
Quantitative Trait Loci for Resistance to Potato Dry Rot Caused by Fusarium sambucinum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fusarium sambucinum
2.3. Dry Rot Resistance Tests
2.4. Genetic Maps and QTL Analysis
2.5. Statistical Analyses
3. Results
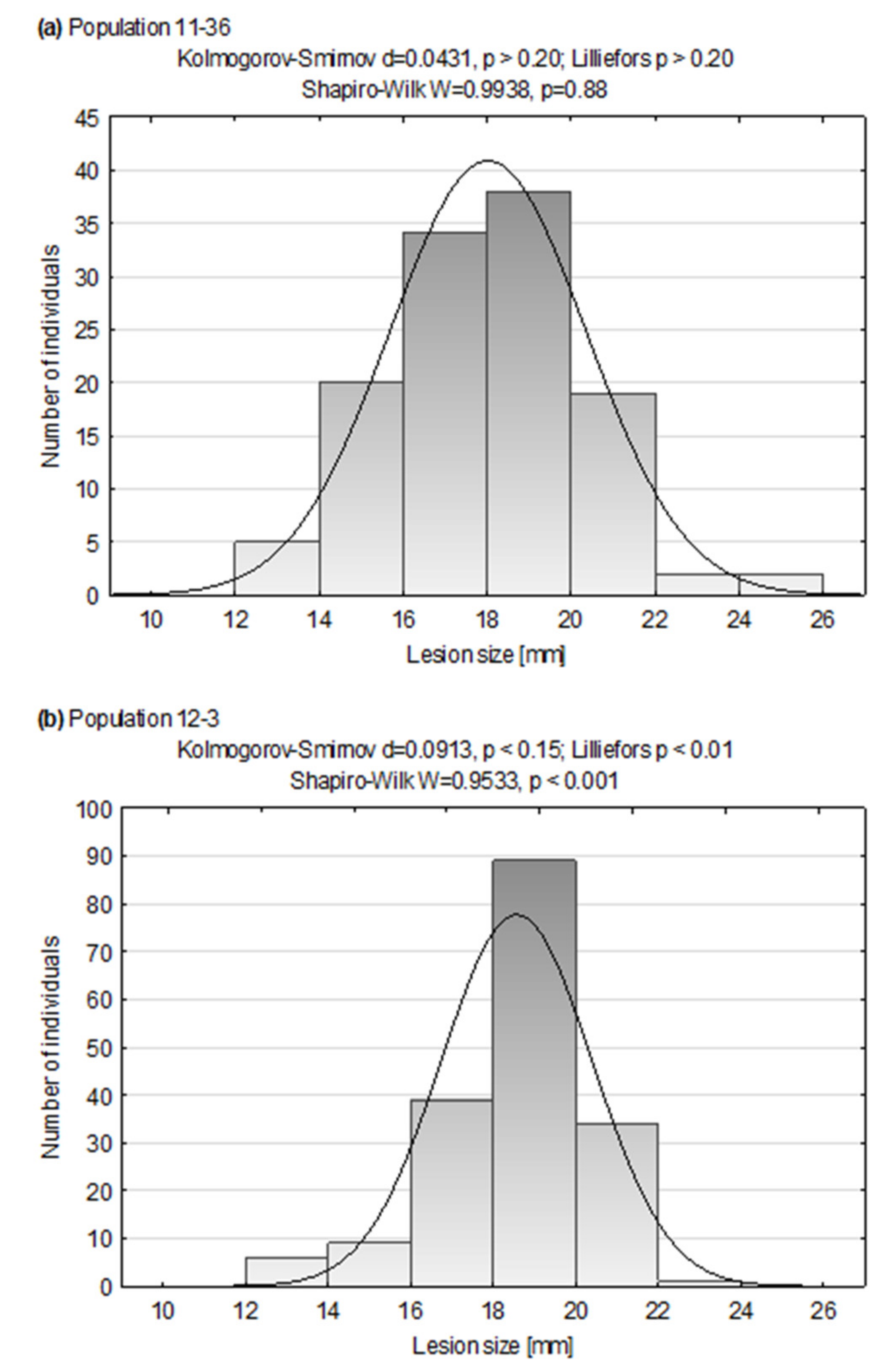
3.1. Dry Rot Resistance Tests
3.2. QTL Analysis in Population 11–36
3.3. QTL Analysis in Population 12–3
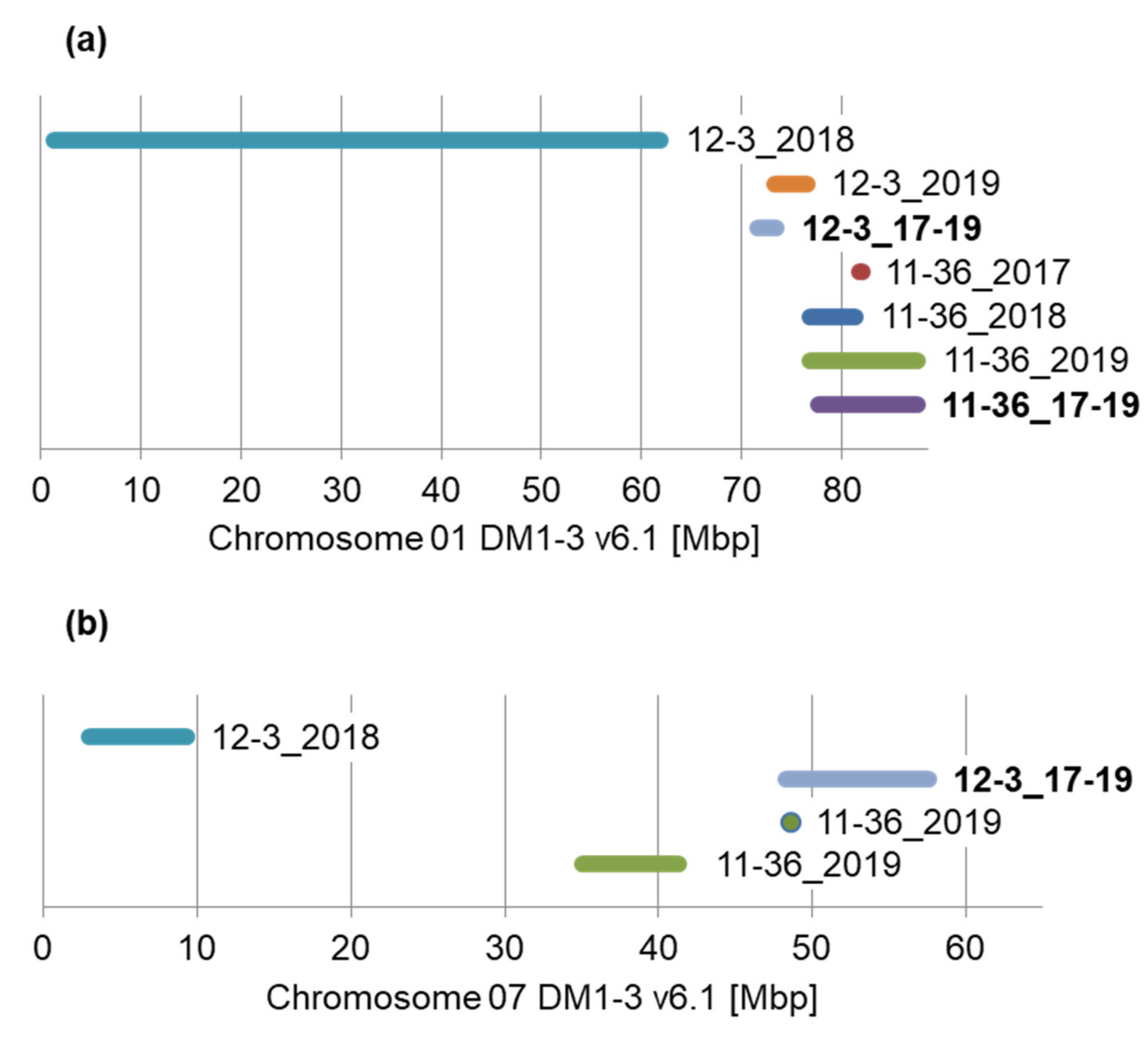
3.4. QTL Location in the Reference Genome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato dry rot disease: Current status, pathogenomics and management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Cullen, D.W.; Toth, I.K.; Pitkin, Y.; Boonham, N.; Walsh, K.; Barker, I.; Lees, A.K. Use of quantitative molecular diagnostic assays to investigate Fusarium dry rot in potato stocks and soil. Phytopathology 2005, 95, 1462–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Denis, M.; Crous, P.W. Removing chaos from confusion: Assigning names to common human and animal pathogens in Neocosmospora. Persoonia 2018, 41, 109–129. [Google Scholar] [CrossRef] [PubMed]
- Stefańczyk, E.; Sobkowiak, S.; Brylińska, M.; Śliwka, J. Diversity of Fusarium spp. associated with dry rot of potato tubers in Poland. Eur. J. Plant Pathol. 2016, 145, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Azil, N.; Stefańczyk, E.; Sobkowiak, S.; Chihat, S.; Boureghda, H.; Śliwka, J. Identification and pathogenicity of Fusarium spp. associated with tuber dry rot and wilt of potato in Algeria. Eur. J. Plant Pathol. 2021, 159, 495–509. [Google Scholar] [CrossRef]
- Ramlawi, S.; Chiu, J.; Cloutier, A.; Avis, T. Suppression of Fusarium dry rot of potato using beneficial bacterial treatments. J. Plant Pathol. 2021, 103, 269–281. [Google Scholar] [CrossRef]
- Vatankhah, M.; Riseh, R.; Eskandari, M.; Sedaghati, E.; Alaei, H.; Afzali, H. Biological control of Fusarium dry rot of potato using some probiotic bacteria. J. Agric. Sci. Technol. 2019, 21, 1301–1312. [Google Scholar]
- Raigond, P.; Sagar, V.; Mishra, T.; Thakur, A.; Singh, B.; Kumar, V.; Gupta, V.; Dutt, S.; Changan, S. Chitosan: A safe alternative to synthetic fungicides to manage dry rot in stored potatoes. Potato Res. 2019, 62, 393–409. [Google Scholar] [CrossRef]
