Characterization of Alternative Splicing (AS) Events during Chicken (Gallus gallus) Male Germ-Line Stem Cell Differentiation with Single-Cell RNA-seq

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
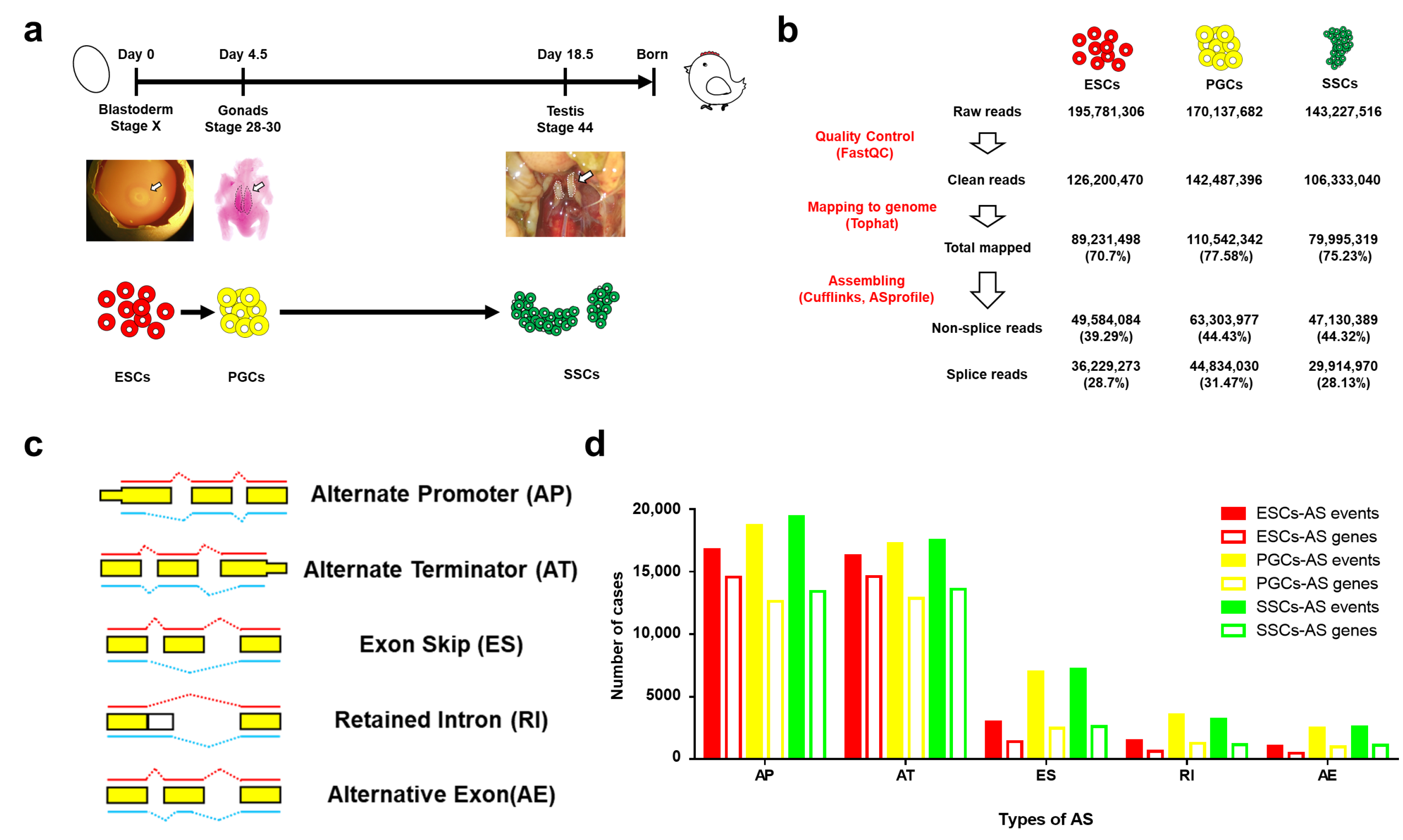
2.2. Cells and RNA-seq
2.3. RNA-Seq Reads Mapping
2.4. Identification, Classification, and Quantification of AS Events
2.5. Intersections and Interaction Network Analysis of the AS Event Gene
2.6. GO Enrichment and KEGG Pathway Analysis of AS Events Gene
2.7. RNA Preparation and RT-qPCR
3. Results
3.1. The AS Events during Male Chicken Germ Cell Differentiation
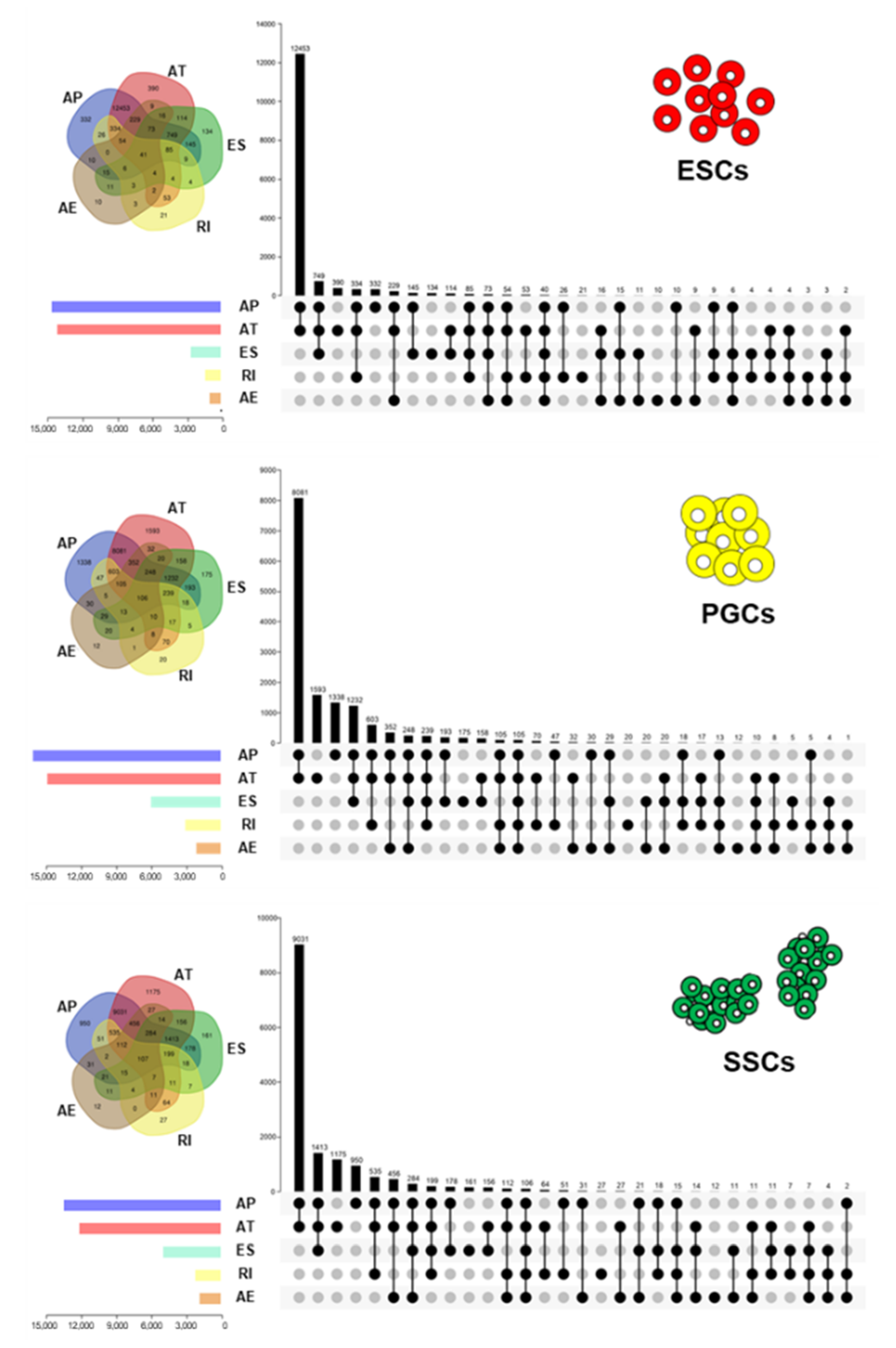
3.2. AS Event Distribution Analysis during Male Chicken Germ Cell Differentiation
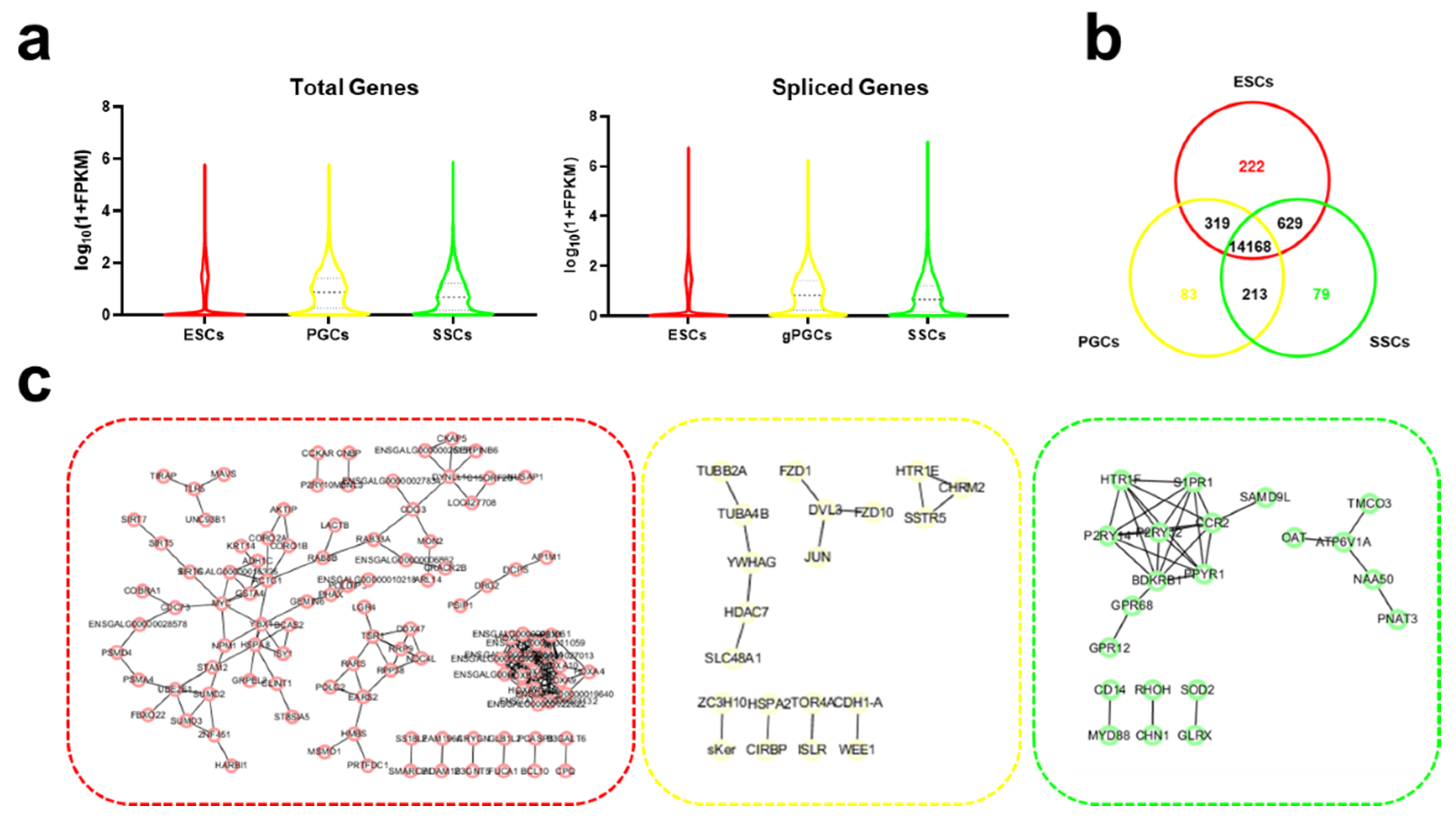
3.3. A Cell-Type-Specific Spliced Gene in ESCs, GPGCs, and SSCs
3.4. Function Enrichment Results of Spliced Genes during Male Germ Cell Differentiation
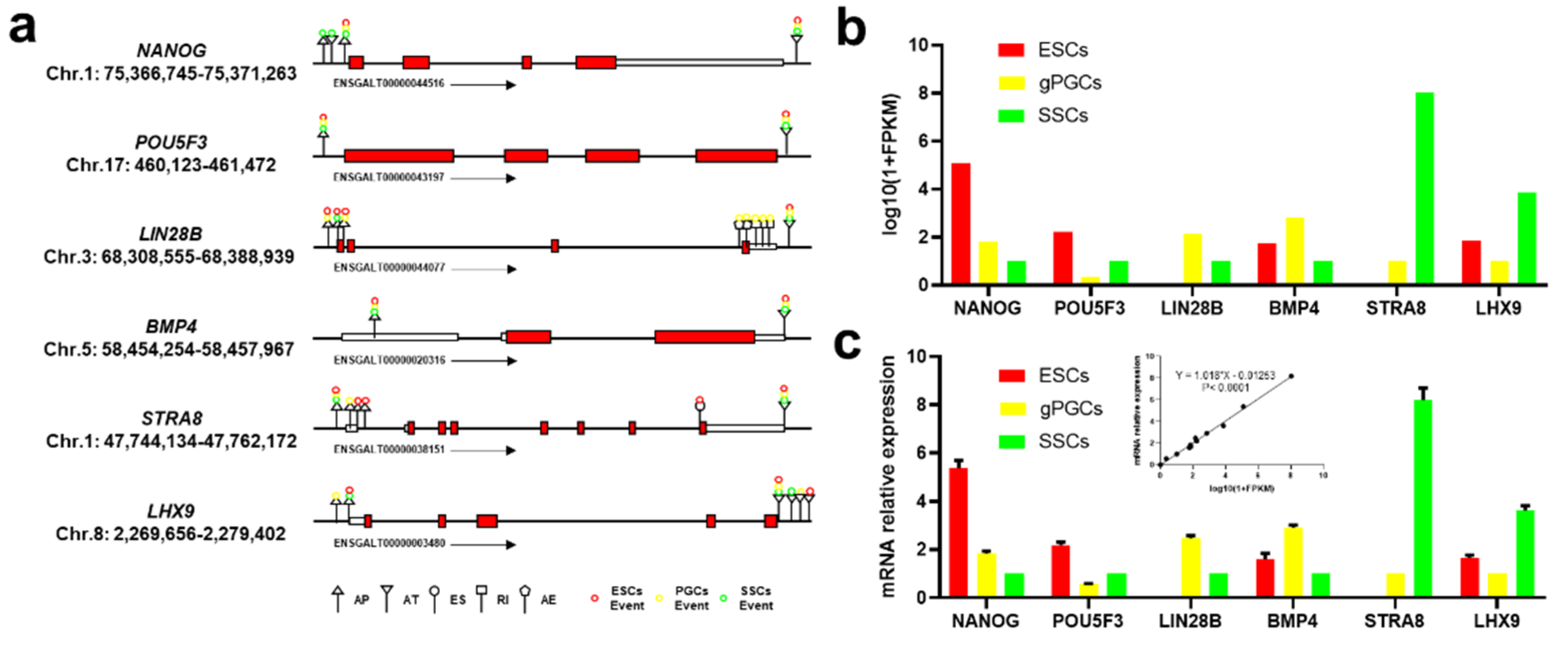
3.5. Validation of AS Events and Crucial Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AS | Alternative Splicing |
| ESCs | Embryonic Stem Cells |
| gPGCs | Primordial Germ Cells |
| SSCs | Spermatogonia Stem Cells |
| AP | Alternative Promoter |
| AT | Alternative Terminator |
| ES | Exon Skipping |
| IR | Intron Retention |
| AE | Alternative Exon |
| FPKM | Fragments Per Kilobase Million |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
References
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nature 2010, 463, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Rio, D.C. Mechanisms and regulation of alternative Pre-mRNA splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Manley, J.L. Mechanisms of alternative splicing regulation: Insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 2009, 10, 741–754. [Google Scholar] [CrossRef]
