
Use of Principal Component Analysis to Combine Genetic Merit for Heat Stress and for Fat and Protein Yield in Spanish Autochthonous Dairy Goat Breeds

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Statistical Procedure
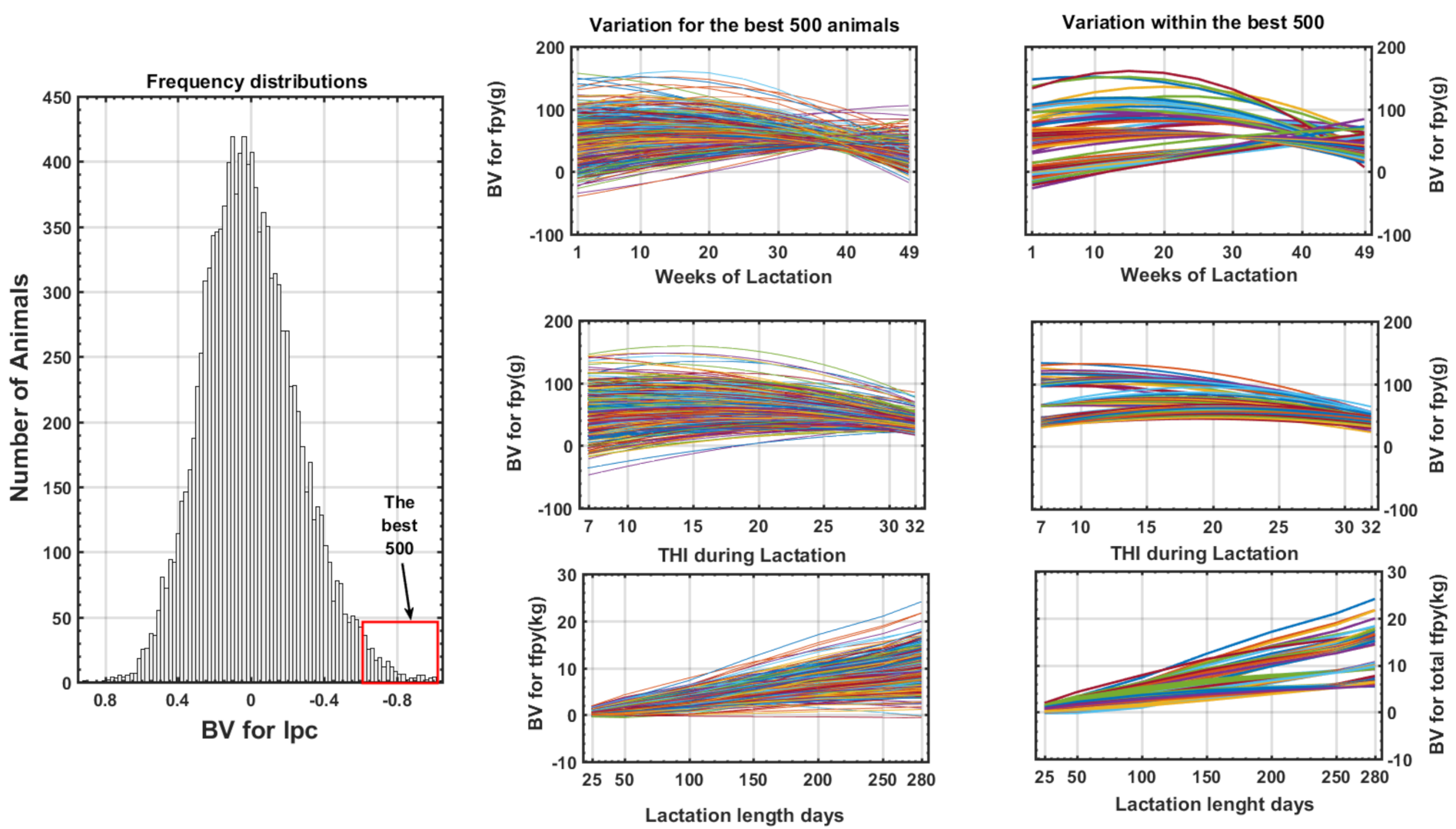
2.2. Breeding Value Estimation for THI and DIM and Development of a New Index
3. Results
3.1. Overall Results
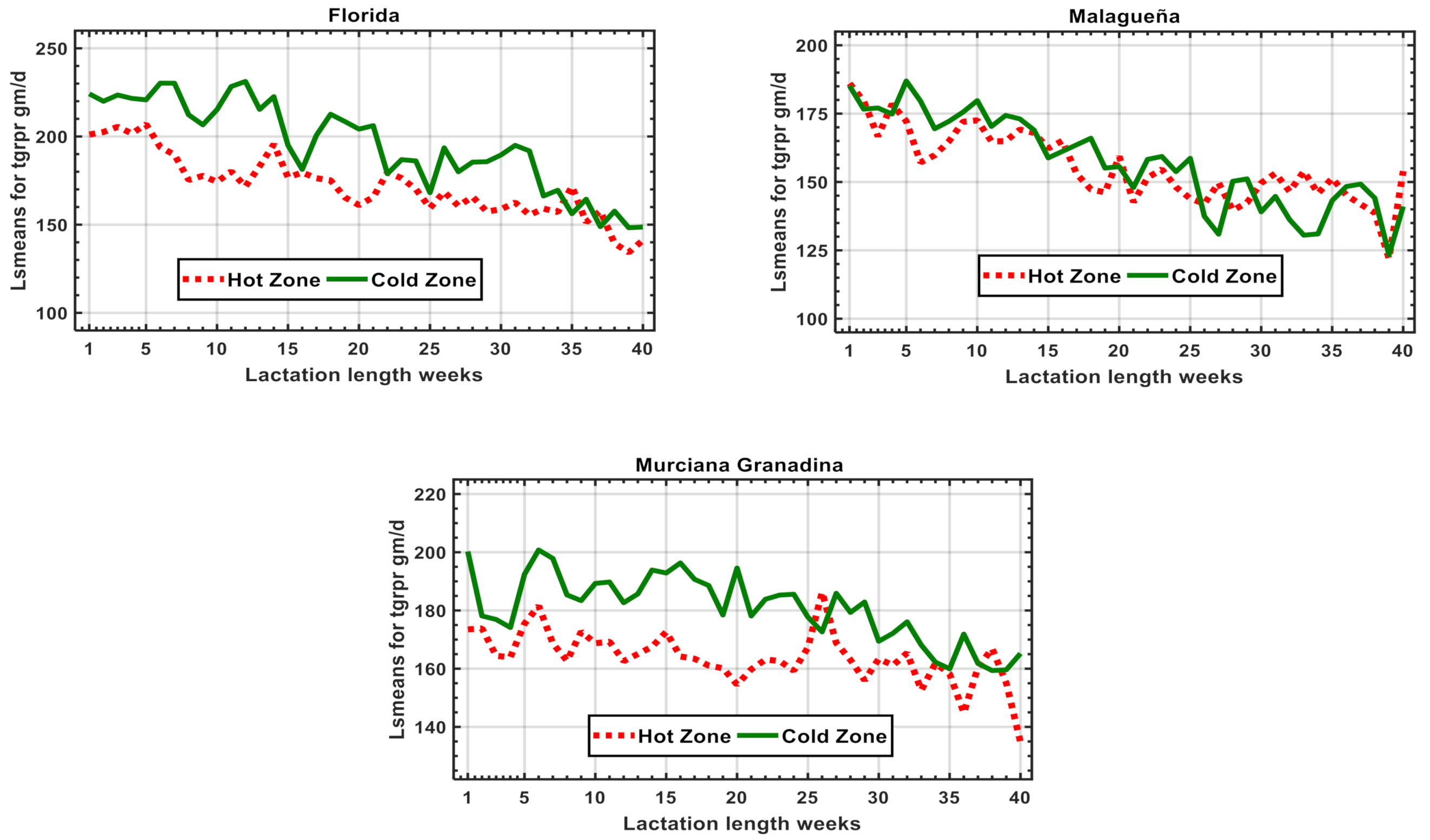
3.2. Results of Genetic Models along DIM. (Model 1)
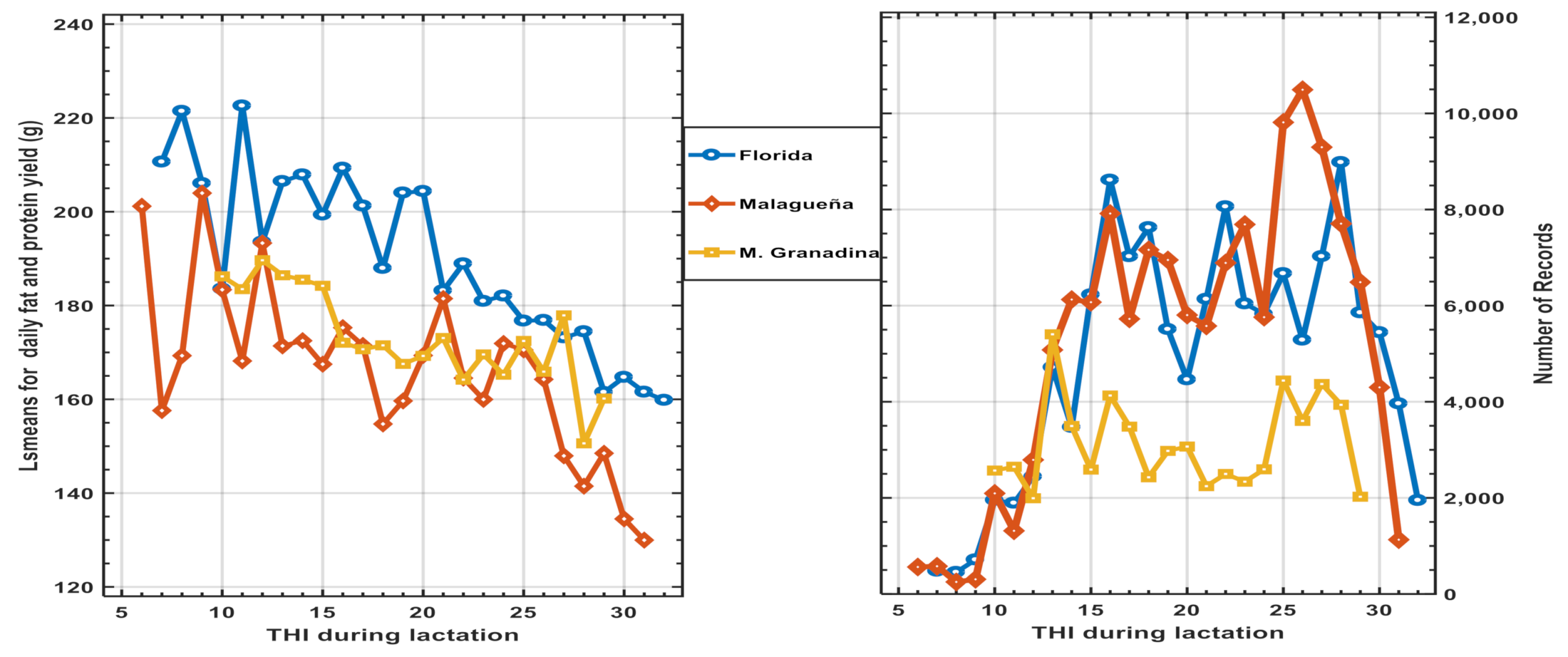
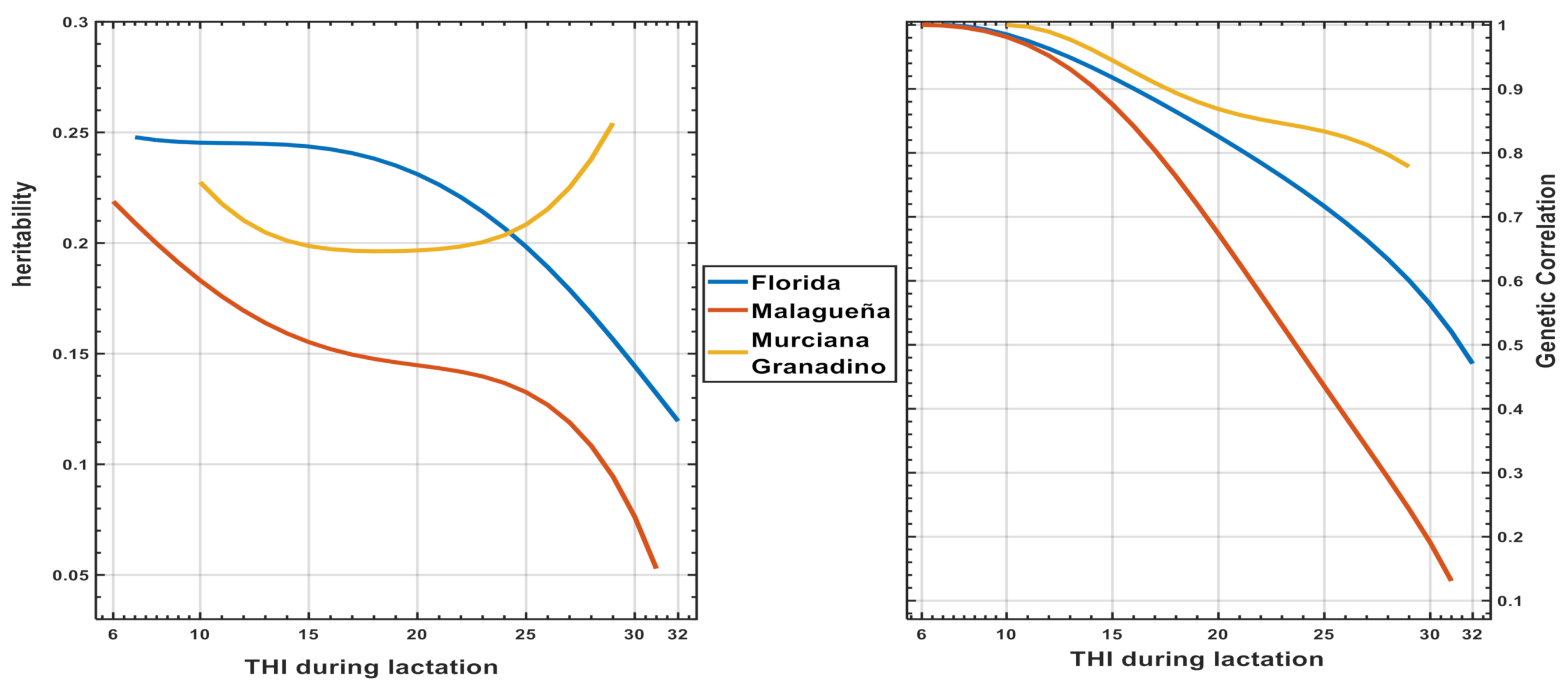
3.3. Results of Genetic Models along THI Effects (Model 2)
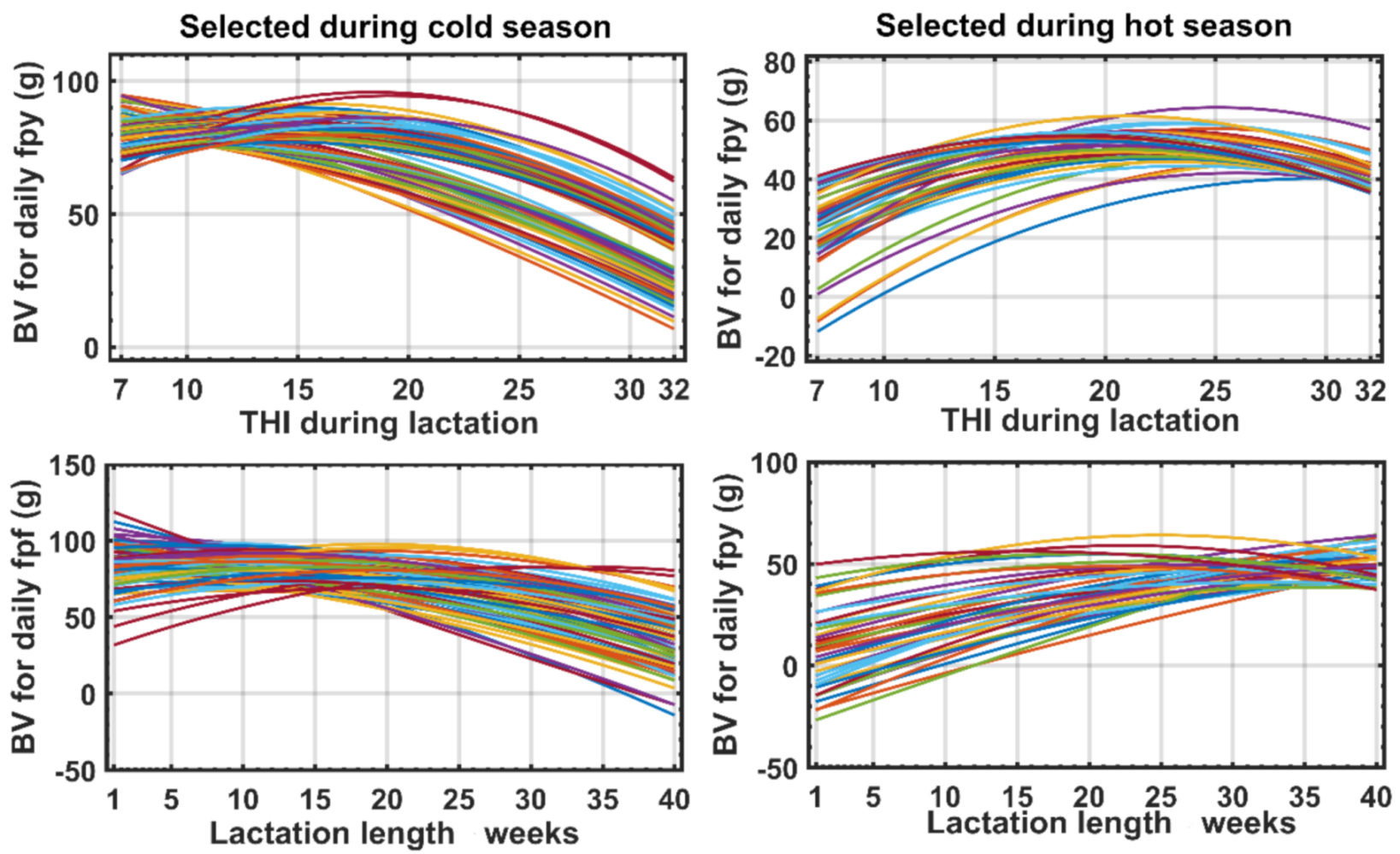
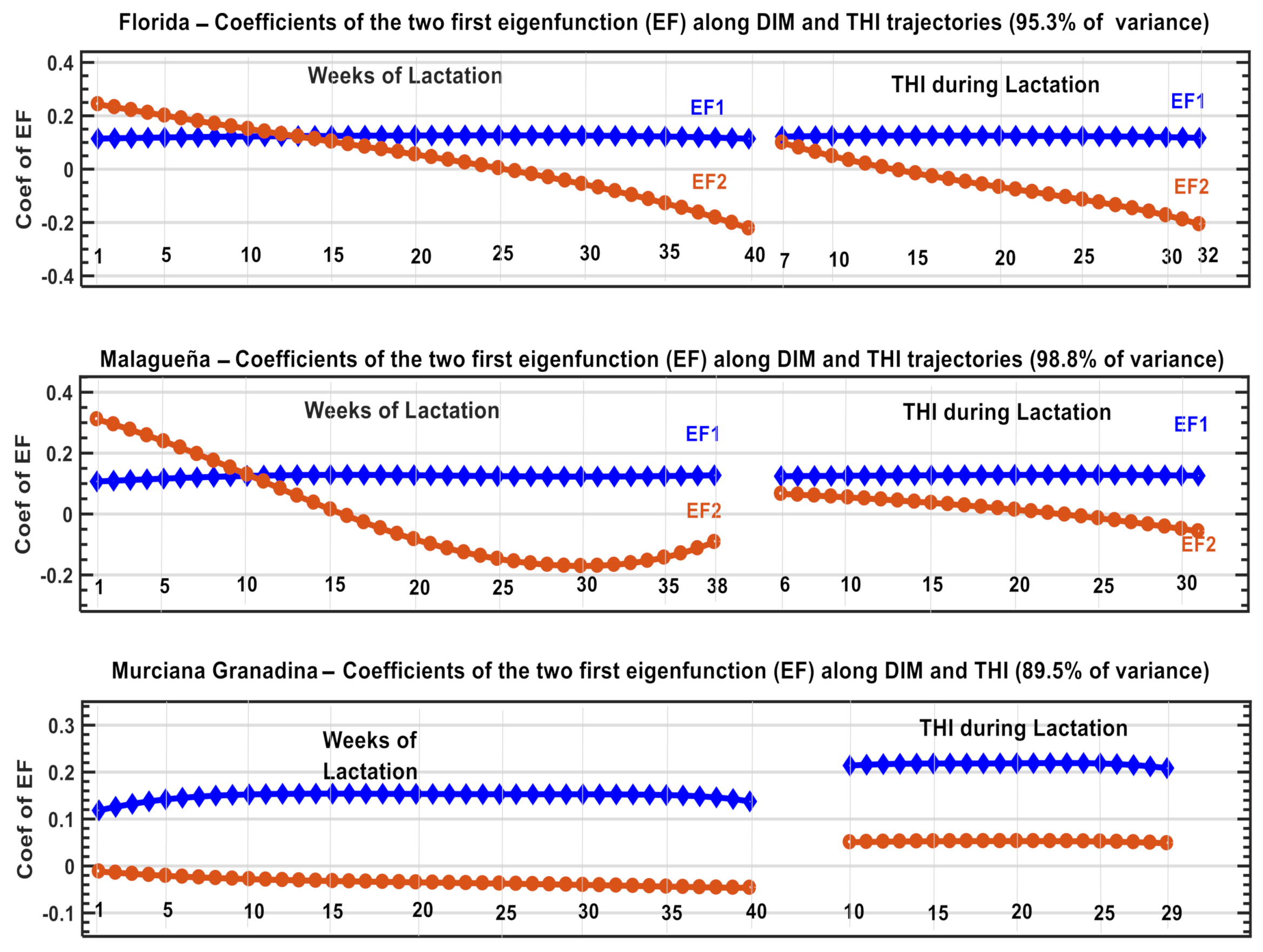
3.4. Generalizing the Response Forms of Fat Plus Protein Yield along the Trajectories of THI and DIM and Construction of the New Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
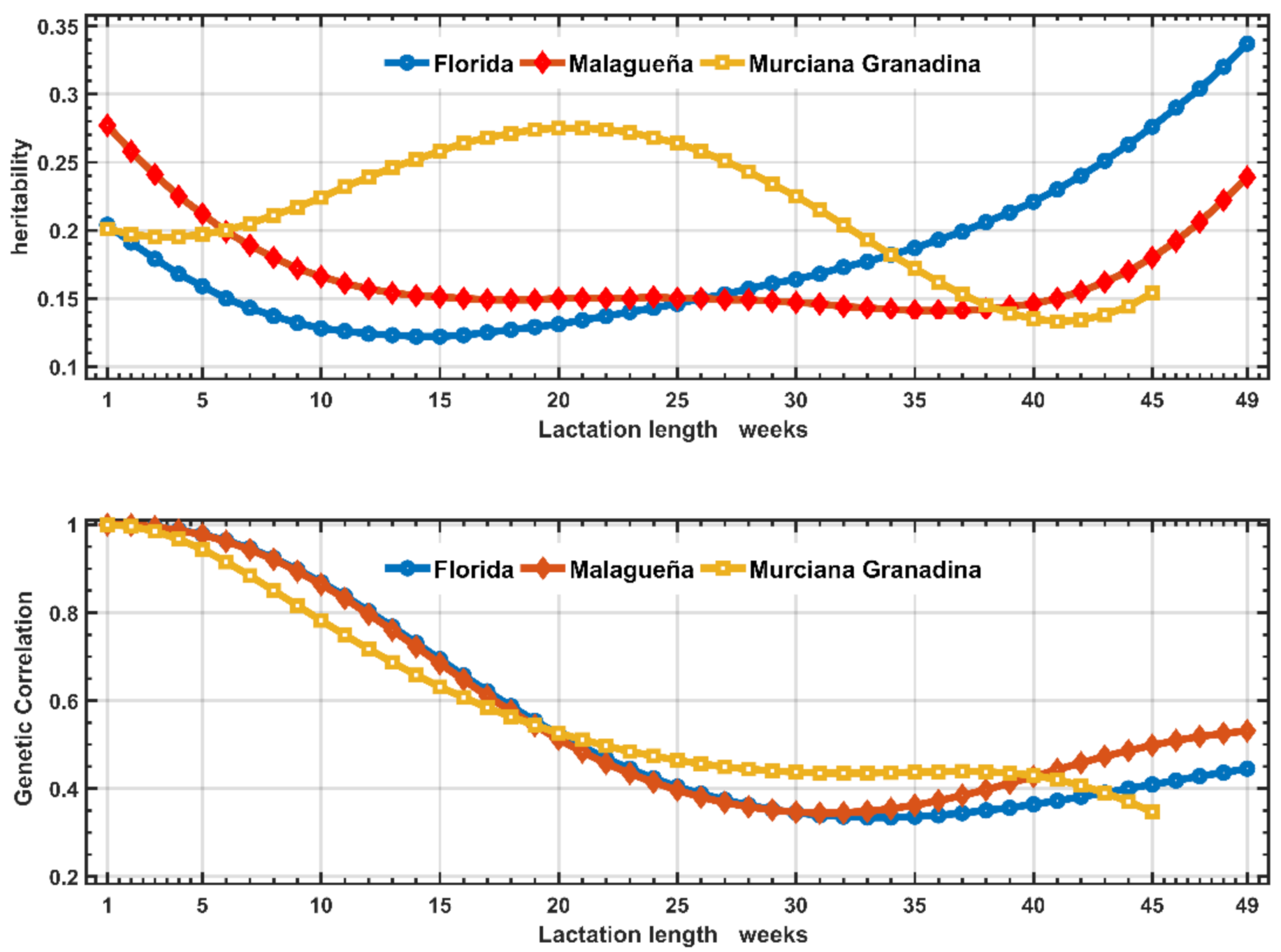
Heritability, Genetic Correlation and Breeding Values
