
Effect of a Black Soldier Fly Ingredient on the Growth Performance and Disease Resistance of Juvenile Pacific White Shrimp (Litopenaeus vannamei)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Shrimp Rearing and Feeding Trial
2.3. Growth Performance Trial
2.4. Salinity Stress Test
2.5. Disease Challenge Tests
2.6. Data Analyses
3. Results
3.1. Growth Performance
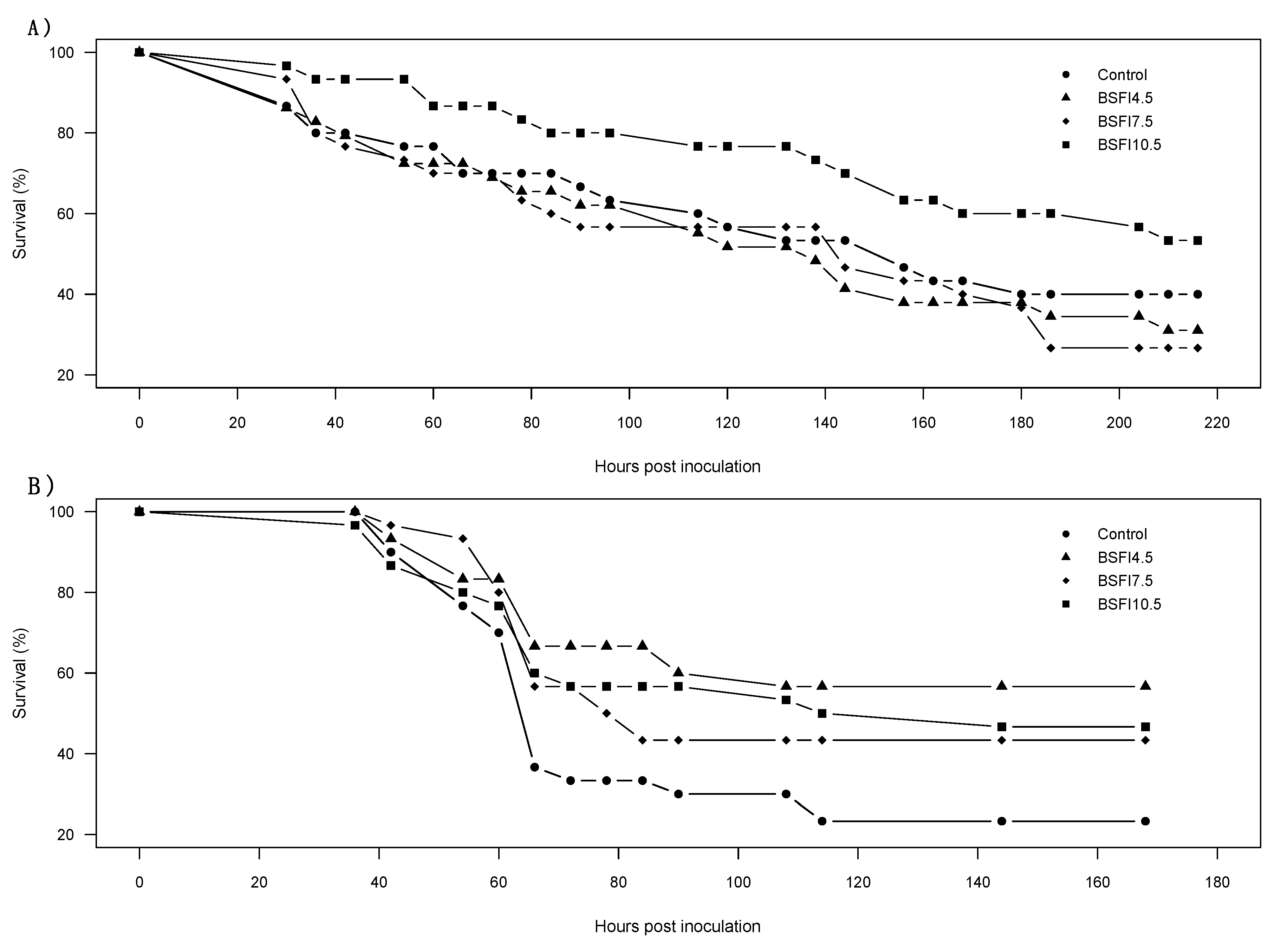
3.2. Salinity and Disease Challenge Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrows, F.; Gaylord, T.; Stone, D.; Smith, C. Effect of protein source and nutrient density on growth efficiency, histology and plasma amino acid concentration of rainbow trout (Oncorhynchus mykiss Walbaum). Aquac. Res. 2007, 38, 1747–1758. [Google Scholar]
- Gatlin, D.; Barrows, F.; Brown, P.; Dabrowski, K.; Gaylord, T.; Hardy, R.; Herman, E.; Hu, G.; Krogdahl, A.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing fishmeal and fish oil in industrial aquafeed. Feed and feeding practices in aquaculture. In Feed and Feeding Practices in Aquaculture; Davis, A., Ed.; Woodhead Publishing Limited: Waltham, MA, USA, 2015; pp. 203–233. [Google Scholar]
- Gai, F.; Gasco, L.; Daprà, F.; Palmegiano, G.B.; Sicuro, B. Enzymatic and histological evaluations of gut and liver in rainbow trout Oncorhynchus mykiss, fed with rice protein concentrate based diets. J. World Aquac. Soc. 2012, 43, 218–229. [Google Scholar] [CrossRef]
- Glencross, B.; Booth, M.; Allan, G.L. A feed is only as good as its ingredients—A review of ingredient evaluation strategies for aquaculture feeds. Aquac. Nutr. 2007, 13, 17–34. [Google Scholar]
- Hardy, R.W. Utilization of plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Krogdahl, A.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar]
- Malcorps, W.; Kok, B.; van‘t Land, M.; Fritz, M.; van Doren, D.; Servin, K.; van der Heijden, P.; Palmer, R.; Auchterlonie, N.A.; Rietkerk, M. The sustainability conundrum of fishmeal substitution by plant ingredients in shrimp feeds. Sustainability 2019, 11, 1212. [Google Scholar]
- Moutinho, S.; Martínez-Llorens, S.; Tomás-Vidal, A.; Jover-Cerdá, M.; Oliva-Teles, A.; Peres, H. Meat and bone meal as partial replacement for fish meal in diets for gilthead seabream (Sparus aurata) juveniles: Growth, feed efficiency, amino acid utilization, and economic efficiency. Aquaculture 2016, 468, 271–277. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2020—Sustainability in Action; The Food and Agricultural Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- European Commission. Commission regulation (EU) 2017/893 of 24 May 2017 amending annexes I and IV to regulation (EC) No 999/2001 of the European Parliament and of the council and annexes X, XIV and XV to commission regulation (EU) No 142/2011 as regards the provisions on processed animal protein. Off. J. Eur. Union 2017, 138, 92–116. [Google Scholar]
