Resistance of Multiple Diploid and Tetraploid Perennial Ryegrass (Lolium perenne L.) Varieties to Three Projected Drought Scenarios for the UK in 2080


Abstract
:1. Introduction
2. Materials and Methods
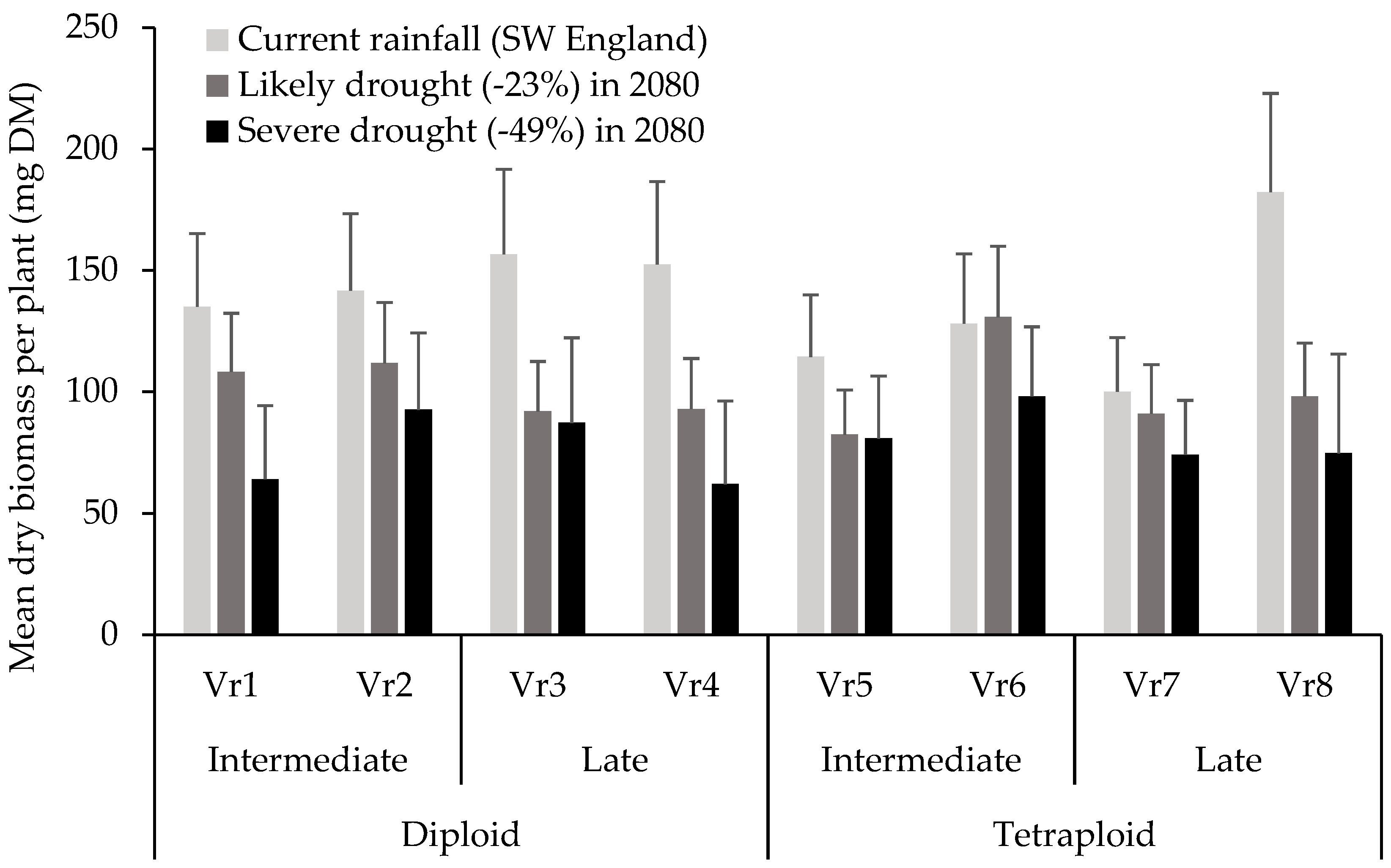
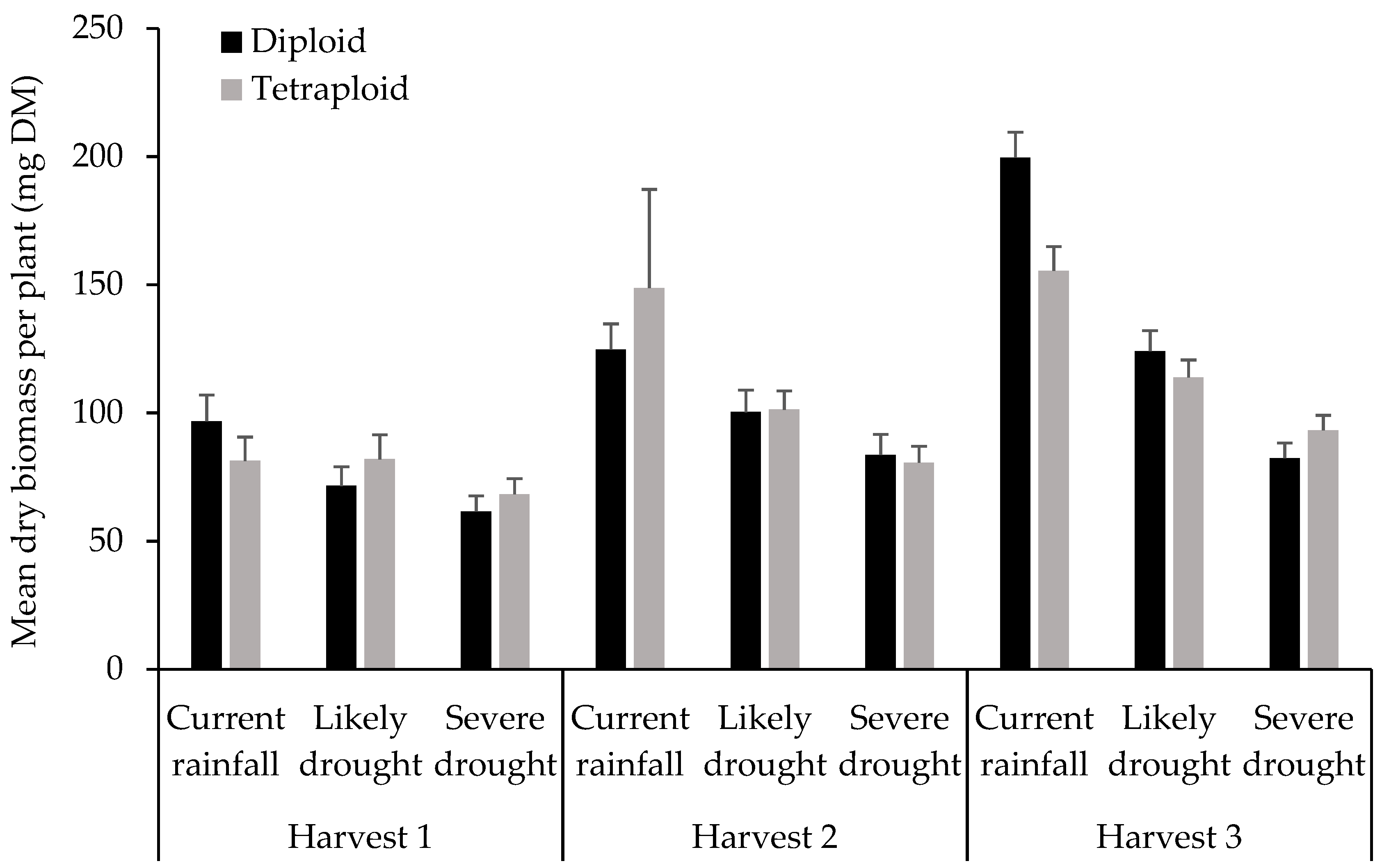
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| Cell dimensions (mm) | 38 × 38 |
| Cell area (mm2) | 1444 |
| Current rainfall 1 (mm) | 7 |
| Volume of rainfall per cell (mm3) | 10,108 |
| Cell number per tray | 40 |
| Total rainfall per tray (mm3) | 404,320 |
| Total volume per tray (ml)—current rainfall | 404 |
| Total volume per tray (ml)—likely drought | 303 |
| Total volume per tray (ml)—severe drought | 202 |
References
- IPCC. IPCC Fifth Assessment Report (AR5)—The Physical Science Basis. IPCC, 2013. Available online: https://www.ipcc.ch/assessment-report/ar5/ (accessed on 1 November 2017).
- Lee, M.A.; Davis, A.P.; Chagunda, M.G.G.; Manning, P. Forage quality declines with rising temperatures, with implications for livestock production and methane emissions. Biogeosciences 2017, 14, 1403–1417. [Google Scholar] [CrossRef]
- Tozer, K.N.; Carswell, K.; Griffiths, W.M.; Crush, J.R.; Cameron, C.A.; Chapman, D.F.; Popay, A.; King, W. Growth responses of diploid and tetraploid perennial ryegrass (Lolium perenne) to soil-moisture deficit, defoliation and a root-feeding invertebrate. Crop Pasture Sci. 2017, 68, 632–642. [Google Scholar] [CrossRef]
- Deru, J.; Schilder, H.; van der Schoot, J.R.; van Eekeren, N. Genetic differences in root mass of Lolium perenne varieties under field conditions. Euphytica 2014, 199, 223–232. [Google Scholar] [CrossRef]
- Ebrahimiyan, M.; Majidi, M.M.; Mirlohi, A. Genotypic variation and selection of traits related to forage yield in tall fescue under irrigated and drought stress environments. Grass Forage Sci. 2013, 68, 59–71. [Google Scholar] [CrossRef]
- Department for Environment, Food and Rural Affairs (DEFRA). Adapting to Climate Change UK Climate Projections; 2009. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/69257/pb13274-uk-climate-projections-090617.pdf (accessed on 1 November 2017).