- Bojanowski, A.; Avis, T.J.; Pelletier, S.; Tweddell, R.J. Management of potato dry rot. Postharvest Biol. Technol. 2013, 84, 99–109. [Google Scholar] [CrossRef]
- Wojciechowska-Kot, H. Susceptibility of potato varieties to Fusarium decay. Biul. Inst. Ziemn. 1975, 15, 97–109. [Google Scholar]
- Pett, B.; Götz, J.; Granzow, H. Untersuchungen zum Sortenverhalten der Kartoffel gegen Fusarium—Trockenfäule. Arch. Zucht. Berl. 1977, 7, 85–93. [Google Scholar]
- Jellis, G.; Starling, N. Resistance to powdery dry rot (Fusarium sulphureum) in potato tubers. Potato Res. 1983, 26, 295–301. [Google Scholar] [CrossRef]
- Du, M.; Ren, X.; Sun, Q.; Wang, Y.; Zhang, R. Characterization of Fusarium spp. causing potato dry rot in China and susceptibility evaluation of Chinese potato germplasm to the pathogen. Potato Res. 2012, 55, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Heltoft, P.; Molteberg, E.L.; Naerstad, R.; Hermansen, A. Effect of maturity level and potato cultivar on development of Fusarium dry rot in Norway. Potato Res. 2015, 58, 205–219. [Google Scholar] [CrossRef]
- Aydin, M.; Inal, B. Comparative susceptibility of some commercial potato cultivars to Fusarium sambucinum and F. solani isolates causing tuber dry rot. Appl. Ecol. Environ. Res. 2018, 16, 4879–4892. [Google Scholar] [CrossRef]
- Wojciechowska-Kot, H. Methods of investigating the susceptibility of potato tubers to Fusarium sulphureum Schl. and F. solani var. coeruleum (Lib. Sacc.) Booth. Zesz. Nauk. Akad. Rol. Tech. Olszt. 1981, 31, 41–48. [Google Scholar]
- Lees, A.; Bradshaw, J.; Stewart, H. Inheritance of resistance to Fusarium spp. and to Phytophthora infestans in crosses between Neotuberosum and Tuberosum potatoes estimated by seedling tests. Potato Res. 1998, 41, 267–275. [Google Scholar] [CrossRef]
- Lynch, D.; Kawchuk, L.; Chen, Q.; Kokko, M. Resistance to Fusarium sambucinum in wild and cultivated Solanum species. Am. J. Potato Res. 2003, 80, 353–358. [Google Scholar] [CrossRef]
- Corsini, D.; Pavek, J. Fusarium dry-rot resistant potato germplasm. Am. Potato J. 1986, 63, 629–638. [Google Scholar] [CrossRef]
- Valluru, R.; Christ, B.; Haynes, K.; Vinyard, B. Inheritance and stability of resistance to Fusarium tuber rot in tetraploid potatoes. Am. J. Potato Res. 2006, 83, 335–341. [Google Scholar] [CrossRef]