- Magalhães, R.; Sánchez-Lópezb, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar]
- Stadtlander, T.; Stamer, A.; Buser, A.; Wohlfahrt, J.; Leiber, F.; Sandrock, C. Hermetia illucens meal as fish meal replacement for rainbow trout on farm. J. Insects Food Feed 2017, 3, 165–175. [Google Scholar]
- Motte, C.; Rios, A.; Lefebvre, T.; Do, H.; Henry, M.; Jintasataporn, O. Replacing fish meal with defatted insect meal (Yellow mealworm Tenebrio molitor) improves the growth and immunity of pacific white shrimp (Litopenaeus vannamei). Animals 2019, 9, E258. [Google Scholar] [CrossRef] [Green Version]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Anim. Sci. Biotechnol. 2017, 8, 57. [Google Scholar]
- Cummins, V.C.; Rawles, S.D.; Thompson, K.R.; Velasquez, A.; Kobayashi, Y.; Hager, J.; Webster, C.D. Evaluation of black soldier fly (Hermetia illucens) larvae meal as partial or total replacement of marine fish meal in practical diets for Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2017, 473, 337–344. [Google Scholar]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar]
- Sánchez-Muros, M.; Barroso, F.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar]
- Mousavi, S.; Zahedinezhad, S.; Loh, J.Y. A review on insect meals in aquaculture: The immunomodulatory and physiological effects. Int. Aquatic Res. 2020, 12, 100–115. [Google Scholar]
- El-Hack, A.; Mohamed, E.; Shafi, M.E.; Alghamdi, W.Y.; Abdelnour, S.A.; Shehata, A.M.; Noreldin, A.E.; Ashour, E.A.; Swelum, A.A.; Al-Sagan, A.A. Black soldier fly (Hermetia illucens) meal as a promising feed ingredient for poultry: A Comprehensive review. Agriculture 2020, 10, 339. [Google Scholar]
- Wang, Y.-S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed–a review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar]
- Barroso, F.; de Haro, C.; Sánchez-Muros, M.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422, 193–201. [Google Scholar]
- Wang, S.H.; Chen, J.C. The protective effect of chitin and chitosan against Vibrio alginolyticus in white shrimp Litopenaeus vannamei. Fish. Shellfish Immunol. 2005, 19, 191–204. [Google Scholar]
- Esteban, M.A.; Mulero, V.; Cuesta, A.; Ortuno, J.; Meseguer, J. Effects of injecting chitin particles on the innate immune response of gilthead seabream (Sparus aurata L.). Fish. Shellfish Immunol. 2000, 10, 543–544. [Google Scholar]