References
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Amponsah, R.; Chauhan, S.S.; Leury, B.J.; Cheng, L.; Cullen, B.; Clarke, I.J.; Dunshea, F.R. Review Genetic Selection for Thermotolerance in Ruminants. Animals 2019, 9, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purohit, P.; Gupta, J.; Chaudhri, J.; Bhatt, T.; Pawar, M.; Srivastava, A.; Patel, M. Effect of Heat Stress on Production and Reproduction Potential of Dairy Animals vis-à-vis Buffaloes. Int. J. Livest. Res. 2020, 10, 1–23. [Google Scholar] [CrossRef]
- Menéndez-Buxadera, A.; Molina, A.; Arrebola, F.; Clemente, I.; Serradilla, J. Genetic variation of adaptation to heat stress in two Spanish dairy goat breeds. J. Anim. Breed. Genet. 2012, 129, 306–315. [Google Scholar] [CrossRef]
- Menéndez-Buxadera, A.; Serradilla, J.M.; Arrebola, F.; Clemente, I.; Castro, J.A.; Osorio, J.; Torres, R.; Molina, A. Genetic variation for tolerance to heat stress in dairy small ruminants: Results obtained in Spain. In Proceedings of the 8th International Seminar Technology Creation and Transfer in Small Ruminants: Roles of Research, Development Services and Farmer Associations, Tangier, Morocco, 11–13 June 2013; pp. 67–73. [Google Scholar]
- Menéndez-Buxadera, A.; Serradilla, J.; Molina, A. Genetic variability for heat stress sensitivity in Merino de Grazalema sheep. Small Rumin. Res. 2014, 121, 207–214. [Google Scholar] [CrossRef]
- Carabaño, M.J.; Ramón, M.; Menéndez-Buxadera, A.; Molina, A.; Díaz, C. Selecting for heat tolerance. Anim. Front. 2019, 9, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravagnolo, O.; Misztal, I. Genetic Component of Heat Stress in Dairy Cattle, Parameter Estimation. J. Dairy Sci. 2000, 83, 2126–2130. [Google Scholar] [CrossRef]
- Carabaño, M.J.; Ramón, M.; Díaz, C.; Molina, A.; Pérez-Guzmán, M.D.; Serradilla, J.M. Breeding and Genet-ics Symposium: Breeding for resilience to heat stress effects in dairy ruminants. A comprehensive review. J. Anim. Sci. 2017, 95, 1813–1826. [Google Scholar]
