We attribute this result to the limited number and variability of the rhizomes in our sample population. Although sprouting and transplant growth may not depend on rhizome morphology, large rhizomes have been shown to increase the final yield of ginger and turmeric [34,36]. Beale et al. [48] showed that ginger rhizomes of 86–114 g produced a larger yield compared with smaller rhizomes of 14–43 g. Similarly, Padmadevi et al. [17] reported a higher shoot biomass and yield in the field using turmeric rhizomes ranging from 45 to 50 g compared with smaller rhizomes (10–15 g). Hossain et al. [49] also showed higher yield of turmeric plants grown from seed rhizomes that weighed 30–40 g compared with those weighing 10 and 20 g. Based on this, rhizome morphology should be considered when growing these crops for commercial production, as they are likely to affect the yield.
4. Conclusions
Temperature is a key factor in the sprouting of ginger and turmeric rhizomes. Based on our model, the optimum temperatures to induce sprouting are 27.5 °C in ginger and 30.1 °C in turmeric. The Tₘᵢₙ and Tₘₐₓ to obtain 5-cm sprouts were 17.1 and 35.2 °C in ginger and 17.2 and 32.8 °C in turmeric, respectively. The shortest times to sprout were 29 and 31 d in ginger and turmeric, respectively. Temperatures below 14 °C or above 40 °C are likely to inhibit sprouting or lead to rhizome loss.
The results from this study showed the potential to produce uniform ginger and turmeric transplants using temperature treatments that accelerate sprouting. By pre-sprouting seed rhizomes, growers could extend the production cycle of these crops, as the time in the field from seed placement to the start of sprouting would be eliminated. This is advantageous, particularly in cold regions that would benefit from early spring planting. Similarly, pre-sprouting in hot regions could prove to be advantageous, as it would enable better crop establishment before high summer temperatures begin to negatively affect plant growth in the field.
Author Contributions
Conceptualization, methodology, and investigation, M.R.-C., P.R.F. and C.G.; formal analysis, data curation, visualization, and writing—original draft, M.R.-C.; writing—review and editing, M.R.-C., P.R.F. and C.G.; and funding acquisition, P.R.F. and C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly funded by the USDA-ARS Floriculture and Nursery Research Initiative 58-5082-8-012 “Resilient Plants”, USDA National Institute of Food and Agriculture, Multistate Research Project NE1835: Resource Optimization in Controlled Environment Agriculture, the Field and Fork Farm at the University of Florida, and industry partners in the Floriculture Research Alliance (FloricultureAlliance.org).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Hawaii Clean Seed, LLC. and Just Ginger Florida for providing the rhizomes. We also thank Maria Paz Kinslow for the experimental and technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nguyen, L.; Duong, L.T.; Mentreddy, R.S. The, U.S. import demand for spices and herbs by differentiated sources. J. Appl. Res. Med. Aromat. Plants 2019, 12, 13–20. [Google Scholar] [CrossRef]
- Tridge. Available online: https://www.tridge.com/intelligences/ginger (accessed on 2 July 2021).
- Tridge. Available online: https://www.tridge.com/intelligences/turmeric1 (accessed on 2 July 2021).
- Ma, R.H.; Ni, Z.J.; Zhu, Y.Y.; Thakur, K.; Zhang, F.; Zhang, Y.Y.; Hu, F.; Zhang, J.G.; Wei, Z.J. A recent update on the multifaceted health benefits associated with ginger and its bioactive components. Food Funct. 2021, 12, 519–542. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, W.; Deng, G.; Wang, P.; Yang, P.; Aggarwal, B. Chemical composition and product quality control of turmeric (Curcuma longa L.). Pharm. Crops 2011, 2, 28–54. [Google Scholar] [CrossRef]
- Ali, B.H.; Blunden, G.; Tanira, M.O.; Nemmar, A. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol. 2008, 46, 409–420. [Google Scholar] [CrossRef] [PubMed]
- El Kutry, M.S. Potential protection effect of using honey, ginger, and turmeric as a natural treatment against chemotherapy of intestinal toxicity. J. Biol. Act. Prod. Nat. 2020, 10, 86–99. [Google Scholar] [CrossRef]
- Nair, A.; Amalraj, A.; Jacob, J.; Kunnunmakkara, A.; Gopi, S. Non-Curcuminoids from Turmeric and Their Potential in Cancer Therapy and Anticancer Drug Delivery Formulations. Biomolecules 2019, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, M.D.; Blázquez, M.A. Ginger and turmeric essential oils for weed control and food crop protection. Plants 2019, 8, 59. [Google Scholar] [CrossRef] [Green Version]
- Shannon, D.A.; van Santen, E.; Salmasi, S.Z.; Murray, T.J.; Duong, L.T.; Greenfield, J.T.; Gonzales, T.; Foshee, W. Shade, establishment method, and varietal effects on rhizome yield and curcumin content in turmeric in Alabama. Crop Sci. 2019, 59, 2701–2710. [Google Scholar] [CrossRef]
- Govindasamy, R.; Nemana, A.; Puduri, V.; Pappas, K. Ethnic produce marketing in the Mid-Atlantic states: Consumer shopping patterns and willingness-to-pay analysis. Choices 2006, 21, 237–241. [Google Scholar]
- Sciarappa, W.J.; Simon, J.; Govindasamy, R.; Kelley, K.; Mangan, F.; Zhang, S.; Arumugam, S.; Nitzsche, P.; Van Vranken, R.; Komar, S.; et al. Asian crops overview: Consumer preference and cultivar growth on the east coast of the United States. HortScience 2016, 51, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Flores, S.; Retana-Cordero, M.; Fisher, P.R.; Freyre, R.; Gómez, C. Effect of photoperiod, propagative material, and production period on greenhouse-grown ginger and turmeric plants. HortScience. in press.
