Effect of Heat Stress on Egg Production, Steroid Hormone Synthesis, and Related Gene Expression in Chicken Preovulatory Follicular Granulosa Cells

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laying Hens, Housing, Heat Stress, and Data Collection
2.2. Isolation, Culture, and Treatment of Granulosa Cells
2.3. Flow Cytometric Analysis of Apoptotic Cells
2.4. Measurement of Cell Viability
2.5. Ultrastructure Observation
2.6. Hormone Secretion Measurements
2.7. RNA Isolation and Quantitative Polymerase Chain Reaction (q-PCR)
2.8. Statistical Analysis
3. Results
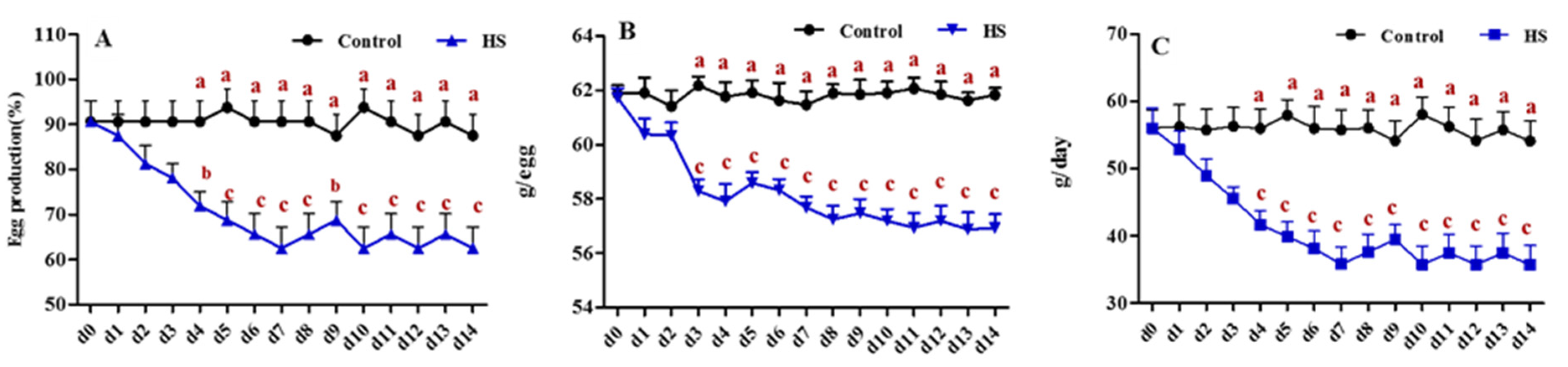
3.1. Effect of Heat Exposure on the Reproductive Activities of Laying Hens
3.2. Effects of Heat Treatment on Cell Viability and Apoptosis of GCs
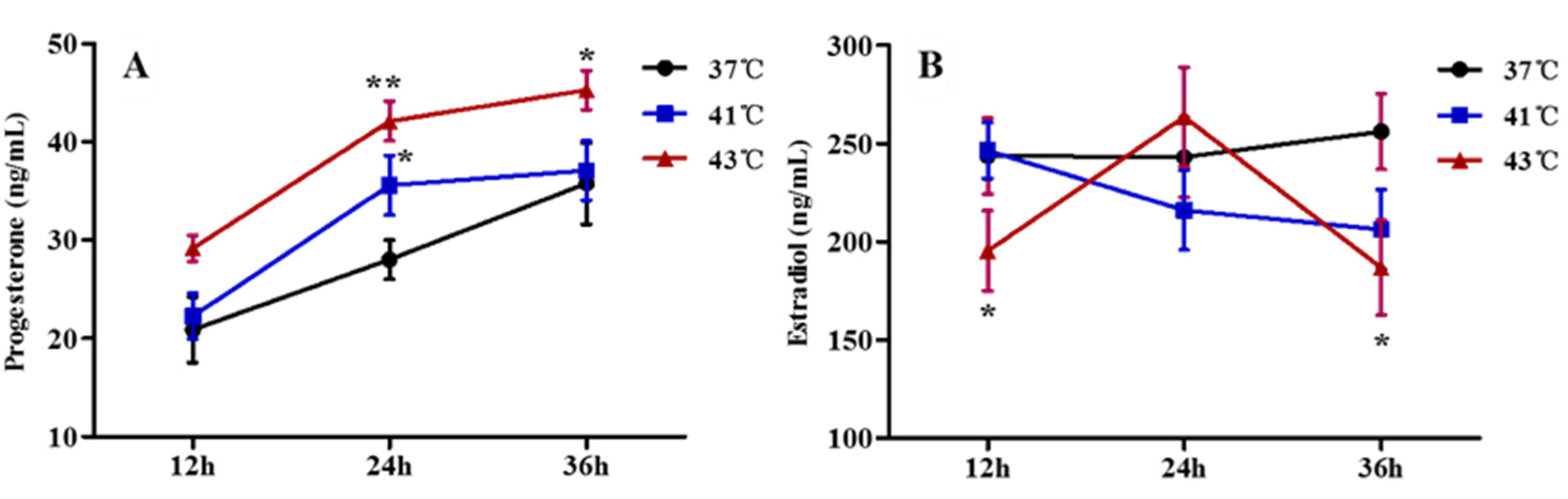
3.3. Effects of Heat Treatment on Progesterone and Estradiol Synthesis in GCs
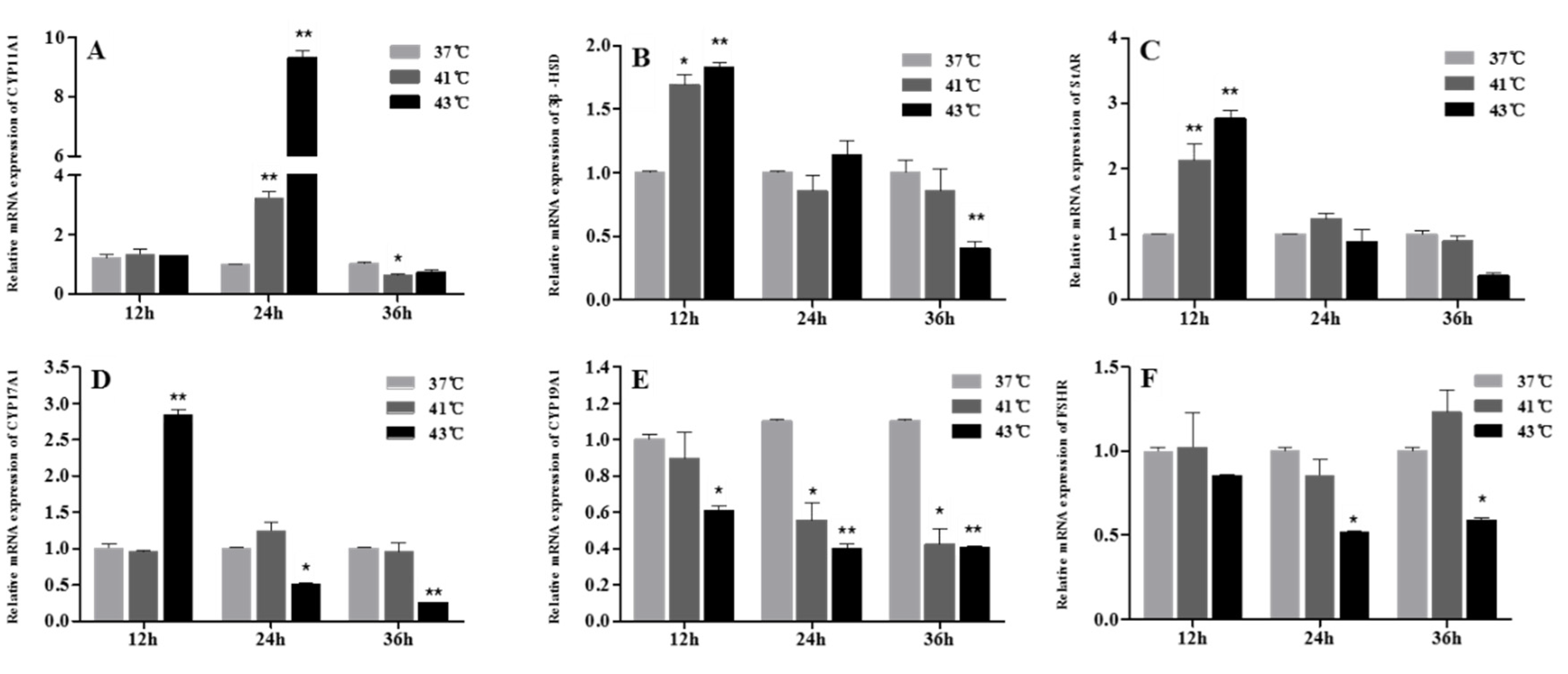
3.4. Effect of Heat Treatment on the Expression of Progesterone- and Estradiol-Synthesizing Enzymes in GCs
3.5. Effect of Heat Treatment on the Expression Levels of HSP70 and HSP90 in GCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mignon-Grasteau, S.; Moreri, U.; Narcy, A.; Rousseau, X.; Rodenburg, T.B.; Tixier-Boichard, M.; Zerjal, T. Robustness to chronic heat stress in laying hens: A meta-analysis. Poult. Sci. 2015, 94, 586–600. [Google Scholar] [CrossRef] [PubMed]
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Hu, J.; Xiong, Y.; Gates, R.S.; Cheng, H.W. Perches as Cooling Devices for Reducing Heat Stress in Caged Laying Hens: A Review. Animals 2021, 11, 3026. [Google Scholar] [CrossRef] [PubMed]
- He, S.P.; Arowolo, M.A.; Medrano, R.F.; Li, S.; Yu, Q.F.; Chen, J.Y. Impact of Heat Stress on Poultry Production. World. Poult. Sci. J. 2018, 74, 647–664. [Google Scholar] [CrossRef]
- Rozenboim, I.; Tako, E.; Gal-Garber, O.; Proudman, J.A.; Uni, Z. The effect of heat stress on ovarian function of laying hens. Poult. Sci. 2007, 86, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Ebeid, T.A.; Suzuki, T.; Sugiyama, T. High temperature influences eggshell quality and calbindin-D28k localization of eggshell gland and all intestinal segments of laying hens. Poult. Sci. 2012, 91, 2282–2287. [Google Scholar] [CrossRef]