- Burkhart, C.; Christ, B.; Haynes, K. Non-additive genetic variance governs resistance to Fusarium dry rot in a diploid hybrid potato population. Am. J. Potato Res. 2007, 84, 199–204. [Google Scholar] [CrossRef]
- Hara-Skrzypiec, A.; Śliwka, J.; Jakuczun, H.; Zimnoch-Guzowska, E. QTL for tuber morphology traits in diploid potato. J. Appl. Genet. 2018, 59, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Skrzypiec, A.; Śliwka, J.; Jakuczun, H.; Zimnoch-Guzowska, E. Quantitative trait loci for tuber blackspot bruise and enzymatic discoloration susceptibility in diploid potato. Mol. Genet. Genom. 2018, 293, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Śliwka, J.; Sołtys-Kalina, D.; Szajko, K.; Wasilewicz-Flis, I.; Strzelczyk-Zyta, D.; Zimnoch-Guzowska, E.; Jakuczun, H.; Marczewski, W. Mapping of quantitative trait loci for tuber starch and leaf sucrose contents in diploid potato. Theor. Appl. Genet. 2016, 129, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Sołtys-Kalina, D.; Szajko, K.; Stefańczyk, E.; Smyda-Dajmund, P.; Śliwka, J.; Marczewski, W. eQTL mapping of the 12S globulin cruciferin gene PGCRURSE5 as a novel candidate associated with starch content in potato tubers. Sci. Rep. 2020, 10, 17168. [Google Scholar] [CrossRef] [PubMed]
- Sołtys-Kalina, D.; Szajko, K.; Wasilewicz-Flis, I.; Mańkowski, D.; Marczewski, W.; Śliwka, J. Quantitative trait loci for starch-corrected chip color after harvest, cold storage and after reconditioning mapped in diploid potato. Mol. Genet. Genom. 2020, 295, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Stefańczyk, E.; Sobkowiak, S. Isolation, identification and preservation of Fusarium spp. causing dry rot of potato tubers. Plant Breed. Seed Sci. 2017, 76, 45–51. [Google Scholar] [CrossRef]
- Stefańczyk, E. Assessment of potato tuber resistance against dry rot. Plant Breed. Seed Sci. 2017, 76, 53–56. [Google Scholar] [CrossRef]
- Van Oojien, J.W. JoinMap®4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma B.V.: Wageningen, The Netherlands, 2006. [Google Scholar]
- Van Ooijen, J.W. MapQTL ® 6. Software for Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; Kyazma B.V.: Wageningen, The Netherlands, 2009. [Google Scholar]
- Van Ooijen, J. LOD significance thresholds for QTL analysis in experimental populations of diploid species. Heredity 1999, 83, 613–624. [Google Scholar] [CrossRef] [Green Version]