- Abelenda, J.A.; Prat, S. Cytokinins: Determinants of sink storage ability. Curr. Biol. 2013, 23, 561–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, P.N.; Nirmal Babu, K.; Shiva, K.N. Botany and Crop Improvement of Ginger. In Ginger: The Genus Zingiber, 1st ed.; Ravindran, P.N., Nirmal Babu, K., Eds.; CRC Press: Boca Raton, FL, USA, 2005; Volume 1, pp. 15–86. [Google Scholar]
- Ravindran, P.N.; Nirmal Babu, K.; Shiva, K.N. Botany and Crop Improvement of Turmeric. In Turmeric: The Genus Curcuma, 1st ed.; Ravindran, P.N., Nirmal Babu, K., Sivaraman, K., Eds.; CRC Press: Boca Raton, FL, USA, 2007; Volume 1, pp. 15–70. [Google Scholar]
- Padmadevi, K.; Jothi, L.J.; Ponnuswami, V.; Durgavathi, V.; Parveen, I.R. Effect of different grades of rhizomes on growth and yield of turmeric (Curcuma longa L.). Asian J. Hortic. 2012, 7, 465–467. [Google Scholar]
- Sharma, K.; Rok Lee, Y.; Park, S.W.; Nile, S.H. Importance of growth hormones and temperature for physiological regulation of dormancy and sprouting in onions. Food Rev. Int. 2016, 32, 233–255. [Google Scholar] [CrossRef]
- Aswathy, T.S.; Jessykutty, P.S. Effect of growth regulators and chemicals on sprouting of minisetts in Curcuma aromatica Salisb. Int. J. Appl. Pure Sci. Agr. 2016, 2, 8–11. [Google Scholar]
- Criley, R.A. Propagation of cut flowers: Strelitzia, Alpinia, and Heliconia. Acta Hortic. 1988, 226, 509–518. [Google Scholar] [CrossRef]
- Esashi, Y.; Leopold, A.C. Dormancy regulation in subterranean clover seeds by ethylene. Plant Physiol. 1969, 44, 1470–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thohirah, L.A.; Flora, C.L.S.; Kamalakshi, N. Breaking bud dormancy and different shade levels for production of pot and cut Curcuma alismatifolia. Am. J. Agr. Biol. Sci. 2010, 5, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Furutani, S.C.; Villanueva, J.; Tanable, M.J. Effect of ethephon and heat on the growth and yield of edible ginger. HortScience 1985, 20, 392–393. [Google Scholar]
- Miedema, P. Bulb dormancy in onion. I. The effects of temperature and cultivar on sprouting and rooting. J. Hortic. Sci. 1994, 69, 29–39. [Google Scholar] [CrossRef]
- Senning, M.; Sonnewald, U.; Sonnewald, S. Deoxyuridine triphosphatase expression defines the transition from dormant to sprouting potato tuber buds. Mol. Breed. 2010, 26, 525–531. [Google Scholar] [CrossRef]
- Kaushal, M.; Gupta, A.; Vaidya, D.; Gupta, M. Postharvest management and value addition of ginger (Zingiber officinale Roscoe): A review. Int. J. Environ. Agric. Biotechnol. 2017, 2, 397–412. [Google Scholar] [CrossRef]
- Paull, R.E.; Chen, N.J.; Goo, T.T.C. Control of weight loss and sprouting of ginger rhizomes in stores. HortScience 1988, 23, 734–736. [Google Scholar]
- Lee, J.H.; Oh, M.W.; Jang, H.D.; Lee, Y.J.; Jeong, J.T.; Park, C.G. Growth characteristics of turmeric (Curcuma longa L.) germplasms and storage conditions of seed rhizomes. Korean J. Med. Crop Sci. 2020, 28, 47–55. [Google Scholar] [CrossRef]