- Ramani, A.K.; Calarco, J.A.; Pan, Q. Genome-wide analysis of alternative splicing in caenorhabditis elegans. Genome Res. 2011, 21, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Chacko, E.; Ranganathan, S. Genome-wide analysis of alternative splicing in cow: Implications in bovine as a model for human diseases. BMC Genom. 2009, 10, S11. [Google Scholar] [CrossRef] [Green Version]
- Keren, H.; Lev-Maor, G.; Ast, G. Alternative splicing and evolution: Diversification, exon definition and function. Nat. Rev. Genet. 2010, 11, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Yabas, M.; Elliott, H.; Hoyne, G.F. The role of alternative splicing in the control of immune homeostasis and cellular differentiation. Int. J. Mol. Sci. 2016, 17, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zhao, W.; Olson, S.D.; Prabhakara, K.S.; Zhou, X. Alternative splicing links histone modifications to stem cell fate decision. Genome Biol. 2018, 19, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Ramanouskaya, T.V.; Grinev, V.V. The determinants of alternative RNA splicing in human cells. Mol. Genet. Genom. 2017, 292, 1175–1195. [Google Scholar] [CrossRef]
- Fiszbein, A.; Kornblihtt, A.R. Alternative splicing switches: Important players in cell differentiation. Bioessays 2017, 39. [Google Scholar] [CrossRef]
- Spiller, C.M.; Burnet, G.; Bowles, J. Mouse fetal germ cell isolation and culture techniques. Methods Mol. Biol. 2017, 1463, 173–183. [Google Scholar] [CrossRef]
- Smallwood, S.A.; Kelsey, G. De novo DNA methylation: A germ cell perspective. Trends Genet. 2012, 28, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Yamaji, M. Primordial germ cells in mice. Cold Spring Harb. Perspect. Biol. 2012, 4, a008375. [Google Scholar] [CrossRef] [Green Version]