- Domański, L.; Michalak, K.; Zimnoch-Guzowska, E. Variation of blackspot susceptibility of the selected potato cultivars. Biul. Inst. Hod. Aklim. Roślin 2007, 246, 145–149. [Google Scholar]
- Pham, G.M.; Hamilton, J.P.; Wood, J.C.; Burke, J.T.; Zhao, H.N.; Vaillancourt, B.; Ou, S.J.; Jiang, J.M.; Buell, C.R. Construction of a chromosome-scale long-read reference genome assembly for potato. Gigascience 2020, 9, giaa100. [Google Scholar] [CrossRef]
- Ayers, G.W.; Robinson, D.B. An inoculation technique for the study of dry rot of the potatoes. Am. Potato J. 1954, 31, 278–281. [Google Scholar] [CrossRef]
- Boyd, A.E.W. Dry-rot disease of the potato. 4. Laboratory methods used in assessing variations in tuber susceptibility. Ann. Appl. Biol. 1952, 39, 322–329. [Google Scholar] [CrossRef]
- Prodhomme, C.; van Arkel, G.; Plich, J.; Tammes, J.; Rijk, J.; van Eck, H.; Visser, R.; Vossen, J. A Hitchhiker’s guide to the potato wart disease resistance galaxy. Theor. Appl. Genet. 2020, 133, 3419–3439. [Google Scholar] [CrossRef] [PubMed]
- Simko, I.; Costanzo, S.; Haynes, K.; Christ, B.; Jones, R. Linkage disequilibrium mapping of a Verticillium dahliae resistance quantitative trait locus in tetraploid potato (Solanum tuberosum) through a candidate gene approach (vol 108, pg 217, 2004). Theor. Appl. Genet. 2004, 108, 1191. [Google Scholar] [CrossRef] [Green Version]
- Uribe, P.; Jansky, S.; Halterman, D. Two CAPS markers predict Verticillium wilt resistance in wild Solanum species. Mol. Breed. 2014, 33, 465–476. [Google Scholar] [CrossRef]
- Lynch, D.; Kawchuk, L.; Hachey, J.; Bains, P.; Howard, R. Identification of a gene conferring high levels of resistance to Verticillium wilt in Solanum chacoense. Plant Dis. 1997, 81, 1011–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simko, I.; Haynes, K.; Ewing, E.; Costanzo, S.; Christ, B.; Jones, R. Mapping genes for resistance to Verticillium albo-atrum in tetraploid and diploid potato populations using haplotype association tests and genetic linkage analysis. Mol. Genet. Genom. 2004, 271, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Massa, A.; Manrique-Carpintero, N.; Coombs, J.; Haynes, K.; Bethke, P.; Brandt, T.; Gupta, S.; Yencho, G.; Novy, R.; Douches, D. Linkage analysis and QTL mapping in a tetraploid russet mapping population of potato. BMC Genet. 2018, 19, 87. [Google Scholar] [CrossRef] [Green Version]