- Misztal, I.; Bohmanova, J.; Freitas, M.; Tsuruta, S.; Norman, H.D.; Lawlor, T.J. Issues in genetic evaluation of dairy cattle for heat tolerance. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Brasil, 13–18 August 2006. [Google Scholar]
- De Jong, G.; Bijma, P. Selection and phenotypic plasticity in evolutionary biology and animal breeding. Livest. Prod. Sci. 2002, 78, 195–214. [Google Scholar] [CrossRef]
- Bohlouli, M.; Alijani, S.; Naderi, S.; Yin, T.; König, S. Prediction accuracies and genetic parameters for test-day traits from genomic and pedigree-based random regression models with or without heat stress interactions. J. Dairy Sci. 2019, 102, 488–502. [Google Scholar] [CrossRef] [Green Version]
- Santana, M.L., Jr.; Bignardi, A.B.; Pereira, R.J.; Stefani, G.; el Faro, L. Genetics of heat tolerance for milk yield and quality in Holsteins. Animal 2016, 11, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hair, J., Jr.; Black, W.; Babin, B.; Anderson, R. Multivariate Data Analysis, 7th ed.; Prentice Hall: Upper Saddle River, UK, 2009. [Google Scholar]
- Togashi, K.; Lin, C.Y. Selection for Milk Production and Persistency Using Eigenvectors of the Random Regression Coefficient Matrix. J. Dairy Sci. 2006, 89, 4866–4873. [Google Scholar] [CrossRef]
- Venturini, G.C.; Savegnago, R.P.; Nunes, B.N.; Ledur, M.C.; Schmidt, G.S.; El Faro, L.; Munari, D.P. Genetic parameters and principal component analysis for egg production from White Leghorn hens. Poult. Sci. 2013, 92, 2283–2289. [Google Scholar] [CrossRef] [PubMed]
- Boligon, A.A.; Vicente, S.; Vaz, R.Z.; Campos, G.S.; Souza, F.R.P.; Carvalheiro, R.; Albuquerque, L.G. Prin-cipal components analysis of breeding values for growth and reproductive traits and genetic association with adult size in beef cattle. J. Anim. Sci. 2016, 94, 5014–5022. [Google Scholar] [CrossRef] [PubMed]
- Macciotta, N.P.P.; Biffani, S.; Bernabucci, U.; Lacetera, N.; Vitali, A.; Ajmone-Marsan, P.; Nardone, A. Derivation and genome-wide association study of a principal component-based measure of heat tolerance in dairy cattle. J. Dairy Sci. 2017, 100, 4683–4697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finocchiaro, R.; Van Kaam, J.; Portolano, B.; Misztal, I. Effect of Heat Stress on Production of Mediterranean Dairy Sheep. J. Dairy Sci. 2005, 88, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, A.R.; Gogel, R.B.J.; Cullis, B.R.; Thompson, R. Asreml User Guide Release 3.0; VSN International Ltd.: Hemel Hempstead, UK, 2009. [Google Scholar]
- Jamrozik, J.; Schaeffer, L.R. Estimates of Genetic Parameters for a Test Day Model with Random Regressions for Yield Traits of First Lactation Holsteins. J. Dairy Sci. 1997, 80, 762–770. [Google Scholar] [CrossRef]
- MATLAB and Statistics Toolbox Release; The Math Works, Inc.: Natick, MA, USA, 2019.
