
Toxic Impact of Soil Microplastics (PVC) on Two Weeds: Changes in Growth, Phenology and Photosynthesis Efficiency

 , , , and
, , , and 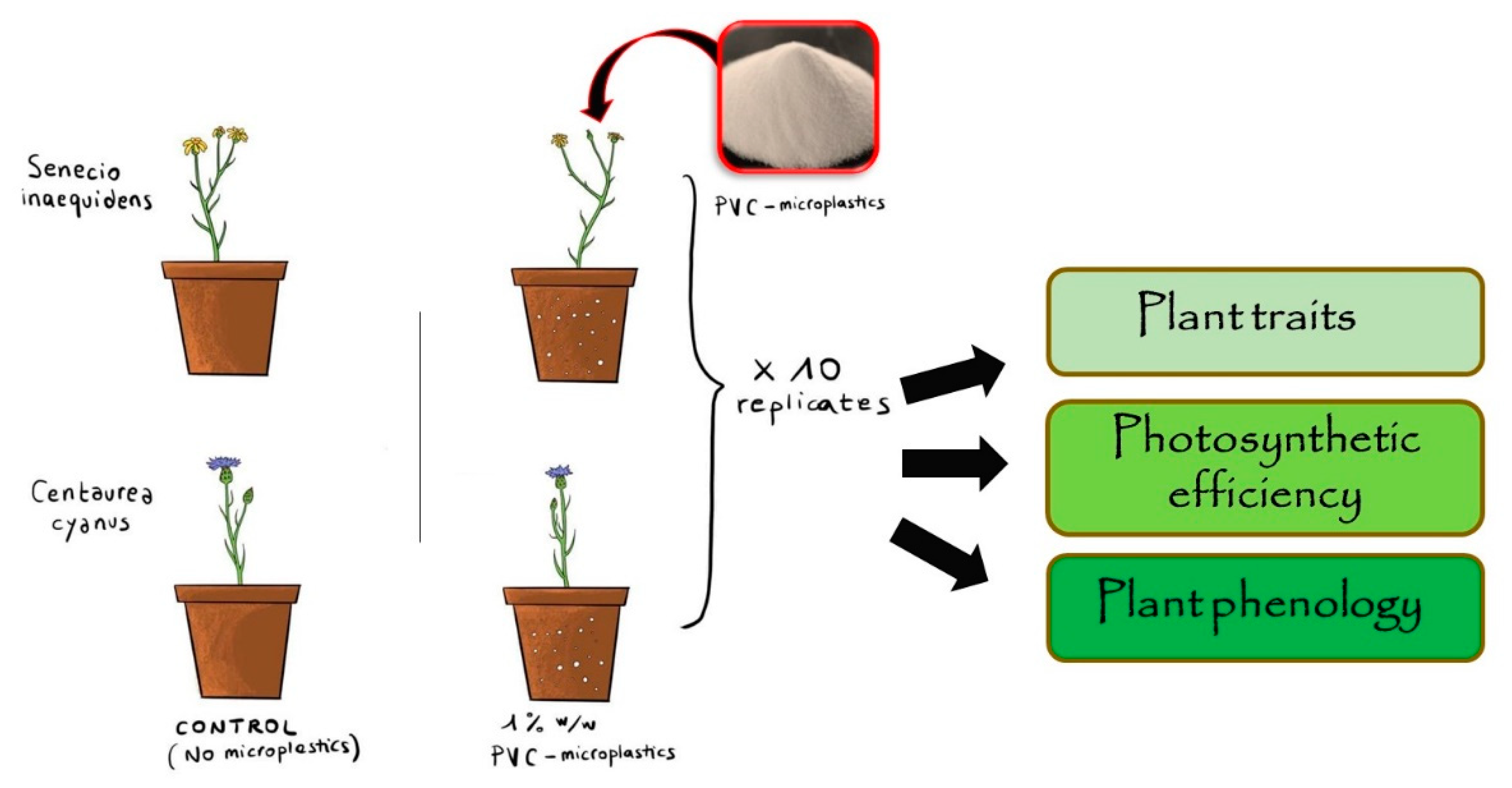{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
- (a)
- C. cyanus is an annual therophyte native to the Eastern Mediterranean and now subcosmopolite having expanded its range following wheat cultivation (archaeophyte; [28]). C. cyanus historically spread worldwide associated with cereal crops as segetal species. In the invasion range (Africa, America, and Asia), the species can be found in several habitats such as grasslands, woodlands, roadsides and disturbed sites. For this experiment, seeds were collected in a cultivated area of the Po valley (Busto Arsizio, N-Italy; 45.599663 N; 8.816384 E).