- Emery, D.A.; Vohra, P.; Ernst, R.A.; Morrison, S.R. The effect of cyclic and constant ambient temperatures on feed consumption, egg production, egg weight, and shell thickness of hens. Poult. Sci. 1984, 63, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Mashaly, M.M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef]
- Barrett, N.W.; Rowland, K.; Schmidt, C.J.; Lamont, S.J.; Rothschild, M.F.; Ashwell, C.M.; Persia, M.E. Effects of acute and chronic heat stress on the performance, egg quality, body temperature, and blood gas parameters of laying hens. Poult. Sci. 2019, 98, 6684–6692. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, Y.K.; Lee, S.D.; Kim, S.H.; Lee, S.R.; Lee, H.G.; Lee, K.W. Changes in production parameters, egg qualities, fecal volatile fatty acids, nutrient digestibility, and plasma parameters in laying hens exposed to ambient temperature. Front. Vet. Sci. 2020, 7, 412. [Google Scholar] [CrossRef]
- Kim, D.H.; Lee, Y.K.; Kim, S.H.; Lee, K.W. The Impact of Temperature and Humidity on the Performance and Physiology of Laying Hens. Animals 2021, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.Q.; Zadworny, D. Effects of nonglycosylated and glycosylated prolactin on basal and gonadotropin-stimulated steroidogenesis in chicken ovarian follicles. Domest. Anim. Endocrinol. 2017, 61, 27–38. [Google Scholar] [CrossRef]
- Li, G.M.; Liu, L.P.; Yin, B.; Liu, Y.Y.; Dong, W.W.; Gong, S.; Zhang, J.; Tan, J.H. Heat stress decreases egg production of laying hens by inducing apoptosis of follicular cells via activating the FasL/Fas and TNF-α systems. Poult. Sci. 2020, 99, 6084–6093. [Google Scholar] [CrossRef]
- Johnson, A.L.; Solovieva, E.V.; Bridgham, J.T. Relationship between steroidogenic acute regulatory protein expression and progesterone production in hen granulosa cells during follicle development. Biol. Reprod. 2002, 67, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Johnson, A.L. Regulation of P450 cholesterol side-chain cleavage messenger ribonucleic acid expression and progesterone production in hen granulosa cells. Biol. Reprod. 1993, 49, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; McNeilly, A.S. Theca: The forgotten cell of the ovarian follicle. Reproduction 2010, 140, 489–504. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Mohammed, A.A.; Jacobs, J.A.; Cramer, T.A.; Cheng, H.W. Effect of synbiotics on thyroid hormones, intestinal histomorphology, and heat shock protein 70 expression in broiler chickens reared under cyclic heat stress. Poult. Sci. 2020, 99, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xue, W.; Zhang, S.; Guo, Y.; Li, Y.; Wu, X.; Huo, S.; Li, Y.; Li, C. The functions of Patchouli and Elsholtzia in the repair of hen follicular granular cells after heat stress. Poult. Sci. 2022, 101, 101306. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, Y.; Hou, Y.; Yi, Y.; Guo, J.; Zheng, X.; Sun, P.; Sun, Y.; Khan, A.; Li, H. Effects of Osthole on Progesterone Secretion in Chicken Preovulatory Follicles Granulosa Cells. Animals 2020, 10, 2027. [Google Scholar] [CrossRef]
- Sun, L.; Lamont, S.J.; Cooksey, A.M.; McCarthy, F.; Tudor, C.O.; Vijay-Shanker, K.; DeRita, R.M.; Rothschild, M.; Ashwell, C.; Persia, M.E.; et al. Transcriptome Response to Heat Stress in a Chicken Hepatocellular Carcinoma Cell Line. Cell Stress Chaperon. 2015, 20, 939–950. [Google Scholar] [CrossRef] [Green Version]
- Mackei, M.; Molnár, A.; Nagy, S.; Pál, L.; Kővágó, C.; Gálfi, P.; Dublecz, K.; Husvéth, F.; Neogrády, Z.; Mátis, G. Effects of Acute Heat Stress on a Newly Established Chicken Hepatocyte—Nonparenchymal Cell Co-Culture Model. Animals 2020, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Huang, X.B.; Chen, S.J.; Li, X.J.; Fu, X.L.; Xu, D.N.; Tian, Y.B.; Liu, W.J.; Huang, Y.M. The effect of heat stress on proliferation, synthesis of steroids, and gene expression of duck granulosa cells. Anim. Sci. J. 2021, 92, e13617. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Guo, S.; Yan, L.; Zhu, H.; Li, H.; Shi, Z. TNFα-Erk1/2 signaling pathway-regulated SerpinE1 and SerpinB2 are involved in lipopolysaccharide-induced porcine granulosa cell proliferation. Cell. Signal. 2020, 73, 109702. [Google Scholar] [CrossRef]
- Zhu, H.; Shao, X.; Chen, Z.; Wei, C.; Lei, M.; Ying, S.; Yu, J.; Shi, Z. Induction of out-of-season egg laying by artificial photoperiod in Yangzhou geese and the associated endocrine and molecular regulation mechanisms. Anim. Reprod. Sci. 2017, 180, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Chen, Z.; Shao, X.; Yu, J.; Wei, C.; Dai, Z.; Shi, Z. Reproductive axis gene regulation during photostimulation and photorefractoriness in Yangzhou goose ganders. Front. Zool. 2017, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Qu, X.; Chen, Z.; Yu, J.; Yan, L.; Zhu, H. Transcriptome analysis reveals transforming growth factor-β1 prevents extracellular matrix degradation and cell adhesion during the follicular-luteal transition in cows. J. Reprod. Develop. 2022, 68, 12–20. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Oguntunji, O.M.; Alabi, A.O. Influence of high environmental temperature on egg production and shell quality, a review. World. Poult. Sci. J. 2010, 66, 739–774. [Google Scholar] [CrossRef]