- Yoshizaki, G.; Lee, S. Production of live fish derived from frozen germ cells via germ cell transplantation. Stem Cell Res. 2018, 29, 103–110. [Google Scholar] [CrossRef]
- Tagami, T.; Miyahara, D.; Nakamura, Y. Avian primordial germ cells. Adv. Exp. Med. Biol. 2017, 1001, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.; Souza, G.; Soares, M.; Ribeiro, R.; van den Hurk, R.; Silva, J.V. In vitro differentiation of primordial germ cells and oocyte-like cells from stem cells. Histol. Histopathol. 2018, 33, 121–132. [Google Scholar] [CrossRef]
- Huch, M.; Koo, B.-K. Modeling mouse and human development using organoid cultures. Development 2015, 142, 3113–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuti, N.; Corallo, C.; Chan, B.M.F.; Ferrari, M.; Gerami-Naini, B. Multipotent differentiation of human dental pulp stem cells: A literature Review. Stem Cell Rev. Rep. 2016, 12, 511–523. [Google Scholar] [CrossRef]
- Farzaneh, M.; Attari, F.; Mozdziak, P.E.; Khoshnam, S.E. The evolution of chicken stem cell culture methods. Br. Poult. Sci. 2017, 58, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Han, J.Y. The early development of germ cells in chicken. Int. J. Dev. Biol. 2018, 62, 145–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.G.; Tickle, C. The chicken as a model for embryonic development. Cytogenet. Genome Res. 2007, 117, 231–239. [Google Scholar] [CrossRef]
- Johnsson, M.; Henriksen, R.; Höglund, A.; Fogelholm, J.; Jensen, P.; Wright, D. Genetical genomics of growth in a chicken model. BMC Genom. 2018, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Sliwinska, P.; Segura, T.; Iruela-Arispe, M.L. The chicken chorioallantoic membrane model in biology, medicine and bioengineering. Angiogenesis 2014, 17, 779–804. [Google Scholar] [CrossRef] [Green Version]