- De Melo-Abreu, J.P.; Barranco, D.; Cordeiro, A.M.; Tous, J.; Rogado, B.M.; Villalobos, F.J. Modelling olive flowering date using chilling for dormancy release and thermal time. Agric. Meteorol. 2004, 125, 117–127. [Google Scholar] [CrossRef]
- Taab, A.; Andersson, L.; Boström, U. Modelling the sprouting capacity from underground buds of the perennial weed. Sonchus Arvensis Weed Res. 2018, 58, 348–356. [Google Scholar] [CrossRef]
- Blanchard, M.G.; Runkle, E.S.; Fisher, P.R. Modeling plant morphology and development of petunia in response to temperature and photosynthetic daily light integral. Sci. Hortic. 2011, 129, 313–320. [Google Scholar] [CrossRef]
- Norouzi, H.A.; Vazin, F. Modelling of the Faba bean (Vicia faba L.) Sprouting Reaction to Temperature in Farm Condition. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Walters, K.J.; Lopez, R.G. Modeling growth and development of hydroponically grown dill, parsley, and watercress in response to photosynthetic daily light integral and mean daily temperature. PLoS ONE 2021, 16, e0248662. [Google Scholar] [CrossRef]
- Girma, H.; Kindie, T. The effects of seed rhizome size on the growth, yield and economic return of ginger (Zingiber officinale Rose.). Asian J. Plant Sci. 2008, 7, 213–217. [Google Scholar]
- Hossain, M. Effects of harvest time on shoot biomass and yield of turmeric (Curcuma longa L.) in Okinawa, Japan. Plant Prod. Sci. 2010, 13, 97–103. [Google Scholar] [CrossRef]
- Kadam, J.H.; Waghmode, R.S.; Kathmale, D.K.; Ranpise, S.A. Effect of seed rhizome cuttings on growth and yield of turmeric. In Advances in Planting Material Production Technology in Spices, Proceedings of National Seminar on Planting Material Production in Spices: Kozhikode, Kerala, India, 21–22 April 2016; Malhotra, S.K., Kandiannan, K., Mini Raj, K., Neema, V.P., Prasath, D., Srinivasan, V., Homey Cheriyan, F., Eds.; Directorate of Arecanut and Spices Development: Kozhikode, India, 2016; pp. 145–149. [Google Scholar]
- Nybe, E.V.; Raj, N.M. Ginger production in India and other South Asian countries. In Ginger: The Genus Zingiber, 1st ed.; Ravindran, P.N., Nirmal Babu, K., Eds.; CRC Press: Boca Raton, FL, USA, 2005; Volume 1, pp. 211–240. [Google Scholar]
- Brøndum, J.J.; Heins, R.D. Modeling temperature and photoperiod effects on growth and development of dahlia. J. Am. Soc. Hortic. Sci. 1993, 118, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, J.J. Some useful equations for biological studies. Expt. Agr. 1977, 13, 273–286. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project (accessed on 10 April 2021).
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-3. 2020. Available online: https://CRAN.R-project:package=agricolae (accessed on 12 April 2021).