- Cock, J.; Salazar, M.; Rye, M. Strategies for managing diseases in non-native shrimp populations. Rev. Aquac. 2017, 9, 211–226. [Google Scholar]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar]
- Rahimnejad, S.; Hu, S.; Song, K.; Wang, L.; Lu, K.; Wu, R.; Zhang, C. Replacement of fish meal with defatted silkworm (Bombyx mori L.) pupae meal in diets for Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2019, 510, 150–159. [Google Scholar]
- Krogdahl, Å.; Bakke-McKellep, A.; Baeverfjord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar]
- Palacios, E.; Perez-Rostro, C.; Ramirez, J.; Ibarra, A.; Racotta, I. Reproductive exhaustion in shrimp (Penaeus vannamei) reflected in larval biochemical composition, survival and growth. Aquaculture 1999, 171, 309–321. [Google Scholar]
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests for Aquatic Animals; Office International des Epizooties: Paris, France, 2016; p. 589. [Google Scholar]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar]
- Escobedo-Bonilla, C.M.; Wille, M.; Sanz, V.A.; Sorgeloos, P.; Pensaert, M.; Nauwynck, H. In vivo titration of white spot syndrome virus (WSSV) in specific pathogen-free Litopenaeus vannamei by intramuscular and oral routes. Dis. Aquat. Org. 2005, 66, 163–170. [Google Scholar]
- Jiravanichpaisal, P.; Bangyeekhun, E.; Söderhäll, K.; Söderhäll, I. Experimental infection of white spot syndrome virus in freshwater crayfish Pacifastacus leniusculus. Dis. Aquatic Org. 2001, 47, 151–157. [Google Scholar]
- Van Thuong, K.; Van Tuan, V.; Li, W.; Sorgeloos, P.; Bossier, P.; Nauwynck, H. Per os infectivity of white spot syndrome virus (WSSV) in white-legged shrimp (Litopenaeus vannamei) and role of peritrophic membrane. Vet. Res. 2016, 47, 39. [Google Scholar]
- Choi, I.H.; Kim, J.M.; Kim, N.J.; Kim, J.D.; Park, C.; Park, J.H.; Chung, T.H. Replacing fish meal by mealworm (Tenebrio molitor) on the growth performance and immunologic responses of white shrimp (Litopenaeus vannamei). Acta Sci. Anim. Sci. 2018, 40. [Google Scholar] [CrossRef] [Green Version]
- Panini, R.L.; Freitas, L.E.L.; Guimarães, A.M.; Rios, C.; da Silva, M.F.O.; Vieira, F.N.; Amboni, R.D. Potential use of mealworms as an alternative protein source for Pacific white shrimp: Digestibility and performance. Aquaculture 2017, 473, 115–120. [Google Scholar]
- Karapanagiotidis, I.; Daskalopoulou, E.; Vogiatzis, I.; Rumbos, C.; Mente, E.; Athanassiou, C. Substitution of fishmeal by fly Hermetia illucens prepupae meal in the diet of gilthead seabream (Sparus aurata). HydroMedit 2014, 110–114. [Google Scholar]