- Hannigan, M.M.; Zagore, L.L.; Licatalosi, D.D. Ptbp2 controls an alternative splicing network required for cell communication during spermatogenesis. Cell Rep. 2017, 19, 2598–2612. [Google Scholar] [CrossRef] [Green Version]
- Zagore, L.L.; Grabinski, S.E.; Sweet, T.J. RNA binding protein Ptbp2 is essential for male germ cell development. Mol. Cell Biol. 2015, 35, 4030–4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zheng, Y.; Gao, Y. Single-Cell RNA-Seq uncovers dynamic processes and critical regulators in mouse spermatogenesis. Cell Res. 2018, 28, 879–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinov, G.K.; Williams, B.A.; McCue, K. From single-cell to cell-pool transcriptomes: Stochasticity in gene expression and RNA splicing. Genome Res. 2014, 24, 496–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Ning, B.; Shi, T. Single-cell RNA-Seq technologies and related computational data analysis. Front. Genet. 2019, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Elsayed, A.K.; Shi, Q. Crucial genes and pathways in chicken germ stem cell differentiation. J. Biol. Chem. 2015, 290, 13605–13621. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Zhang, C.; Jin, K.; Zhang, Y.; Zuo, Q.; Li, B. Analysis of LncRNA expression profile during the formation of male germ cells in chickens. Animals 2020, 10, 1850. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L. Differential gene and transcript expression analysis of RNA-Seq experiments with TopHat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000Research 2013, 2, 188. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef]
- Onichtchouk, D. Evolution and functions of Oct4 homologs in non-mammalian vertebrates. Biochim. Biophys. Acta 2016, 1859, 770–779. [Google Scholar] [CrossRef]
