Assessing the Individual and Combined Effects of QTL for Adult Plant Stripe Rust Resistance Derived from Cappelle-Desprez

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Pathogen Materials
2.2. Phenotypic Analysis
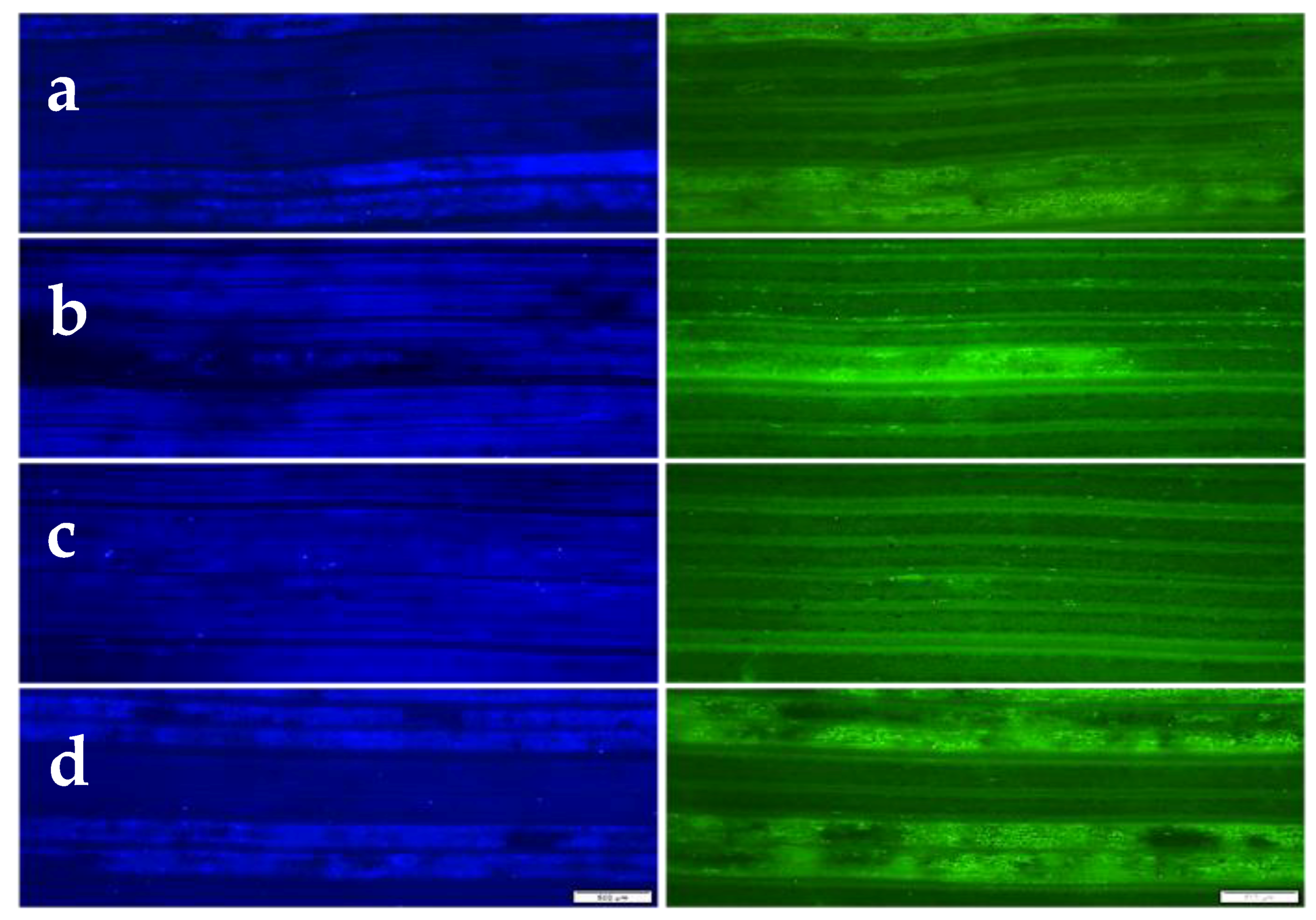
2.3. Fluorescence Microscopy
2.4. Molecular Analysis
2.5. Statistical Analysis
3. Results
3.1. Phenotypic Analysis of Stripe Rust Field Infections
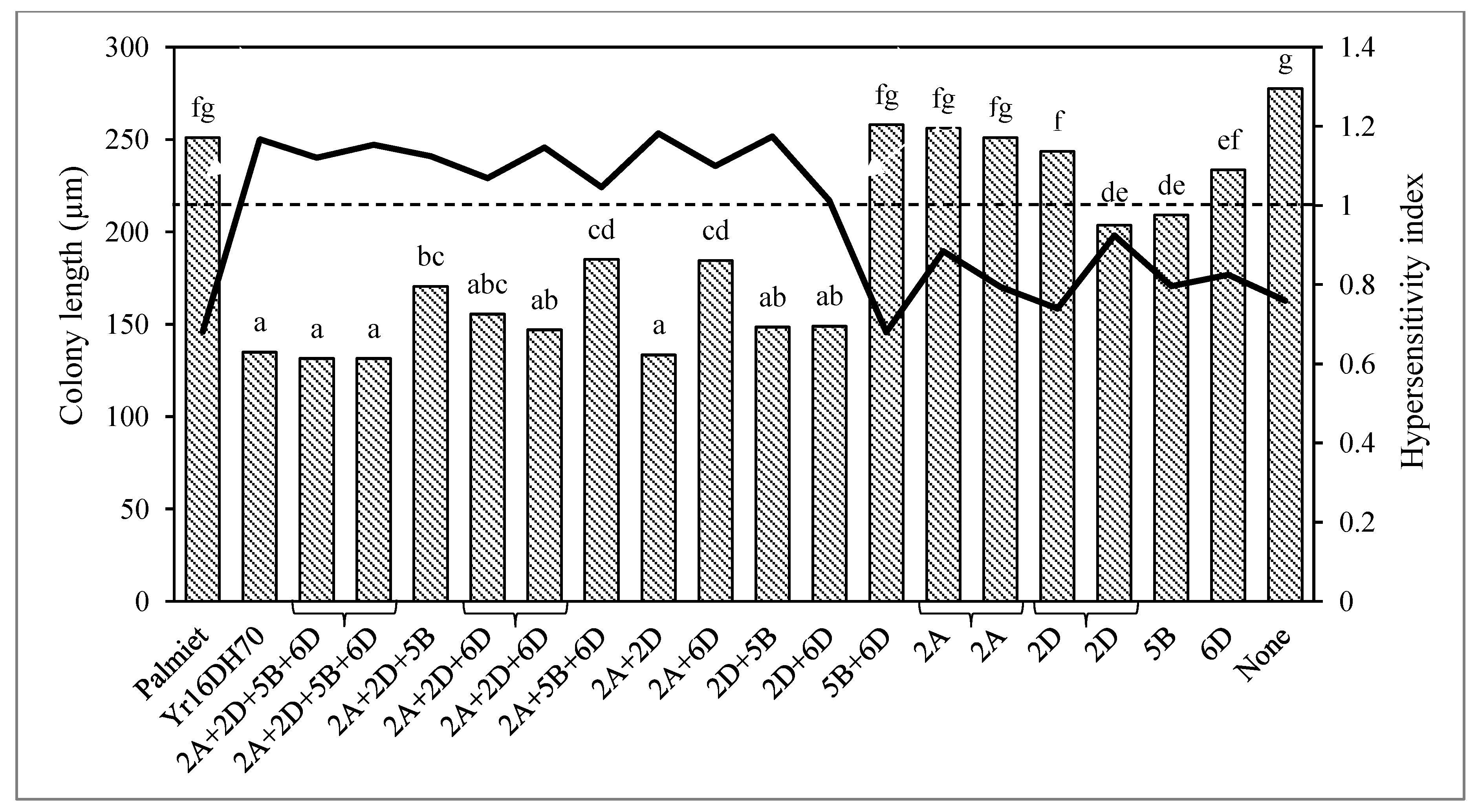
3.2. Microscopy of Puccinia striiformis f. sp. tritici Development and Host Cell Necrosis
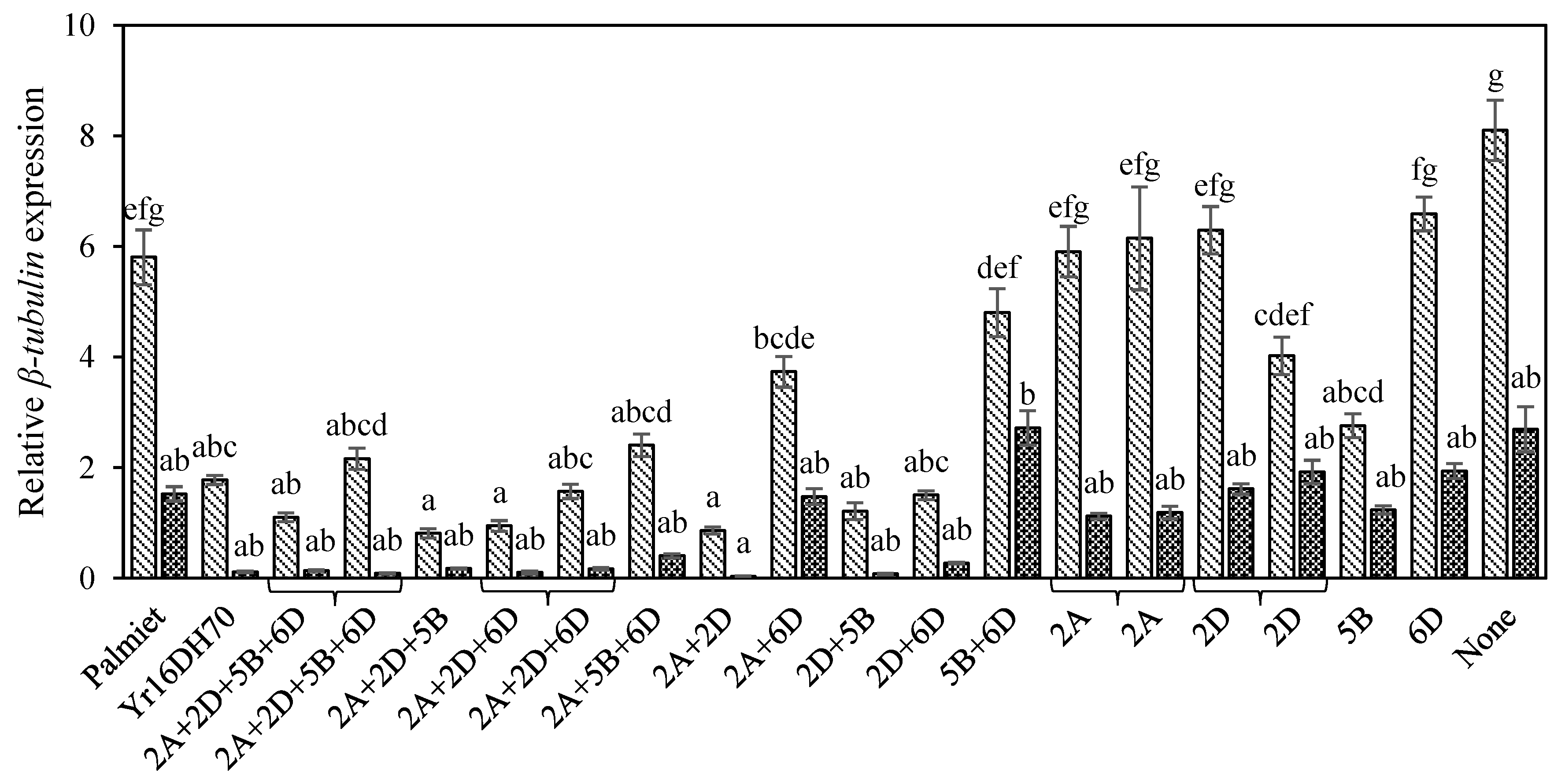
3.3. Molecular Estimation of Fungal Biomass
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef] [PubMed]
- Lagudah, E.S. Molecular genetics of race non-specific rust resistance in wheat. Euphytica 2011, 179, 81–91. [Google Scholar] [CrossRef]
- Boyd, L.A. Can Robigus defeat an old enemy?—Yellow rust of wheat. J. Agric. Sci. 2005, 143, 233–343. [Google Scholar] [CrossRef]
- Ali, S.; Rodriguez-Algaba1, J.; Thach, T.; Sørensen, C.K.; Hansen, J.G.; Lassen, P.; Nazari, K.; Hodson, D.P.; Justesen, A.F.; Hovmøller, M.S. Yellow rust epidemics worldwide were caused by pathogen races from divergent genetic lineages. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; Enjalbert, J.; De Vallavieille-Pope, C. Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f. sp. tritici. PLoS Pathog. 2014, 10, e1003903. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Xia, X.C. Catalogue of Gene symbols for wheat: 2017 Supplement. 2017. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 7 December 2018).
- Chen, X.M. Review article: High-temperature adult-plant resistance, key for sustainable control of stripe rust. AJPS 2013, 4, 608–627. [Google Scholar] [CrossRef]
- Wang, M.; Chen, X. Stripe Rust Resistance. In Stripe Rust; Chen, X., Kang, Z., Eds.; Springer: Dordrecht, The Netherlands, 2017; Available online: https://0-doi-org.brum.beds.ac.uk/10.1007/978-94-024-1111-9_5 (accessed on 10 January 2019).
- Mundt, C.C. Pyramiding for resistance durability: Theory and Practice. Phytopathology 2018, 108, 792–802. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; William, H.M. Genetics and breeding for durable resistance to leaf and stripe rusts in wheat. Turk. J. Agric. For. 2005, 29, 121–127. [Google Scholar]
- Sørensen, C.K.; Hovmøller, M.S.; Leconte, M.; Dedryver, F.; de Vallavieille-Pope, C. New races of Puccinia striiformis found in Europe reveal race specificity of long-term effective adult plant resistance in wheat. Phytopathology 2014, 104, 1042–1051. [Google Scholar] [CrossRef]
- Jagger, I.J.; Newell, C.; Berry, S.T.; MacCormack, R.; Boyd, L.A. The genetic characterisation of stripe rust resistance in the German wheat cultivar Alcedo. Theor. Appl. Genet. 2011, 122, 723–733. [Google Scholar] [CrossRef]
- Boshoff, W.H.P.; Pretorius, Z.A.; Van Niekerk, B.D. Resistance in South African and foreign wheat cultivars to pathotypes 6E16A- and 6E22A- of Puccinia striiformis f. sp. tritici. S. Afr. J. Plant Soil 2002, 19, 27–36. [Google Scholar] [CrossRef]
- Agenbag, G.M.; Pretorius, Z.A.; Boyd, L.A.; Bender, C.M.; Prins, R. Identification of adult plant resistance to stripe rust in the wheat cultivar Cappelle-Desprez. Theor. Appl. Genet. 2012, 125, 109–120. [Google Scholar] [CrossRef] [PubMed]
- De Vallavieille-Pope, C.; Picard-Formery, H.; Radulovic, S.; Johnson, R. Specific resistance factors to yellow rust in seedlings of some French wheat varieties and races of Puccinia striiformis Westend in France. Agronomie 1990, 2, 103–113. [Google Scholar] [CrossRef]