- (b)
- S. inaequidens is a perennial chamaephyte native to South Africa and highly invasive in Europe. S. inaequidens has been introduced in Europe by the end of the 19th century with sheep’s wool commerce [29]. It is currently invasive in many European countries where it can be mostly found in disturbed sites, such as roadsides, railways embankments and quarries, but also in semi-natural grasslands and vineyards [30]. For this experiment, seeds of S. inaequidens were collected in a highly invaded area in the former quarry of Collepedrino (Bergamo, N-Italy; 45.779639 N; 9.523546 E).
2.2. Exposure of Plants to Soil Microplastics
2.3. Measurements of Growth Parameters, Phenology and Photosynthetic Efficiency
2.4. Data Analysis
- (a)
- Linear model, Y = b0, + b1X, where b0 is the value of Y when X = 0 while b1 is the slope;
- (b)
- Exponential, Y = aekX, where a is the value of Y when X = 0, while k represents the relative increase or decrease in Y for a unit increase in X;
- (c)
- The three-parameter logistic, , where d is the upper asymptote, e is X value producing a response half-way between d and 0, while b is the slope around the inflection point.
- (d)
- Gompertz, Y = c + (d)exp {−exp [b (X − e)]}, where the parameters have the same meaning as those in the logistic function.
3. Results
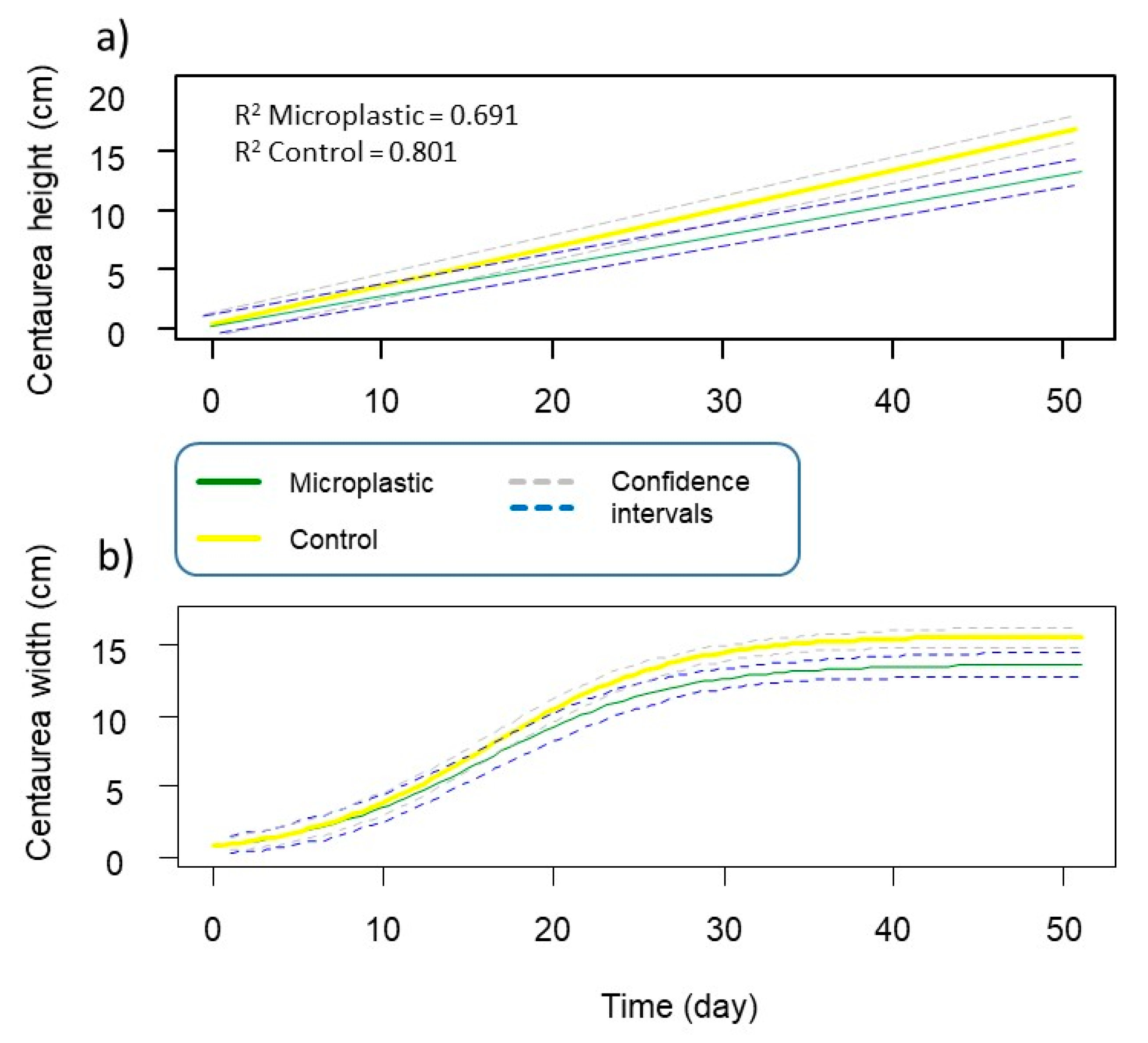
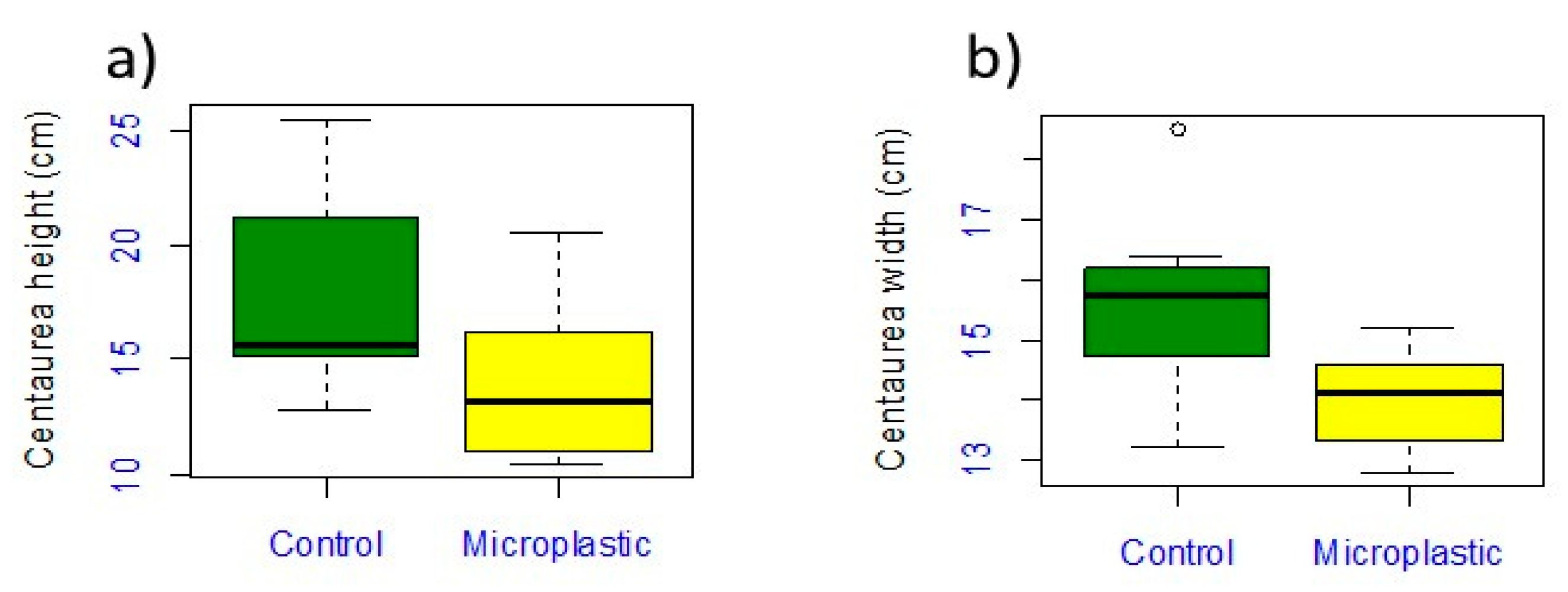
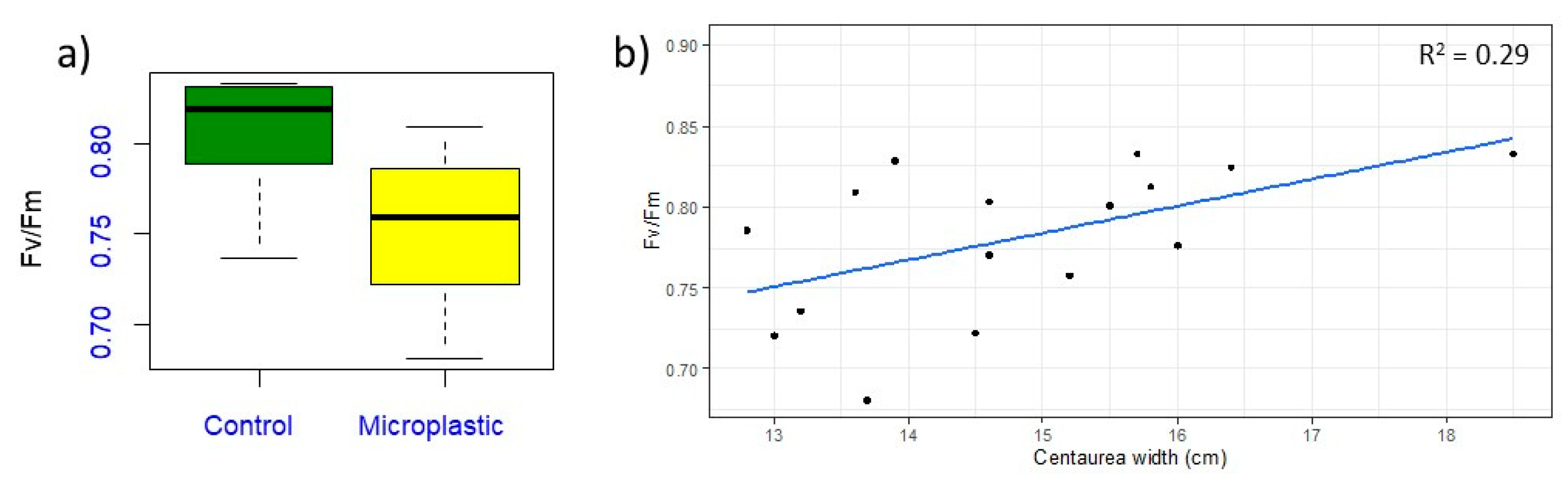
3.1. Centaurea cyanus
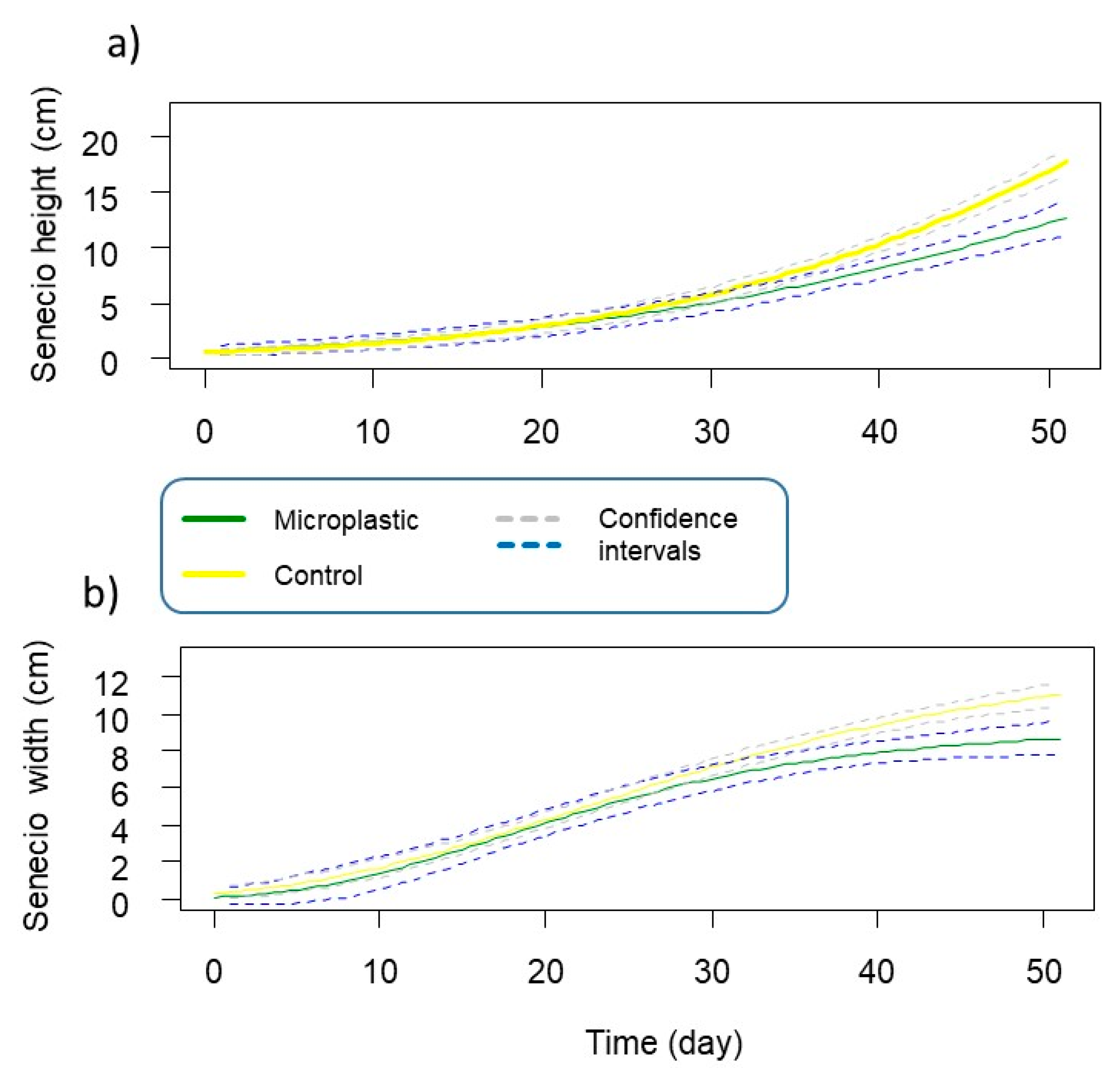
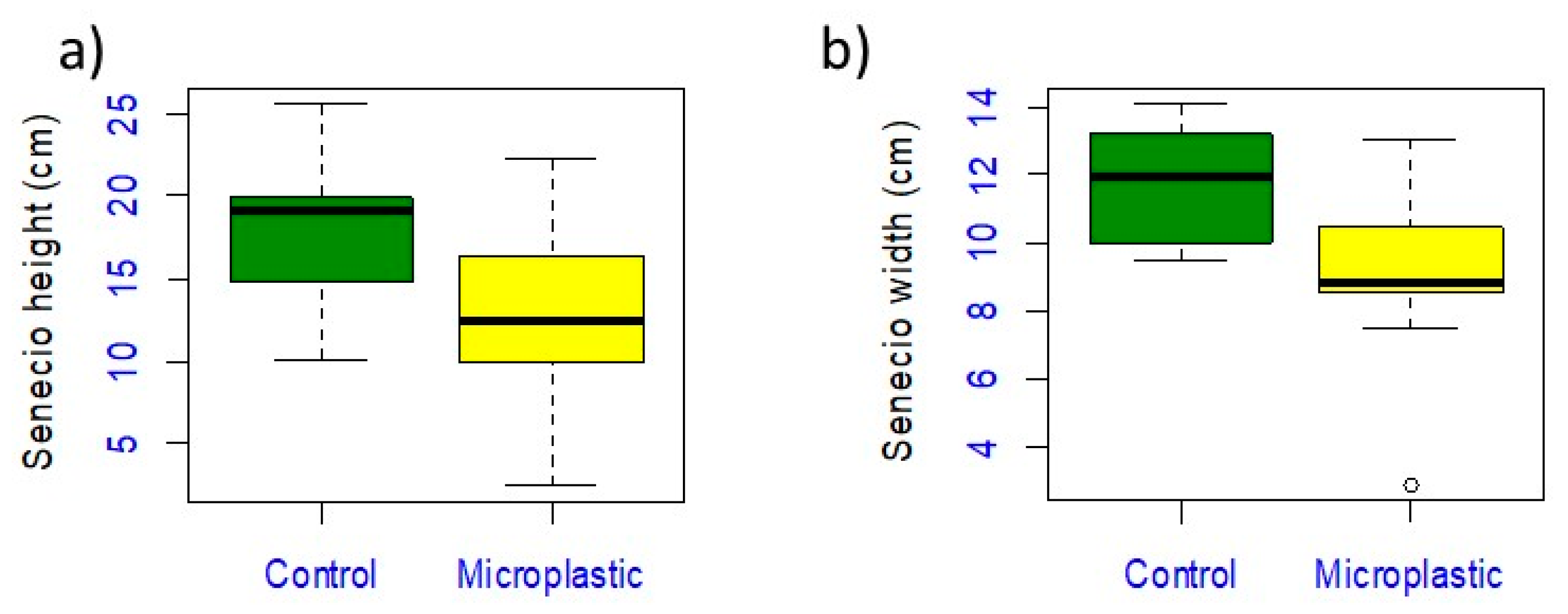
3.2. Senecio inaequidens
4. Discussion
- (a)
- (b)
- (c)
- (d)
- The alteration of the soil’s physical properties, with potential modifications in moisture retention and root penetration dynamics [50];
- (e)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barrows, A.P.W.; Christiansen, K.S.; Bode, E.T.; Hoellein, T.J. A watershed-scale, citizen science approach to quantifying microplastic concentration in a mixed land-use river. Water Res. 2018, 147, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Rao, Q.; Deng, X.; Chen, J.; Xie, P. Rainfall is a significant environmental factor of microplastic pollution in inland waters. Sci. Total Environ. 