- Odilbekov, F.; Selga, C.; Ortiz, R.; Chawade, A.; Liljeroth, E. QTL Mapping for resistance to early blight in a tetraploid potato population. Agronomy 2020, 10, 728. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plant Material | Mean Lesion Size in mm (±SD) |
|---|---|
| Standard cultivars | |
| cv. Bartek | 14.1 (±5.1) aa |
| cv. Kuba | 18.2 (±8.1) b |
| cv. Harpun | 20.3 (±5.3) c |
| cv. Gawin | 20.4 (±6.6) c |
| cv. Hinga | 24.8 (±8.5) d |
| Parental forms | |
| DG 06-5 | 12.2 (±3.8) a |
| DG 08-28/13 | 19.9 (±4.9) bc |
| DG 00-683 | 21.4 (±4.5) c |
| DG 03-226 | 23.7 (±7.2) d |
| Population 11–36 | |
| Population mean | 18.0 (±2.3) |
| Population median | 18.0 |
| Mean lesion size range | 12.9–24.7 |
| Range of Pearson’s coefficients between results from years 2017–2019 b | 0.55–0.67 |
| Population 12–3 | |
| Population mean | 18.6 (±1.8) |
| Population median | 18.8 |
| Mean lesion size range | 12.7–22.2 |
| Range of Pearson’s coefficients between results from years 2017–2019 | 0.05–0.37 |
| Factor | ANOVA | |||||
|---|---|---|---|---|---|---|
| SS | df | MS | F Value | p | Hb | |
| (a) 11–36 | ||||||
| {1} genotype | 3566.9 | 112 | 31.8 | 5.42 | <0.001 | 0.58 |
| {2} year | 15357.1 | 2 | 7678.5 | 1306.97 | <0.001 | |
| {1} × {2} | 1400.2 | 224 | 6.3 | 1.06 | 0.302 | |
| Error | 1991.6 | 339 | 5.9 | |||
| (b) 12–3 | ||||||
| {1} genotype | 1343.2 | 107 | 12.6 | 2.26 | <0.001 | 0.22 |
| {2} year | 6210.4 | 2 | 3105.2 | 557.95 | <0.001 | |
| {1} × {2} | 1445.6 | 214 | 6.8 | 1.21 | 0.058 | |
| Error | 1803.2 | 324 | 5.6 | |||
| Trait and Dataset | Chromosome | QTL Peak (cM) | Marker or Marker Interval at Peak | LOD | R2 (%) | Marker Origin a | Peak Position DM1-3 v6.1 b | QTL Range (cM) | |
|---|---|---|---|---|---|---|---|---|---|
| Chromosome | bp | ||||||||
| Population 11–36 | |||||||||
| 2017 | I | 74.9 | pPt-537438 | 3.65 | 14.3 | P2 | 01 | 84651175–84651556 | 69.4–81.5 |
| 2018 | I | 66.8 | pPt-533988 | 3.66 | 13.3 | P2 | 01 | 78163480–78164064 | 50.7–71.5 |
| 2019 | I | 71.274 | toPt-440651 | 6.05 | 20.9 | P2 | 01 | 81329689–81330960 | 50.7–108.8 |
| 17–19 | I | 81.5–84.5 | pPt-535624–pPt-652982 | 4.89 | 17.1 | P1–P2 | 01 | 87666433–87667054 | 54.1–108.1 |