- Lock, E.; Arsiwalla, T.; Waagbo, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar]
- Sealey, W.; Gaylord, T.; Barrows, F.; Tomberlin, J.; McGuire, M.; Ross, C.; St-Hilaire, S. Sensory analysis of rainbow trout, Oncorhynchus mykiss, fed enriched black soldier fly prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar]
- Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T.; Miura, C.; Miura, T. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pargus major). Animals 2019, 9, 100. [Google Scholar]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; Gasco, L. Tenebrio molitor meal in rainbow trout (Oncorhynchus mykiss) diets: Effects on animal performance, nutrient digestibility and chemical composition of fillets. Italian J. Anim. Sci. 2015, 14, 670–676. [Google Scholar]
- Gasco, L.; Henry, M.; Piccolo, G.; Marono, S.; Gai, F.; Renna, M.; Chatzifotis, S. Tenebrio molitor meal in diets for European sea bass (Dicentrarchus labrax L.) juveniles: Growth performance, whole body composition and in vivo apparent digestibility. Anim. Feed Sci. Technol. 2016, 220, 34–45. [Google Scholar]
- Rahimnejad, S.; Yuan, X.; Wang, L.; Lu, K.; Song, K.; Zhang, C. Chitooligosaccharide supplementation in low-fish meal diets for Pacific white shrimp (Litopenaeus vannamei): Effects on growth, innate immunity, gut histology, and immune-related genes expression. Fish. Shellfish Immunol. 2018, 80, 405–415. [Google Scholar]
- Liu, Y.; Wang, W.-N.; Wang, A.-L.; Wang, J.-M.; Sun, R.-Y. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei (Boone, 1931) exposed to acute salinity changes. Aquaculture 2007, 265, 351–358. [Google Scholar]
- Setiarto, A.; Augusto Strüssmann, C.; Takashima, F.; Watanabe, S.; Yokota, M. Short-term responses of adult kuruma shrimp Marsupenaeus japonicus (Bate) to environmental salinity: Osmotic regulation, oxygen consumption and ammonia excretion. Aquac. Res. 2004, 35, 669–677. [Google Scholar]
- Joseph, A.; Philip, R. Acute salinity stress alters the haemolymph metabolic profile of Penaeus monodon and reduces immunocompetence to white spot syndrome virus infection. Aquaculture 2007, 272, 87–97. [Google Scholar]
- Lieberman, S.; Enig, M.G.; Preuss, H.G. A review of monolaurin and lauric acid: Natural virucidal and bactericidal agents. Altern. Complementary Ther. 2006, 12, 310–314. [Google Scholar]
- Gasco, L.; Finke, M.; Van Huis, A. Can Diets Containing Insects Promote Animal Health? Wageningen Academic Publishers: Wageningen, The Netherlands, 2018. [Google Scholar]