2020, 732, 139065. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Zhang, Q.; Zhang, Z.; Hu, B.; Chen, J.; Chen, J.; Qian, H. Pollutant toxicology with respect to microalgae and cyanobacteria. J. Environ. Sci. 2021, 99, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in freshwater and terrestrial environments: Evaluating the current understanding to identify the knowledge gaps and future research priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Nizzetto, L.; Bussi, G.; Futter, M.N.; Butterfield, D.; Whitehead, P.G. A theoretical assessment of microplastic transport in river catchments and their retention by soils and river sediments. Environ. Sci. Process. Impacts 2016, 18, 1050–1059. [Google Scholar] [CrossRef]
- Verla, A.W.; Enyoh, C.E.; Verla, E.N. Microplastics, an emerging concern: A review of analytical techniques for detecting and quantifying microplastic. Anal. Methods Environ. Chem. J. 2019, 2, 15–32. [Google Scholar] [CrossRef]
- Kelessidis, A.; Stasinakis, A.S. Comparative study of the methods used for treatment and final disposal of sewage sludge in European countries. Waste Manag. 2012, 32, 1186–1195. [Google Scholar] [CrossRef]
- Singh, R.P.; Agrawal, M. Potential benefits and risks of land application of sewage sludge. Waste Manag. 2008, 28, 347–358. [Google Scholar] [CrossRef]
- Ambrosini, R.; Azzoni, R.S.; Pittino, F.; Diolaiuti, G.; Franzetti, A.; Parolini, M. First evidence of microplastic contamination in the supraglacial debris of an alpine glacier. Environ. Pollut. 2019, 253, 297–301. [Google Scholar] [CrossRef]
- Dris, R.; Gasperi, J.; Saad, M.; Mirande, C.; Tassin, B. Synthetic fibers in atmospheric fallout: A source of microplastics in the environment? Mar. Pollut. Bull. 2016, 104, 290–293. [Google Scholar] [CrossRef]
- Farrell, P.; Nelson, K. Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.E.; Hamann, M.; Kroon, F.J. Bioaccumulation and biomagnification of microplastics in marine organisms: A review and meta-analysis of current data. PLoS ONE 2020, 15, e0240792. [Google Scholar] [CrossRef] [PubMed]