- Blinka, S.; Rao, S. Nanog expression in embryonic stem cells—An ideal model system to dissect enhancer function. Bioessays 2017, 39, 1700086. [Google Scholar] [CrossRef] [Green Version]
- Lawson, K.A.; Dunn, N.R.; Roelen, B.A. Bmp4 is required for the generation of primordial germ cells in the mouse embryo. Genes Dev. 1999, 13, 424–436. [Google Scholar] [CrossRef]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.-H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A role for Lin28 in primordial germ-cell development and germ-cell malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, F.; Møller, M.; Fu, C. The Lhx9 homeobox gene controls pineal gland development and prevents postnatal hydrocephalus. Brain Struct. Funct. 2015, 220, 1497–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, Y.; Zuo, Q. CRISPR/Cas9 mediated chicken Stra8 gene knockout and inhibition of male germ cell differentiation. PLoS ONE 2017, 12, e0172207. [Google Scholar] [CrossRef] [PubMed]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef]
- Zhao, S. Alternative splicing, RNA-Seq and drug discovery. Drug Discov. Today 2019, 24, 1258–1267. [Google Scholar] [CrossRef] [PubMed]
- Dvinge, H. Regulation of alternative MRNA splicing: Old players and new perspectives. FEBS Lett. 2018, 592, 2987–3006. [Google Scholar] [CrossRef]
- Song, H.; Wang, L.; Chen, D.; Li, F. The function of Pre-MRNA alternative splicing in mammal spermatogenesis. Int. J. Biol. Sci. 2020, 16, 38–48. [Google Scholar] [CrossRef]
- Guan, Y.; Liang, G.; Martin, G.B.; Guan, L.L. Functional changes in MRNA expression and alternative Pre-MRNA splicing associated with the effects of nutrition on apoptosis and spermatogenesis in the adult testis. BMC Genom. 2017, 18, 64. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Qi, H. Dynamic expression of combinatorial replication-dependent histone variant genes during mouse spermatogenesis. Gene Expr. Patterns 2014, 14, 30–41. [Google Scholar] [CrossRef]