- Linares, J.M. La crisis del clima. In Evidencias del Cambio Climático en España; Greenpeace España: Madrid, Spain, 2009; p. 75. [Google Scholar]
- Carabaño, M.J.; Ramón, M.; Díaz, C.; Molina, A.; Pérez-Guzmán, M.D.; Serradilla, J.M. Breeding for resilience to heat stress effects. A comparison across dairy ruminant species. J. Anim. Sci. 2016, 94 (Suppl. 5), 195. [Google Scholar]
- Cheruiyot, E.; Nguyen, T.; Haile-Mariam, M.; Cocks, B.; Abdelsayed, M.; Pryce, J. Genotype-by-environment (temperature-humidity) interaction of milk production traits in Australian Holstein cattle. J. Dairy Sci. 2020, 103, 2460–2476. [Google Scholar] [CrossRef]
- Serradilla, J.M.; Carabaño, M.J.; Ramón, M.; Molina, A.; Diaz, C.; Menéndez-Buxadera, A. Characterisation of Goats’ Response to Heat Stress: Tools to Improve Heat Tolerance. Goat Sci. 2018, 15, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Thuy, T.T.N.; Bowman, P.J.; Haile-Mariam, M.; Pryce, J.E.; Hayes, B.J. Genomic selection for toler-ance to heat stress in Australian dairy cattle. J. Dairy Sci. 2016, 99, 2849–2862. [Google Scholar]
- Brügemann, K.; Gernand, E.; Von Borstel, U.; König, S. Genetic analyses of protein yield in dairy cows applying random regression models with time-dependent and temperature × humidity-dependent covariates. J. Dairy Sci. 2011, 94, 4129–4139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravagnolo, O.; Misztal, I.; Hoogenboom, G. Genetic Component of Heat Stress in Dairy Cattle, Development of Heat Index Function. J. Dairy Sci. 2000, 83, 2120–2125. [Google Scholar] [CrossRef]
- Stinchcombe, J.R.; Kirkpatrick, M. Genetics and evolution of function valued traits: Understanding environmentally responsive phenotypes. Trends Ecol. Evol. 2012, 27, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Togashi, K.; Lin, C.Y. Improvement of lactation milk and persistency using the eigenvectors of the genetic covariance matrix between lactation stages. Livest. Sci. 2007, 110, 64–72. [Google Scholar] [CrossRef]
- Hammami, H.; Vandenplas, J.; Carabaño, M.J.; Logar, B.; Bormann, J.; Bertozzi, C.; Gengler, N. Strategies to Combine Novel Traits across Countries: Example of Heat Stress; Interbull Bulletin No. 48; Interbull Center: Berlin, Germany, 2014; pp. 79–84. [Google Scholar]
- Carabaño, M.; Logar, B.; Bormann, J.; Minet, J.; Vanrobays, M.-L.; Díaz, C.; Tychon, B.; Gengler, N.; Hammami, H. Modeling heat stress under different environmental conditions. J. Dairy Sci. 2016, 99, 3798–3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, M.; Meyer, K. Direct Estimation of Genetic Principal Components. Simplified Analysis of Complex Phenotypes. Genetics 2004, 168, 2295–2306. [Google Scholar] [CrossRef] [Green Version]
- Druet, T.; Jaffrézic, F.; Ducroc, V. Estimation of genetic parameters for test day records of dairy traits in the first three lactations. Genet. Sel. Evol. 2005, 37, 257–271. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeds | |||
|---|---|---|---|
| Florida | Malagueña | Murciano-Granadina | |
| Number of records | 126,825 | 141,856 | 62,834 |
| Number of animals | 11,244 | 12,215 | 5162 |
| Number of dams | 1999 | 2650 | 2198 |
| Number of sires | 233 | 158 | 215 |
| Number of flocks | 20 | 17 | 20 |
| Animals in pedigree | 12,268 | 14,075 | 6037 |
| THI 1 level | 21.4 (5.9) 2 | 21.4 (5.7) | 19.6 (5.8) |
| No of recorded flock-date | 725 | 637 | 474 |
| Average days in milk | 122.6 (72.8) | 131.3 (77.3) | 130.1 (76.7) |