- Policegoudra, R.S.; Aradhya, S.M.; Singh, L. Mango ginger (Curcuma amada Roxb.)—A promising spice for phytochemicals and biological activities. J. Biosci. 2011, 36, 739–748. [Google Scholar] [CrossRef]
- Ishimine, Y.; Hossain, M.A.; Motomura, K.; Akamine, H.; Hirayama, T. Effects of planting date on emergence, growth and yield of turmeric (Curcuma longa L.) in Okinawa Prefecture, Southern Japan. Jpn. J. Trop. Agric. 2004, 48, 10–16. [Google Scholar] [CrossRef]
- Chen, W.; Li, R.; Tsai, Z.; Hsiao, C. Study on improving sprout forcing methods for early ginger (Zingiber officinale) production. J. Taiwan Soc. Hortic. Sci. 2014, 60, 253–264. [Google Scholar]
- Dourado, C.; Pinto, C.; Barba, F.J.; Lorenzo, J.M.; Delgadillo, I.; Saraiva, J.A. Innovative non-thermal technologies affecting potato tuber and fried potato quality. Trends Food Sci. Technol. 2019, 88, 274–289. [Google Scholar] [CrossRef]
- Grevsen, K.; Sorensen, J.N. Sprouting and yield in bulb onions (Allium cepa L.) as influenced by cultivar, plant establishment methods, maturity at harvest and storage conditions. J. Hortic. Sci. Biotechnol. 2004, 79, 877–884. [Google Scholar] [CrossRef]
- Sonnewald, S.; Sonnewald, U. Regulation of potato tuber sprouting. Planta 2014, 239, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Beale, A.J.; Ramírez, M.; Díaz, M.; Muñoz, A.; Flores, C. Effect of seed sett weight of ginger (Zingiber officinale) on yield. In Food Safety and Value Added Production and Marketing in Tropical Crops, Proceedings of the Caribbean Food Crops Society 42th Annual Meeting, Carolina, Puerto Rico, 9–15 July 2006; Santiago, H.L., Lugo, W.I., Eds.; Caribbean Food Crops Society: Carolina, Puerto Rico, 2006; pp. 407–411. [Google Scholar] [CrossRef]
- Hossain, A.; Ishimine, Y.; Akamine, H.; Motomura, K. Effects of seed rhizome size on growth and yield of turmeric (Curcuma longa L.). Plant Prod. Sci. 2005, 8, 86–94. [Google Scholar] [CrossRef]
Figure 1.
Calibration of ginger and turmeric data to the Landsberg function used in Equation (1) based on the parameters from Table 1. Symbols represent the mean ± 95% confidence intervals. Data were pooled across two cultivars and include 12 rhizomes per species per temperature treatment.
Figure 1.
Calibration of ginger and turmeric data to the Landsberg function used in Equation (1) based on the parameters from Table 1. Symbols represent the mean ± 95% confidence intervals. Data were pooled across two cultivars and include 12 rhizomes per species per temperature treatment.

Figure 2.
Validation of ginger and turmeric data to the Landsberg function used in Equation (1) based on the parameters from Table 2. Symbols represent the mean ± 95% confidence intervals. Data were pooled across two cultivars and include 12 rhizomes per species per temperature treatment.
Figure 2.
Validation of ginger and turmeric data to the Landsberg function used in Equation (1) based on the parameters from Table 2. Symbols represent the mean ± 95% confidence intervals. Data were pooled across two cultivars and include 12 rhizomes per species per temperature treatment.

Table 1.
Nonlinear regression results from fitting the Landsberg function (Equation (1)) to the inverse of days to 5-cm sprouting for the calibration dataset.
Table 1.
Nonlinear regression results from fitting the Landsberg function (Equation (1)) to the inverse of days to 5-cm sprouting for the calibration dataset.
| Asymptotic 95% Confidence Interval | |||||
| Species | Parameter z | Estimate | SE | Lower | Upper |
| Ginger | Tₘᵢₙ | 17.1 | 0.535 | 16.0 | 18.1 |
| Tₒₚₜ | 27.5 | 0.385 | 26.7 | 28.3 | |
| Tₘₐₓ | 35.2 | 1.387 | 32.5 | 38.0 | |
| Rₘₐₓ | 0.0342 | 0.000535 | 0.0331 | 0.0353 | |
| Turmeric | Tₘᵢₙ | 17.2 | 0.826 | 15.5 | 18.8 |
| Tₒₚₜ | 30.1 | 0.636 | 28.8 | 31.4 | |
| Tₘₐₓ | 32.8 | 0.676 | 31.5 | 34.2 | |
| Rₘₐₓ | 0.0318 | 0.00134 | 0.0291 | 0.0344 | |
z Tₘᵢₙ = Minimum temperature (°C) for sprouting, Topt = Optimum temperature (°C) for sprouting, and Tₘₐₓ = Maximum temperature (°C) for sprouting.
Table 3.
Seed rhizome growth parameters measured from “Indira Yellow” and “Hawaiian Red” turmeric kept under different temperatures inside incubators and then grown in a greenhouse in two experimental phases.
Table 3.
Seed rhizome growth parameters measured from “Indira Yellow” and “Hawaiian Red” turmeric kept under different temperatures inside incubators and then grown in a greenhouse in two experimental phases.