- Lord, T.; Nixon, B. Metabolic changes accompanying spermatogonial stem cell differentiation. Dev. Cell 2020, 52, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, L.F. Calcium explosions as triggers of development. Ann. N. Y. Acad. Sci. 1980, 339, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Brodskiy, P.A.; Zartman, J.J. Calcium as a signal integrator in developing epithelial tissues. Phys. Biol. 2018, 15, 051001. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Lim, S.; Han, J.Y. Wnt/β-catenin signaling pathway activation is required for proliferation of chicken primordial germ cells in vitro. Sci. Rep. 2016, 6, 34510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Du, X.; Zhao, L. SIRT1 facilitates primordial follicle recruitment independent of deacetylase activity through directly modulating Akt1 and MTOR transcription. FASEB J. 2019, 33, 14703–14716. [Google Scholar] [CrossRef] [Green Version]
- Jouni, F.J.; Abdolmaleki, P.; Behmanesh, M.; Movahedin, M. An in vitro study of the impact of 4mT static magnetic field to modify the differentiation rate of rat bone marrow stem cells into primordial germ cells. Differentiation 2014, 87, 230–237. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Transcript ID | Primer (5′-3′) | Title 3 |
|---|---|---|---|
| NANOG | ENSGALT00000044516 | F: TACCCCAGACTCTGCCACTA | 163 bp |
| R: CTTCTGGCTCTGAAACCGC | |||
| POU5F3 | ENSGALT00000043197 | F: GATGCGCCGACCTCAGAG | 112 bp |
| R: CATAGAGCGTGCCCAGAG | |||
| LIN28B | ENSGALT00000044077 | F: CAATGTGAGGATGGGCTTCG | 220 bp |
| R: ACTTCCTAAACAGGGGCTCC | |||
| BMP4 | ENSGALT00000020316 | F: GATCTCTACCGGCTCCAGTC | 174 bp |
| R: GTTGAAGACGAAGCGGATCC | |||