- Rubio, M.; Alves, L.; Viana, G.; Benevides, V.P.; Junior, A.B. Heat stress impairs egg production in commercial laying hens infected by fowl typhoid. Avian Pathol. 2020, 50, 1–18. [Google Scholar] [CrossRef]
- Vandana, G.D.; Sejian, V.; Lees, A.M.; Pragna, P.; Maloney, S.K. Heat stress and poultry production: Impact and amelioration. Int. J. Biometeorol. 2021, 65, 163–179. [Google Scholar] [CrossRef]
- Li, L.; Wu, J.; Luo, M.; Sun, Y.; Wang, G. The effect of heat stress on gene expression, synthesis of steroids, and apoptosis in bovine granulosa cells. Cell Stress Chaperon. 2016, 21, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Stocco, D.M. StAR protein and the regulation of steroid hormone biosynthesis. Annu. Rev. Physiol. 2001, 63, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Burris-Hiday, S.D.; Scott, E.E. Steroidogenic cytochrome P450 17A1 structure and function. Mol. Cell. Endocrinol. 2021, 528, 111261. [Google Scholar] [CrossRef] [PubMed]
- Nteeba, J.; Sanz-Fernandez, M.V.; Rhoads, R.P.; Baumgard, L.H.; Ross, J.W.; Keating, A. Heat stress alters ovarian insulin mediated phosphatidylinositol-3 kinase and steroidogenic signaling in gilt ovaries. Biol. Reprod. 2015, 92, 148. [Google Scholar] [CrossRef]
- Heidarzadehpilehrood, R.; Pirhoushiaran, M.; Abdollahzadeh, R.; Binti Osman, M.; Sakinah, M.; Nordin, N.; Abdul Hamid, H. A Review on CYP11A1, CYP17A1, and CYP19A1 Polymorphism Studies: Candidate Susceptibility Genes for Polycystic Ovary Syndrome (PCOS) and Infertility. Genes 2022, 13, 302. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Bauer, M. Heat shock proteins in porcine ovary: Synthesis, accumulation and regulation by stress and hormones. Cell Stress Chaperon. 2011, 16, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maak, S.; Melesse, A.; Schmidt, R.; Schneider, F.; Von Lengerken, G. Effect of long-term heat exposure on peripheral concentrations of heat shock protein 70 (Hsp70) and hormones in laying hens with different genotypes. Br. Poult. Sci. 2003, 44, 133–138. [Google Scholar] [CrossRef]