- Chen, X.M.; Jones, S.S.; Line, R.F. Chromosomal location of genes for resistance to Puccinia striiformis in seven wheat cultivars with resistance genes at the Yr3 and Yr4 loci. Phytopathology 1996, 86, 1228–1233. [Google Scholar] [CrossRef]
- Worland, A.J.; Law, C.N. Genetic analysis of chromosome 2D of wheat. Z. Pflanzenzüchtung 1986, 96, 331–345. [Google Scholar]
- Law, C.N.; Worland, A.J. The control of adult-plant resistance to yellow rust by the translocated chromosome 5BS-7BS of bread wheat. Plant Breed. 1997, 116, 59–63. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.A. Close genetic linkage of genes conferring adult plant resistance to leaf rust and stripe rust in wheat. Plant Pathol. 1992, 41, 523–527. [Google Scholar] [CrossRef]
- Mallard, S.; Gaudet, D.; Aldeia, A.; Abelard, C.; Besnard, A.L.; Sourdille, P.; Dedryver, F. Genetic analysis of durable resistance to yellow rust in bread wheat. Theor. Appl. Genet. 2005, 110, 1401–1409. [Google Scholar] [CrossRef] [PubMed]
- Boukhatem, N.; Baret, P.V.; Mingeot, D.; Jacquemin, J.M. Quantitative trait loci for resistance against yellow rust in two wheat derived recombinant inbred line populations. Theor. Appl. Genet. 