- De Souza Machado, A.A.; Lau, C.W.; Kloas, W.; Bergmann, J.; Bachelier, J.B.; Faltin, E.; Becker, R.; Görlich, A.S.; Rillig, M.C. Microplastics can change soil properties and affect plant performance. Environ. Sci. Technol. 2019, 53, 6044–6052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza Machado, A.A.; Lau, C.W.; Till, J.; Kloas, W.; Lehmann, A.; Becker, R.; Rillig, M.C. Impacts of microplastics on the soil biophysical environment. Environ. Sci. Technol. 2018, 52, 9656–9665. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Yang, X.; Pelaez, A.M.; Huerta Lwanga, E.; Beriot, N.; Gertsen, H.; Garbeva, P.; Geissen, V. Macro- and micro- plastics in soil-plant system: Effects of plastic mulch film residues on wheat (Triticum aestivum) growth. Sci. Total Environ. 2018, 645, 1048–1056. [Google Scholar] [CrossRef]
- Dong, H.G.; Liu, T.; Han, Z.Q.; Sun, Q.M.; Li, R. Determining time limits of continuous film mulching and examining residual effects on cotton yield and soil properties. J. Environ. Biol. 2015, 36, 677–684. [Google Scholar]
- Urbina, M.A.; Correa, F.; Aburto, F.; Ferrio, J.P. Adsorption of polyethylene microbeads and physiological effects on hydroponic maize. Sci. Total Environ. 2020, 741, 140216. [Google Scholar] [CrossRef]
- Qi, Y.; Ossowicki, A.; Yang, X.; Huerta Lwanga, E.; Dini-Andreote, F.; Geissen, V.; Garbev, P. Effects of plastic mulch film residues on wheat rhizosphere and soil properties. J. Hazard. Mater. 2020, 387, 121711. [Google Scholar] [CrossRef]
- Meng, F.; Yang, X.; Riksen, M.; Xu, M.; Geissen, V. Response of common bean (Phaseolus vulgaris L.) growth to soil contaminated with microplastics. Sci. Total Environ. 2021, 755, 142516. [Google Scholar] [CrossRef]
- Li, Z.; Li, Q.; Li, R.; Zhao, Y.; Geng, J.; Wang, G. Physiological responses of lettuce (Lactuca sativa L.) to microplastic pollution. Environ. Sci. Pollut. Res. 2020, 27, 30306–30314. [Google Scholar] [CrossRef]