- Destoumieux, D.; Munoz, M.; Bulet, P.; Bachere, E. Penaeidins, a family of antimicrobial peptides from penaeid shrimp (Crustacea, Decapoda). Cell. Molec. Life Sci. 2000, 57, 1260–1271. [Google Scholar]
- Choi, S.; Ingale, S.; Kim, J.; Park, Y.; Kwon, I.; Chae, B. An antimicrobial peptide-A3: Effects on growth performance, nutrient retention, intestinal and faecal microflora and intestinal morphology of broilers. Br. Poult. Sci. 2013, 54, 738–746. [Google Scholar]
- Gyan, W.R.; Yang, Q.; Tan, B.; Jan, S.S.; Jiang, L.; Chi, S.; Dong, X.; Liu, H.; Shuang, Z. Effects of antimicrobial peptides on growth, feed utilization, serum biochemical indices and disease resistance of juvenile shrimp, Litopenaeus vannamei. Aquac. Res. 2020, 51, 1222–1231. [Google Scholar]
- Xiao, B.; Fu, Q.; Niu, S.; Zhu, P.; He, J.; Li, C. Penaeidins restrict white spot syndrome virus infection by antagonizing the envelope proteins to block viral entry. Emerg. Microbes Infect. 2020, 9, 390–412. [Google Scholar]


{kind=link}
| Raw Materials | Dietary Treatments | |||
|---|---|---|---|---|
| Control (g/100 g) | BSFI4.5 (g/100 g) | BSFI7.5 (g/100 g) | BSFI10.5 (g/100 g) | |
| Black Soldier Fly Ingredient a | 0.00 | 4.50 | 7.50 | 10.50 |
| Wheat, flour | 35.25 | 32.74 | 31.74 | 31.59 |
| Soybean meal b | 30.50 | 32.50 | 33.25 | 33.50 |
| Fish meal c | 15.00 | 10.50 | 7.50 | 4.50 |
| Poultry by-product meal d | 7.00 | 7.00 | 7.00 | 7.00 |
| Sepiolite e | 1.50 | 1.49 | 1.21 | 0.60 |
| Fish oil f | 2.84 | 2.90 | 2.84 | 2.96 |
| Squid meal g | 2.50 | 2.50 | 2.65 | 2.65 |
| Soy lecithin h | 1.50 | 1.57 | 1.67 | 1.78 |
| Limestone i | 0.60 | 0.60 | 0.70 | 0.80 |
| Vit min premix j | 0.50 | 0.50 | 0.50 | 0.50 |
| Monocalcium phosphate k | 0.65 | 0.97 | 1.17 | 1.30 |
| Cholesterol, feed grade l | 0.10 | 0.10 | 0.10 | 0.10 |
| Methionine m | 0.05 | 0.12 | 0.16 | 0.21 |
| Gelatine n | 2.00 | 2.00 | 2.00 | 2.00 |
| Astaxanthin premix o | 0.01 | 0.01 | 0.01 | 0.01 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 |
| Parameter | Proximate Analysis | |||
| DM% | 100 | 100 | 100 | 100 |
| Ash% | 7.00 | 7.21 | 7.42 | 7.53 |
| CP% | 40.40 | 40.30 | 40.10 | 39.70 |
| Lipid% | 9.63 | 9.72 | 9.74 | 9.96 |
| Fiber% | 1.82 | 1.86 | 1.87 | 1.88 |
| LOA (18:2n-6)% | 2.03 | 2.08 | 2.15 | 2.23 |
| LNA (18:3n3)% | 0.16 | 0.16 | 0.17 | 0.17 |
| ARA (20:4n-6)% | 0.08 | 0.08 | 0.07 | 0.07 |
| EPA (20:5n-3)% | 0.36 | 0.33 | 0.30 | 0.28 |
| DHA (22:6n-3)% | 0.95 | 0.94 | 0.90 | 0.92 |
| Total n-3% | 1.47 | 1.43 | 1.37 | 1.37 |
| Total n-6% | 2.10 | 2.15 | 2.22 | 2.30 |
| n-3:n-6 | 0.79 | 0.75 | 0.69 | 0.67 |
| Total phospholipid% | 2.56 | 2.51 | 2.51 | 2.51 |