| 2019 | VII | 21.1 | pPt-471208 | 3.60 | 13.0 | H | 07 | 3733794–3734717 | 20.8–21.4 |
| 2019 | VII | 38.6 | pPt-457469 | 3.28 | 11.9 | H | 07 | 48569422–48569808 | 37.3–38.6 |
| 2019 | IX | 42.6 | pPt-472930 | 5.20 | 18.2 | P1 | 09 | 59439764–59440711 | 27.3–49.0 |
| 2017 | XI | 37.6 | pPt-456657 | 3.42 | 13.5 | H | 11 | 39659759–39660130 | 37.6–38.3 |
| Population 12–3 | |||||||||
| 2018 | I | 19.5 | pPt-539549 | 3.5 | 9.0 | P2 | 01 | 1255475–1255976 | 19.0–26.6 |
| 2019 | I | 67.7 | pPt-558970–pPt-473819 | 4.87 | 11.9 | P2H | 01 | 73152139–7315272674921680–74922384 | 61.2–71.7 |
| 17–19 | I | 62.5 | capPt-672722 | 3.78 | 9.3 | P1 | 01 | 73308821–73309676 | 58.1–69.8 |
| 2018 | II | 77.9 | pPt-536505 | 3.43 | 8.8 | P1 | 02 | 14674331–14674870 | 77.3–77.9 |
| 2019 | II | 4.5 | pPt-536998 | 3.31 | 8.3 | P2 | 02 | 42632179–42632477 | 4.4–6.5 |
| 2019 | II | 15.2 | pPt-538350 | 3.42 | 8.5 | H | 02 | 41355373–41356084 | 15.2–15.3 |
| 2019 | II | 74.7 | pPt-456871 | 3.27 | 8.2 | H | 02 | 14674331–14674875 | 74.6–75.7 |
| 17–19 | II | 52.5 | pPt-457616 | 3.99 | 9.8 | P2 | 02 | 32226075–32226674 | 52.0–52.7 |
| 2018 | VII | 36.8 | pPt-459331 | 6.58 | 16.2 | P1 | 07 | 47918885–47919484 | 13.1–67.7 |
| 17–19 | VII | 43.0 | pPt-552007 | 4.39 | 10.7 | H | 07 | 32689974–32690374 | 36.8–53.9 |
| 2019 | IX | 17.7 | pPt-653179 | 3.72 | 9.2 | H | 09 | 8316412–8316923 | 16.9–18.8 |
| 2018 | XII | 125.3 | pPt-655073–pPt-456622 | 3.28 | 8.5 | P1 | 02 | 29779568–29780095 | 121.6–128.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobkowiak, S.; Janiszewska, M.; Stefańczyk, E.; Wasilewicz-Flis, I.; Śliwka, J. Quantitative Trait Loci for Resistance to Potato Dry Rot Caused by Fusarium sambucinum. Agronomy 2022, 12, 203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010203
Sobkowiak S, Janiszewska M, Stefańczyk E, Wasilewicz-Flis I, Śliwka J. Quantitative Trait Loci for Resistance to Potato Dry Rot Caused by Fusarium sambucinum. Agronomy. 2022; 12(1):203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010203
Chicago/Turabian StyleSobkowiak, Sylwester, Marta Janiszewska, Emil Stefańczyk, Iwona Wasilewicz-Flis, and Jadwiga Śliwka. 2022. "Quantitative Trait Loci for Resistance to Potato Dry Rot Caused by Fusarium sambucinum" Agronomy 12, no. 1: 203. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010203