- Li, Z.; Li, R.; Li, Q.; Zhou, J.; Wang, G. Physiological response of cucumber (Cucumis sativus L.) leaves to polystyrene nanoplastics pollution. Chemosphere 2020, 255, 127041. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, H.; Liao, Y.; Ye, Z.; Li, M.; Klobucar, G. Ecotoxicity and genotoxicity of polystyrene microplastics on higher plant Vicia faba. Environ. Pollut. 2019, 250, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Cárdenas, A.; van Pelt, F.N.A.M.; O’Halloran, J.; Jansen, M.A.K. Adsorption, uptake and toxicity of micro- and nanoplastics: Effects on terrestrial plants and aquatic macrophytes. Environ. Pollut. 2021, 84, 117183. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Song, S.; Xu, X.; Ma, Q.; Lu, Y. Sources, migration, accumulation and influence of microplastics in terrestrial plant communities. Environ. Exp. Bot. 2021, 192, 104635. [Google Scholar] [CrossRef]
- Menicagli, V.; Balestri, E.; Lardicci, C. Exposure of coastal dune vegetation to plastic bag leachates: A neglected impact of plastic litter. Sci. Total Environ. 2019, 683, 737–748. [Google Scholar] [CrossRef]
- Menicagli, V.; Balestri, E.; Vallerini, F.; Castelli, A.; Lardicci, C. Combined effect of plastic litter and increased atmospheric nitrogen deposition on vegetative propagules of dune plants: A further threat to coastal ecosystems. Environ. Pollut. 2020, 266, 115281. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Rillig, M.C. Effects of Microplastic fibers and drought on plant communities. Environ. Sci. Technol. 2020, 54, 6166–6173. [Google Scholar] [CrossRef]
- Bakels, C.C. The early history of Cornflower (Centaurea cyanus L.) in the Netherlands. Acta Palaeobot. 2012, 52, 25–31. [Google Scholar]
- Ernst, W.H.O. Invasion, dispersal and ecology of the South African neophyte Senecio inaequidens in The Netherlands: From wool alien to railway and road alien. Acta Bot. Neerl. 1998, 47, 131–151. [Google Scholar]