- Gardiner, L.M.; Bone, R.; Kilgallen, N.M. Orchids and emonocot—Assembling research resources and facilitating collaborative taxonomy online. Lankesteriana 2013, 13, 33–37. [Google Scholar] [CrossRef]
- Met Office Met Office Integrated Data Archive System (MIDAS). Land and Marine Surface Stations Data (1853–current). NCAS Br. Atmos. Data Cent. 2017. Available online: http://catalogue.ceda.ac.uk/uuid/1bb479d3b1e38c339adb9c82c15579d8 (accessed on 1 November 2017).
- Brown, R.N.; Percivalle, C.; Narkiewicz, S.; DeCuollo, S. Relative rooting depths of native grasses and amenity grasses with potential for use on roadsides in New England. HortScience 2010, 45, 393–400. [Google Scholar] [CrossRef]
- Wedderburn, M.E.; Crush, J.R.; Pengelly, W.J.; Walcroft, J.L. Root growth patterns of perennial ryegrasses under well-watered and drought conditions. N. Z. J. Agric. Res. 2010, 53, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis HISTORICAL commentary NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- The R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3_900051_00_3. [Google Scholar]
- Sugiyama, S. Differentiation in competitive ability and cold tolerance between diploid and tetraploid cultivars in Lolium perenne. Euphytica 1998, 103, 55–59. [Google Scholar] [CrossRef]
- Cattivelli, L.; Rizza, F.; Badeck, F.W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Marè, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- Fry, E.L.; Evans, A.L.; Sturrock, C.J.; Bullock, J.M.; Bardgett, R.D. Root architecture governs plasticity in response to drought. Plant Soil 2018, 433, 189–200. [Google Scholar] [CrossRef]
- Lee, M.A. A global comparison of the nutritive values of forage plants grown in contrasting environments. J. Plant Res. 2018, 131, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Godfree, R.C.; Marshall, D.J.; Young, A.G.; Miller, C.H.; Mathews, S. Empirical evidence of fixed and homeostatic patterns of polyploid advantage in a keystone grass exposed to drought and heat stress. R. Soc. Open Sci. 2017, 4, 170934. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Live Biomass (g) | Dry Biomass (g) | DM (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| df | F | p | F | p | F | p | ||
| Harvest 1 | Ploidy | 1 | 0.40 | 0.54 | 0.01 | 0.90 | 0.001 | 0.97 |
| Rainfall | 2 | 38.80 | <0.001 | 4.40 | <0.05 | 30.20 | <0.001 | |
| Timing | 1 | 3.20 | 0.07 | 6.60 | <0.05 | 0.05 | 0.82 | |
| Harvest 2 | Ploidy | 1 | 0.40 | 0.50 | 0.30 | 0.60 | 2.40 | 0.10 |
| Rainfall | 2 | 34.50 | <0.001 | 5.10 | <0.01 | 7.00 | <0.01 | |
| Timing | 1 | 2.50 | 0.10 | 0.60 | 0.50 | 0.30 | 0.60 | |
| Harvest 3 | Ploidy | 1 | 6.40 | <0.05 | 4.80 | <0.05 | 2.50 | 0.10 |
| Rainfall | 2 | 62.00 | <0.001 | 63.10 | <0.001 | 7.40 | <0.001 | |
| Timing | 1 | 0.40 | 0.50 | 0.001 | 0.90 | 1.90 | 0.20 | |
| Stomatal Length (µm) | Stomatal Conductance (mmol m−2 s−1) | |||
|---|---|---|---|---|
| Mean | SE | Mean | SE | |
| Vr1 | 26.52 | 0.29 | 46.62 | 11.61 |
| Vr2 | 29.59 | 0.26 | 41.63 | 4.85 |
| Vr3 | 33.29 | 0.85 | 36.13 | 5.04 |
| Vr4 | 29.86 | 0.43 | 36.25 | 3.02 |
| Vr5 | 35.67 | 0.39 | 48.79 | 8.81 |
| Vr6 | 40.77 | 0.54 | 41.30 | 7.52 |
| Vr7 | 34.20 | 0.43 | 38.78 | 4.74 |
| Vr8 | 37.60 | 0.26 | 46.73 | 9.29 |
| Diploid | 30.67 | 0.40 | 40.15 | 3.30 |
| Tetraploid | 38.84 | 0.45 | 44.03 | 3.63 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.A.; Howard-Andrews, V.; Chester, M. Resistance of Multiple Diploid and Tetraploid Perennial Ryegrass (Lolium perenne L.) Varieties to Three Projected Drought Scenarios for the UK in 2080. Agronomy 2019, 9, 159. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030159
Lee MA, Howard-Andrews V, Chester M. Resistance of Multiple Diploid and Tetraploid Perennial Ryegrass (Lolium perenne L.) Varieties to Three Projected Drought Scenarios for the UK in 2080. Agronomy. 2019; 9(3):159. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030159
Chicago/Turabian StyleLee, Mark A., Victoria Howard-Andrews, and Michael Chester. 2019. "Resistance of Multiple Diploid and Tetraploid Perennial Ryegrass (Lolium perenne L.) Varieties to Three Projected Drought Scenarios for the UK in 2080" Agronomy 9, no. 3: 159. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030159