| Experiment | Sprouts per Plant (No.) | Average Sprout Height (mm) | Roots per Plant (No.) | Primary Root Length (mm) | Rooting Index (1 to 5) | Days to 1-cm Sprout | Days to 5-cm Sprout | Transplant Stage in Greenhouse (days) |
|---|---|---|---|---|---|---|---|---|
| Calibration | ||||||||
| 21 °C | 5.1 | 3.1 c z | - | - | - | 57.9 a | 81.9 a | 29.2 a |
| 25 °C | 6.0 | 4.4 b | 3.0 ab | 19.8 c | 2.1 a | 32.7 b | 51.6 c | 23.4 b |
| 27 °C | 5.3 | 5.8 a | 4.5 a | 28.1 b | 3.2 a | 20.3 c | 31.2 c | 18.2 b |
| 30 °C | 4.8 | 6.7 a | 4.6 a | 37.9 a | 2.9 a | 21.0 c | 32.3 c | 20.5 b |
| 32 °C | 5.5 | 5.2 a | 2.7 b | 31.3 a | 1.4 b | 24.3 c | 38.1 b | 32.7 a |
| Treatment (T) | NS | *** | *** | *** | ** | *** | *** | ** |
| Variety (V) | NS | NS | NS | NS | NS | NS | NS | NS |
| T × V | NS | NS | NS | NS | NS | NS | NS | NS |
| Validation | ||||||||
| 14 °C | 0.3 c | 0.1 c | - | - | - | - | - | - |
| 20 °C | 4.6 a | 2.2 b | - | - | - | 62.2 a | 87.0 a | 24.6 a |
| 25 °C | 5.5 a | 3.8 a | 6.5 a | 27.7 a | 2.3 a | 35.3 b | 52.0 b | 22.7 a |
| 30 °C | 4.0 a | 3.7 a | 5.9 a | 30.9 a | 2.8 a | 22.6 c | 34.5 c | 19.2 b |
| 35 °C | 3.3 b | 2.6 b | 0.3 b | 4.4 b | 0.5 b | 30.1 b | 55.4 b | - |
| 40 °C | - | - | - | - | - | - | - | - |
| T | ** | *** | *** | ** | *** | *** | *** | ** |
| V | NS | NS | NS | NS | NS | NS | NS | NS |
| T × V | NS | NS | NS | NS | NS | NS | NS | NS |
z Means within columns followed by the same letter are not different based on Tukey’s honestly significant different test at p ≤ 0.05 (n = 12). ***, **, and NS indicate statistical significance at the 0.001 and 0.01 p ≤ levels or not significant, respectively. - indicates no data.
Table 4.
Pearson correlation of the morphological parameters of ginger and turmeric seed rhizomes.
| Parameter | Weight | Length | Width | Thickness |
|---|---|---|---|---|
| Ginger | ||||
| Weight | 1 | |||
| Length | 0.44 *** | 1 | ||
| Width | 0.65 *** | 0.19 * | 1 | |
| Thickness | 0.32 ** | 0.04 | 0.19 * | 1 |
| Turmeric | ||||
| Weight | 1 | |||
| Length | 0.76 *** | 1 | ||
| Width | 0.62 *** | 0.42 *** | 1 | |
| Thickness | 0.08 | 0.04 | 0.07 | 1 |
***, **, *, and NS indicate statistical significance at the 0.001, 0.01, and 0.05 p ≤ levels or not significant, respectively.
Table 5.
Analysis of covariance on the effect of the temperature and morphological parameters on the sprouting of ginger and turmeric seed rhizomes from the calibration experiment.
Table 5.
Analysis of covariance on the effect of the temperature and morphological parameters on the sprouting of ginger and turmeric seed rhizomes from the calibration experiment.
| p-Value | Reg. Coef. | |
|---|---|---|
| Ginger | ||
| Model | <0.0001 | |
| Treatment | ||
| Temperature | <0.0001 | |
| Covariates | ||
| Weight | 0.5018 | −0.09 |
| Length | 0.2410 | 0.08 |
| Width | 0.6085 | −0.04 |
| Thickness | 0.2029 | −0.003 |
| Turmeric | ||
| Model | <0.0001 | |
| Treatment | ||
| Temperature | <0.0001 | |
| Covariates | ||
| Weight | 0.4151 | −0.1 |
| Length | 0.2744 | 0.09 |
| Width | 0.6642 | −0.02 |
| Thickness | 0.4427 | −0.004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Retana-Cordero, M.; Fisher, P.R.; Gómez, C. Modeling the Effect of Temperature on Ginger and Turmeric Rhizome Sprouting. Agronomy 2021, 11, 1931. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101931
AMA Style
Retana-Cordero M, Fisher PR, Gómez C. Modeling the Effect of Temperature on Ginger and Turmeric Rhizome Sprouting. Agronomy. 2021; 11(10):1931. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101931
Chicago/Turabian StyleRetana-Cordero, Marlon, Paul R. Fisher, and Celina Gómez. 2021. "Modeling the Effect of Temperature on Ginger and Turmeric Rhizome Sprouting" Agronomy 11, no. 10: 1931. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11101931
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.