- Heger, T.; Böhmer, H.J. The invasion of Central Europe by Senecio inaequidens DC.—A complex biogeographical problem. Erdkunde 2005, 59, 34–49. [Google Scholar] [CrossRef] [Green Version]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jajoo, A.; Mehta, P.; Bharti, S. Analysis of elevated temperature-induced inhibition of photosystem II using chlorophyll a fluorescence induction kinetics in wheat leaves (Triticum aestivum). Plant Biol. 2011, 13, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Paine, C.E.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Onofri, A. Some Useful Equations for Nonlinear Regression in R. 2019. Available online: https://www.statforbiology.com/nonlinearregression/usefulequations (accessed on 2 May 2022).
- Onofri, A. Aomisc: Statistical Methods for the Agricultural Sciences, R Package Version 0.62. 2021. Available online: https://rdrr.io/github/OnofriAndreaPG/aomisc/src/R/TukeyNA.r (accessed on 2 May 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 May 2022).
- Zhang, M.; Liu, Y.H.; Xu, W.; Smith, W.; Murray, S.C.; Zhang, H.-B. Analysis of the genes controlling three quantitative traits in three diverse plant species reveals the molecular basis of quantitative traits. Sci. Rep. 2020, 10, 10074. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Devriese, L.; Galgani, F.; Robbens, J.; Janssen, C.R. Microplastics in sediments: A review of techniques, occurrence and effects. Mar. Environ. Res. 2015, 111, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalčíková, G.; Žgajnar Gotvajn, A.; Kladnik, A.; Jemec, A. Impact of polyethylene microbeads on the floating freshwater plant duckweed Lemna minor. Environ Pollut. 2017, 230, 1108–1115. [Google Scholar] [CrossRef]
- Bandmann, V.; Müller, J.D.; Köhler, T.; Homann, U. Uptake of fluorescent nano beads into BY2-cells involves clathrin-dependent and clathrin-independent endocytosis. Fed. Eur. Biochem. Soc. Lett. 2012, 586, 3626–3632. [Google Scholar] [CrossRef] [Green Version]
- Spanò, C.; Muccifora, S.; Ruffini Castiglione, M.; Bellani, L.; Bottega, S.; Giorgetti, L. Polystyrene nanoplastics affect seed germination, cell biology and physiology of rice seedlings in-short term treatments: Evidence of their internalization and translocation. Plant Phys. Biochem. 2022, 172, 158–166. [Google Scholar] [CrossRef]