2002, 104, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Chen, Z.; Wang, Y.; Bland, D.; Buck, J.; Brown-Guedira, G.; Johnson, J. Characterization of a major QTL for adult plant resistance to stripe rust in US soft red winter wheat. Theor. Appl. Genet. 2011, 123, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Jones, S.S.; Line, R.F. Chromosomal location of genes for stripe rust resistance in spring wheat cultivars Compair, Fielder, Lee, and Lemhi and interactions of aneuploid wheats with races of Puccinia striiformis. Phytopathology 1995, 85, 375–381. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Visser, B.; Herselman, L.; Pretorius, Z.A. Microsatellite characterisation of South African Puccinia striiformis races. S. Afr. J. Plant Soil 2016, 33, 161–166. [Google Scholar] [CrossRef]
- Peterson, R.F.; Champbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity of leaves and stem of cereals. Can. J. Res. 1948, 26c, 496–500. [Google Scholar] [CrossRef]
- Stubbs, R.W.; Prescott, J.M.; Saari, E.E.; Dubin, H.J. Cereal Disease Methodology Manual; Centro Internacional de Mejoramiento de Maίz yTrigo (CIMMYT): Texcoco, Mexico, 1986; Available online: https://repository.cimmyt.org/handle/10883/3997 (accessed on 7 December 2018).