| STRA8 | ENSGALT00000038151 | F: TCCACGGCTATTTCACACCT | 242 bp |
| R: TCAAGGAAACCAGCAGCAAC | |||
| LHX9 | ENSGALT00000003480 | F: GAACTCACCTGCTTTGCCAA | 181 bp |
| R: GAGTCTTGTTGCAGGTGGTG | |||
| GAPDH | ENSGALT00000023323 | F: TGGGAAGCTGTGGAGAGATG | 166 bp |
| R: GCAGGTCAGGTCAACAACAG |
| AS Classification | ESCs | gPGCs | SSCs | |||
|---|---|---|---|---|---|---|
| AS Events | Gene | AS Events | Gene | AS Events | Gene | |
| Alternative 5′ Splice Site (A5′) | 16,741 | 14,560 | 18,697 | 12,639 | 19,391 | 13,402 |
| Alternative 3′ Splice Site (A3′) | 16,273 | 14,609 | 17,217 | 12,874 | 17,515 | 13,601 |
| Exon Skip (ES) | 2964 | 1412 | 7002 | 2487 | 7212 | 2605 |
| Retained Intron (RI) | 1476 | 649 | 3564 | 1271 | 3186 | 1169 |
| Alternative Exon (AE) | 1040 | 486 | 2475 | 995 | 2596 | 1113 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Jin, K.; Zuo, Q.; Sun, H.; Song, J.; Zhang, Y.; Chen, G.; Li, B. Characterization of Alternative Splicing (AS) Events during Chicken (Gallus gallus) Male Germ-Line Stem Cell Differentiation with Single-Cell RNA-seq. Animals 2021, 11, 1469. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051469
Sun C, Jin K, Zuo Q, Sun H, Song J, Zhang Y, Chen G, Li B. Characterization of Alternative Splicing (AS) Events during Chicken (Gallus gallus) Male Germ-Line Stem Cell Differentiation with Single-Cell RNA-seq. Animals. 2021; 11(5):1469. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051469
Chicago/Turabian StyleSun, Changhua, Kai Jin, Qisheng Zuo, Hongyan Sun, Jiuzhou Song, Yani Zhang, Guohong Chen, and Bichun Li. 2021. "Characterization of Alternative Splicing (AS) Events during Chicken (Gallus gallus) Male Germ-Line Stem Cell Differentiation with Single-Cell RNA-seq" Animals 11, no. 5: 1469. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11051469