- Colzi, I.; Renna, L.; Bianchi, E.; Castellani, M.B.; Coppi, A.; Pignattelli, S.; Loppi, S.; Gonnelli, C. Impact of microplastics on growth, photosynthesis and essential elements in Cucurbita pepo L. J. Hazard. Mater. 2022, 423, 127238. [Google Scholar] [CrossRef]
- Zhou, J.; Cao, Y.; Liu, X.; Jiang, H.; Li, W. Bladder entrance of microplastic likely induces toxic effects in carnivorous macrophyte Utricularia aurea Lour. Environ. Sci. Pollut. Res. Int. 2020, 27, 32124–32131. [Google Scholar] [CrossRef]
- Pignattelli, S.; Broccoli, A.; Renzi, M. Physiological responses of garden cress (L. sativum) to different types of microplastics. Sci. Total Environ. 2020, 727, 138609. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Yan, Z.; Shen, R.; Wang, M.; Huang, Y.; Ren, H.; Zhang, Y.; Lemos, B. Microplastics release phthalate esters and cause aggravated adverse effects in the mouse gut. Environ. Int. 2020, 143, 105916. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Kaur, R. A review on morpho-physiological traits of plants under phthalates stress and insights into their uptake and translocation. Plant Growth Regul. 2020, 91, 327–347. [Google Scholar] [CrossRef]
- Verla, A.W.; Enyoh, C.E.; Verla, E.N.; Nwarnorh, K.O. Microplastic–toxic chemical interaction: A review study on quantified levels, mechanism and implication. SN Appl. Sci. 2019, 1, 1400. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Changyin, Z.; Chao, W.; Cheng, G. Occurrence and ecological impacts of microplastics in soil systems: A review. Bull. Environ. Contam. Toxicol. 2019, 102, 741–749. [Google Scholar] [CrossRef]
- Yan, Y.; Chen, Z.; Zhu, F.; Zhu, C.; Wang, C.; Gu, C. Effect of Polyvinyl Chloride microplastics on bacterial community and nutrient status in two agricultural soils. Bull. Environ. Contam. Toxicol. 2021, 107, 602–609. [Google Scholar] [CrossRef]
- Boots, B.; Russell, C.W.; Green, D.S. Effects of Microplastics in soil ecosystems: Above and below ground. Environ. Sci. Technol. 2019, 53, 11496–11506. [Google Scholar] [CrossRef]