- Rohringer, R.; Kim, W.K.; Samborsky, D.J.; Howes, N.K. Calcofluor: An optical brightener for fluorescence microscopy of fungal plant parasites in leaves. Phytopathology 1977, 67, 808–810. [Google Scholar] [CrossRef]
- Niks, R.E.; Dekens, R.G. Histological studies on the infection of triticale, wheat and rye by Puccinia recondita f. sp. tritici and P. recondita f. sp. recondita. Euphytica 1987, 36, 275–285. [Google Scholar] [CrossRef]
- Moldenhauer, J.; Moerschbacher, B.M.; Van der Westhuizen, A.J. Histological investigation of stripe rust (Puccinia striiformis f. sp. tritici) development in resistant and susceptible wheat cultivars. Plant Pathol. 2006, 55, 469–474. [Google Scholar] [CrossRef]
- Dugyala, S.; Borowicz, P.; Acevedo, M. Rapid protocol for visualization of rust fungi structures using fluorochrome Uvitex 2B. Plant Methods 2015, 11, 54. [Google Scholar] [CrossRef]
- Kloppers, F.J.; Pretorius, Z.A. Effects of combinations amongst genes Lr13, Lr34 and Lr37 on components of resistance in wheat to leaf rust. Plant Pathol. 1997, 46, 737–750. [Google Scholar] [CrossRef]
- Coram, T.E.; Settles, M.L.; Chen, X.M. Transcriptome analysis of high-temperature adult-plant resistance conditioned by Yr39 during the wheat—Puccinia striiformis f. sp. tritici interaction. Mol. Plant Pathol. 2008, 9, 479–493. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2000; pp. A8.19–A8.21; 7.31–7.34. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Ling, P.; Wang, M.N.; Chen, X.M.; Campbell, K.G. Construction and characterization of a full-length cDNA library for the wheat stripe rust pathogen (Puccinia striiformis f. sp. tritici). BMC Genom. 2007, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Scholtz, J.J.; Visser, B. Reference gene selection for qPCR gene expression analysis of rust-infected wheat. Physiol. Mol. Plant Pathol. 2013, 81, 22–25. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Wellings, C.R.; Park, R.F. Wheat Rusts: An Atlas of Resistance Genes; CSIRO: Canberra, Australia, 1995; 200p. [Google Scholar]
- Singh, R.P.; Herrera-Foessel, S.; Huerta-Espino, J.; Singh, S.; Bhavani, S.; Lan, C.; Basnet, B.R. Progress towards genetics and breeding for minor genes based resistance to Ug99 and other rusts in CIMMYT high-yielding spring wheat. J. Integr. Agric. 2014, 13, 255–261. [Google Scholar] [CrossRef]
- Periyannan, S.; Milne, R.J.; Figueroa, M.; Lagudah, E.S.; Dodds, P.N. An overview of genetic rust resistance: From broad to specific mechanisms. PLoS Pathog. 2017, 13, e1006380. [Google Scholar] [CrossRef]
- Germán, S.; Kolmer, J.A. Effect of gene Lr34 in the enhancement of resistance to leaf rust of wheat. Theor. Appl. Genet. 1992, 84, 97–105. [Google Scholar] [CrossRef]
- Kolmer, J.A. Enhanced leaf rust resistance in wheat conditioned by resistance gene pairs with Lr13. Euphytica 1992, 61, 123–130. [Google Scholar] [CrossRef]
- Knott, D.R. The inheritance of resistance to stem rust races 56 and 15B-1L (Can.) in the wheat varieties Hope and H-44. Can. J. Genet. Cytol. 1968, 10, 311–320. [Google Scholar] [CrossRef]
- Maree, G.J.; Prins, R.; Bender, C.M.; Boshoff, W.H.P.; Negussie, T.G.; Pretorius, Z.A. Phenotyping Kariega × Avocet S doubled haploid lines containing individual and combined adult plant stripe rust resistance loci. Plant Pathol. 2019. [Google Scholar] [CrossRef]
- Kloppers, F.J.; (Pannar Seed [Pty] Ltd., Greytown, KwaZulu-Natal, South Africa). Personal Communication, 2015.