| Daily fat and protein yield (g) | 190.7 (83.5) | 162.1 (70.9) | 176.4 (80.5) |
| Florida | Malagueña | Murciana-Granadina | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Cold | Neutral | Hot | Cold | Neutral | Hot | Cold | Neutral | Hot | |
| %Time of the year | 24.4 | 45.2 | 30.4 | 24.3 | 44.6 | 30.9 | 36.3 | 34.3 | 29.2 |
| LSmeans in g | 194.5(9.8) | 201.9(3.8) | 170.8(4.6) | 167.2(0.6) | 163.8(0.4) | 159.4(0.6) | 176.7(0.7) | 187.8(0.73) | 180.9(0.8) |
| Regr-cold ** | Y280 = 240.9 − 0.38 + 0.0004, R2 = 75.9% | Y280 = 196.1 − 0.36 + 0.00008, R2 = 63.1% | Y280 = 194.9 − 0.09, R2 = 20.0% | ||||||
| Regr-hot *** | Y280 = 209.5 − 0.24, R2 = 83.2% | Y280 = 186.0 − 0.25 + 0.00008, R2 = 67.0% | Y280 = 179.9 − 0.06, R2 = 17.1% | ||||||
| Tot fpy **** hot | 47.3 kg | 42.1 kg | 46.9 kg | ||||||
| Tot fpy **** cold | 53.0 kg | 44.3 kg | 50.2 kg | ||||||
| Differences | −5.8 kg | −2.2 kg | −3.3 kg | ||||||
| Equivalent in days | 30 | 13.5 | 18.8 | ||||||
| Florida | Malagueña | Murciano−Granadina | |||||||
|---|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| Intercept | 0.990 | 0.126 | −0.052 | 0.9814 | 0.1905 | 0.0245 | −0.974 | 0.131 | 0.183 |
| Slope | 0.133 | −0.975 | 0.178 | −0.1911 | 0.9564 | 0.2210 | 0.111 | 0.987 | −0.117 |
| Quadratic | 0.028 | 0.183 | 0.983 | 0.0187 | −0.2216 | 0.9750 | 0.196 | 0.094 | 0.976 |
| Eigenvalues, % | 78.5 | 14.7 | 6.8 | 74.9 | 15.5 | 9.6 | 83.6 | 9.5 | 6.9 |
| Cumulative Variance explained | 93.2 | 100 | 89.7 | 100 | 93.1 | 100 | |||
| Florida | Malagueña | Murciano-Granadina | ||||||
|---|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC1 | PC2 | PC3 | |
| Intercept | −0.9875 | 0.1187 | 0.1037 | 0.9853 | 0.1711 | 0.9991 | 0.0212 | 0.0356 |
| Slope | 0.1356 | 0.9752 | 0.1750 | −0.1711 | 0.9853 | −0.0265 | 0.9874 | 0.1562 |
| Quadratic | 0.0804 | −0.1869 | 0.9791 | na | na | 0.0318 | 0.1571 | −0.9871 |
| Eigenvalues % | 92.70 | 7.20 | 0.10 | 92.77 | 7.23 | 93.30 | 4.85 | 1.85 |
| Cumulative Variance explained | 99.9 | 100 | 100 | 98.15 | 100 | |||
| Florida | Malagueña | Murciano-Granadina | |||||||
|---|---|---|---|---|---|---|---|---|---|
| aoTHI | asTHI | aqTHI | aoTHI | asTHI | aqTHI | aoTHI | asTHI | aqTHI | |
| aoDIM | 0.880 | −0.418 | −0.252 | 0.968 | −0.389 | X | 0.950 | −0.489 | −0.527 |
| asDIM | −0.497 | 0.351 | 0.021 | −0.483 | 0.401 | X | −0.182 | 0.155 | 0.177 |
| aqDIM | −0.905 | 0.403 | 0.292 | 0.068 | −0.302 | X | 0.225 | −0.153 | −0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menéndez-Buxadera, A.; Muñoz-Mejías, E.; Sánchez, M.; Serradilla, J.M.; Molina, A. Use of Principal Component Analysis to Combine Genetic Merit for Heat Stress and for Fat and Protein Yield in Spanish Autochthonous Dairy Goat Breeds. Animals 2021, 11, 736. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030736
Menéndez-Buxadera A, Muñoz-Mejías E, Sánchez M, Serradilla JM, Molina A. Use of Principal Component Analysis to Combine Genetic Merit for Heat Stress and for Fat and Protein Yield in Spanish Autochthonous Dairy Goat Breeds. Animals. 2021; 11(3):736. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030736
Chicago/Turabian StyleMenéndez-Buxadera, Alberto, Eva Muñoz-Mejías, Manuel Sánchez, Juan Manuel Serradilla, and Antonio Molina. 2021. "Use of Principal Component Analysis to Combine Genetic Merit for Heat Stress and for Fat and Protein Yield in Spanish Autochthonous Dairy Goat Breeds" Animals 11, no. 3: 736. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11030736