- Zhao, T.; Lozano, Y.M.; Rillig, M.C. Microplastics increase soil pH and decrease microbial activities as a function of microplastic shape, polymer type, and exposure time. Front. Environ. Sci. 2021, 9, 675803. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, Y.; Wang, J.; Zhang, M.; Jia, W.; Qin, X. LDPE microplastic films alter microbial community composition and enzymatic activities in soil. Environ. Pollut. 2019, 254 Pt A, 112983. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Lehnert, T.; Linck, L.T.; Lehmann, A.; Rillig, M.C. Microplastic shape, polymer type, and concentration affect soil properties and plant biomass. Front. Plant Sci. 2021, 12, 616645. [Google Scholar] [CrossRef]
- Van Weert, S.; Redondo-Hasselerharm, P.E.; Diepens, N.J.; Koelmans, A.A. Effects of nanoplastics and microplastics on the growth of sediment-rooted macrophytes. Sci. Total Environ. 2019, 654, 1040–1047. [Google Scholar] [CrossRef]
- Rillig, M.C.; Ingraffia, R.; de Souza Machado, A.A. Microplastic incorporation into soil in agroecosystems. Front. Plant Sci. 2017, 8, 1805. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentili, R.; Quaglini, L.; Cardarelli, E.; Caronni, S.; Montagnani, C.; Citterio, S. Toxic Impact of Soil Microplastics (PVC) on Two Weeds: Changes in Growth, Phenology and Photosynthesis Efficiency. Agronomy 2022, 12, 1219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051219
Gentili R, Quaglini L, Cardarelli E, Caronni S, Montagnani C, Citterio S. Toxic Impact of Soil Microplastics (PVC) on Two Weeds: Changes in Growth, Phenology and Photosynthesis Efficiency. Agronomy. 2022; 12(5):1219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051219
Chicago/Turabian StyleGentili, Rodolfo, Lara Quaglini, Elisa Cardarelli, Sarah Caronni, Chiara Montagnani, and Sandra Citterio. 2022. "Toxic Impact of Soil Microplastics (PVC) on Two Weeds: Changes in Growth, Phenology and Photosynthesis Efficiency" Agronomy 12, no. 5: 1219. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051219