{kind=link}
{kind=link}
{kind=link}
| Parent/RIL | QTL 1 | First Sampling | Second Sampling | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LAI 2 (%) | RT 3 Range | Mean CI 2 | LAI 2 (%) | RT 3 Range | Mean CI 2 | ||||
| Range | Mean | Range | Mean | ||||||
| Palmiet | 4B | 30 | 30 efgh | MRMS | 18 cde | 70 | 70 ef | MRMS | 42 ij |
| Yr16DH70 | 2A+2D+5B+6D | 10 | 10 a | MR | 4 a | 20 | 20 a | R | 4 a |
| 20 | 2A+2D+5B+6D (4) | 20–25 | 22.5 bcde | MRMS | 13.5 abcd | 20–30 | 25 ab | R-RMR | 6 a |
| 148 | 2A+2D+5B+6D (4) | 10–20 | 15 abc | MR-MRMS | 8 abc | 20–30 | 25 ab | RMR | 7.5 a |
| 39 | 2A+2D+5B (3) | 10–15 | 12.5 ab | MRMS | 7.5 ab | 30–40 | 35 bc | R-MR | 10 abc |
| 34 | 2A+2D+6D (3) | 15–20 | 17.5 abcd | MRMS | 10.5 abc | 50–60 | 55 d | R-MRMS | 21 def |
| 142 | 2A+2D+6D (3) | 15–20 | 17.5 abcd | MRMS | 10.5 abc | 25–30 | 27.5 ab | RMR | 8.25 ab |
| 33 | 2A+5B+6D (3) | 20–30 | 25 cdef | MRMS | 15 bcd | 40 | 40 c | RMR-MRMS | 18 bcde |
| 10 | 2A+2D (2) | 5–15 | 10 a | RMR-MR | 3.75 a | 25–30 | 27.5 ab | R-RMR | 6.75 a |
| 144 | 2A+6D (2) | 25–30 | 27.5 defg | MRMS | 16.5 bcd | 40 | 40 c | MR-MRMS | 20 cdef |
| 153 | 2D+5B (2) | 20–25 | 22.5 bcde | MRMS | 13.5 abcd | 50–70 | 60 de | R | 12 abcd |
| 17 | 2D+6D (2) | 15–20 | 17.5 abcd | MRMS | 10.5 abc | 30–35 | 32.5 bc | RMR | 9.75 abc |
| 45 | 5B+6D (2) | 30–40 | 35 fgh | MRMS | 21 de | 60–70 | 65 de | MRMS | 39 hi |
| 112 | 2A (1) | 40 | 40 hi | MRMS-MS | 28 ef | 40–70 | 55 d | MRMS-MS | 40 hi |
| 178 | 2A (1) | 35–40 | 37.5 ghi | MRMS | 22.5 de | 60 | 60 de | MR-MRMS | 30 fgh |
| 15 | 2D (1) | 30–50 | 40 hi | MS-MSS | 34.5 f | 80–90 | 85 g | MRMS | 51 jk |
| 190 | 2D (1) | 20–30 | 25 cdef | MRMS | 15 bcd | 50–60 | 55 d | MR-MRMS | 28 efg |
| 31 | 5B (1) | 30–40 | 35 fghi | MRMS | 21 de | 50–60 | 55 d | MRMS | 33 ghi |
| 149 | 6D (1) | 40 | 40 hi | MRMS-MS | 28 ef | 50–60 | 55 d | MRMS | 33 ghi |
| 52 | None (0) | 40–50 | 45 i | MRMS-MSS | 34.5 f | 70–90 | 80 fg | MRMS-MS | 57 k |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maree, G.J.; Prins, R.; Boyd, L.A.; Castelyn, H.D.; Bender, C.M.; Boshoff, W.H.; Pretorius, Z.A. Assessing the Individual and Combined Effects of QTL for Adult Plant Stripe Rust Resistance Derived from Cappelle-Desprez. Agronomy 2019, 9, 154. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030154
Maree GJ, Prins R, Boyd LA, Castelyn HD, Bender CM, Boshoff WH, Pretorius ZA. Assessing the Individual and Combined Effects of QTL for Adult Plant Stripe Rust Resistance Derived from Cappelle-Desprez. Agronomy. 2019; 9(3):154. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030154
Chicago/Turabian StyleMaree, Gerrie J, Renée Prins, Lesley A Boyd, Howard D Castelyn, Cornelia M Bender, Willem HP Boshoff, and Zacharias A Pretorius. 2019. "Assessing the Individual and Combined Effects of QTL for Adult Plant Stripe Rust Resistance Derived from Cappelle-Desprez" Agronomy 9, no. 3: 154. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030154