| Cholesterol% | 0.18 | 0.19 | 0.19 | 0.19 |
| Astaxanthin (mg/kg) | 9.03 | 9.00 | 8.99 | 8.98 |
| Arginine% | 2.60 | 2.52 | 2.47 | 2.40 |
| Histidine% | 1.03 | 1.03 | 1.02 | 1.01 |
| Isoleucine% | 1.82 | 1.77 | 1.73 | 1.68 |
| Leucine% | 3.05 | 2.97 | 2.91 | 2.82 |
| Lysine% | 2.70 | 2.57 | 2.48 | 2.36 |
| Methionine% | 0.92 | 0.91 | 0.90 | 0.90 |
| Phenylalanine% | 1.88 | 1.85 | 1.82 | 1.78 |
| Threonine% | 1.63 | 1.59 | 1.56 | 1.52 |
| Tryptophan% | 0.50 | 0.63 | 0.71 | 0.79 |
| Valine% | 2.07 | 2.04 | 2.01 | 1.96 |
| Ca% | 1.49 | 1.42 | 1.42 | 1.40 |
| Available P% | 1.11 | 1.12 | 1.12 | 1.11 |
| Dietary Treatment | Final Body Weight (G) | Weight Gain (G) | Feed Conversion Ratio | Specific Growth Rate (%/Day) | Survival (%) |
|---|---|---|---|---|---|
| Control | 0.52 ± 0.05 | 0.42 ± 0.05 a | 1.70 ± 0.17 a | 6.01 ± 0.33 a | 78.7 ± 14.6 |
| BSFI4.5 | 0.66 ± 0.03 | 0.57 ± 0.03 b | 1.42 ± 0.07 a,b | 6.83 ± 0.14 b | 80.6 ± 2.3 |
| BSFI7.5 | 0.71 ± 0.01 | 0.61 ± 0.01 b,c | 1.31 ± 0.10 b | 7.08 ± 0.22 b,c | 80 ± 8.1 |
| BSFI10.5 | 0.80 ± 0.03 | 0.70 ± 0.03 c | 1.23 ± 0.12 b | 7.53 ± 0.18 c | 84.5 ± 4.3 |
| p value | - | 0.0006 | 0.0067 | 0.0003 | 0.8646 |
| Osmotic Stress | AHPND Challenge | WSSV Challenge | |
|---|---|---|---|
| Dietary Treatment | Final Survival (%) | Final Survival (%) | Final Survival (%) |
| Mock | 0.0 ± 0.0 | 0 ± 0 | 0 ± 0 |
| Control | 33.3 ± 5.8 | 40.0 ± 20.0 | 23.3 ± 23.1 |
| BSFI4.5 | 20.0 ± 10.0 | 31.1 ± 1.9 | 56.7 ± 15.3 |
| BSFI7.5 | 26.7 ± 15.3 | 26.7 ± 15.3 | 43.3 ± 15.3 |
| BSFI10.5 | 16.7 ± 20.8 | 53.3 ± 15.3 | 46.7 ± 11.5 |
| p value | 0.1523 | 0.1458 | 0.0662 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Richardson, A.; Dantas-Lima, J.; Lefranc, M.; Walraven, M. Effect of a Black Soldier Fly Ingredient on the Growth Performance and Disease Resistance of Juvenile Pacific White Shrimp (Litopenaeus vannamei). Animals 2021, 11, 1450. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051450
Richardson A, Dantas-Lima J, Lefranc M, Walraven M. Effect of a Black Soldier Fly Ingredient on the Growth Performance and Disease Resistance of Juvenile Pacific White Shrimp (Litopenaeus vannamei). Animals. 2021; 11(5):1450. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051450
Chicago/Turabian StyleRichardson, Andrew, João Dantas-Lima, Maxime Lefranc, and Maye Walraven. 2021. "Effect of a Black Soldier Fly Ingredient on the Growth Performance and Disease Resistance of Juvenile Pacific White Shrimp (Litopenaeus vannamei)" Animals 11, no. 5: 1450. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051450