- Li, H.; Guo, S.; Cai, L.; Ma, W.; Shi, Z. Lipopolysaccharide and heat stress impair the estradiol biosynthesis in granulosa cells via increase of HSP70 and inhibition of smad3 phosphorylation and nuclear translocation. Cell. Signal. 2017, 30, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Khanna, A.; Aten, R.F.; Behrman, H.R. Heat shock protein induction blocks hormone-sensitive steroidogenesis in rat luteal cells. Steroids 1994, 59, 4–9. [Google Scholar] [CrossRef]
- Attia, Y.A.; El-Hamid, A.E.; Abedalla, A.A.; Berika, M.A.; Al-Harthi, M.A.; Kucuk, O.; Sahin, K.; Abou-Shehema, B.M. Laying performance, digestibility and plasma hormones in laying hens exposed to chronic heat stress as affected by betaine, vitamin C, and/or vitamin E supplementation. Springerplus 2016, 5, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Accession Number | Primer Sequence (5′–3′) | Fragment Size (bp) |
|---|---|---|---|
| β-actin | NM_205518.2 | F: TGTCCCTGTATGCCTCTGGTR: GGGCACCTGAACCTCTCATT | 355 |
| HSP70 | NM_001006685.1 | F: CGGGCAAGTTTGACCTAAR: TTGGCTCCCACCCTATCTCT | 250 |
| HSP90 | NM_001109785.2 | F: AGTCCCAGTTCATTGGCTACR: TCCAGTCATTGGTGAGGCT | 324 |
| CYP11A1 | NM_001001756.1 | F: ACCGTGACTACCGCAACAAGR: AGGCCTCCCCTGTCTTGA | 54 |
| 3β-HSD | NM_205118.1 | F: GCCAAAGAGGAGCAAACCAGAGR: TCCAGCAGTAAGCGAACGATCC | 104 |
| StAR | NM_204686.3 | F: CGCTGCCATCTCCTACCAACACAGR: GACATCTCCATCTCGCTGAAGG | 197 |
| FSHR | NM_205079.2 | F: ATGTCTCCGGCAAAGCAAGAR: AACGACTTCGTTGCACAAGC | 147 |
| CYP17A1 | NM_001001901.3 | F: CCACTACCCTGAGGTCCAGAR: GTATTCCCCGATGCTGGTGT | 196 |
| CYP19A1 | NM_001001761.4 | F: CTCGGGGCTGTGTAGGAAAGR: TGTCTGTACTCTGCACCGTC | 86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, L.; Hu, M.; Gu, L.; Lei, M.; Chen, Z.; Zhu, H.; Chen, R. Effect of Heat Stress on Egg Production, Steroid Hormone Synthesis, and Related Gene Expression in Chicken Preovulatory Follicular Granulosa Cells. Animals 2022, 12, 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111467
Yan L, Hu M, Gu L, Lei M, Chen Z, Zhu H, Chen R. Effect of Heat Stress on Egg Production, Steroid Hormone Synthesis, and Related Gene Expression in Chicken Preovulatory Follicular Granulosa Cells. Animals. 2022; 12(11):1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111467
Chicago/Turabian StyleYan, Leyan, Mengdie Hu, Lihong Gu, Mingming Lei, Zhe Chen, Huanxi Zhu, and Rong Chen. 2022. "Effect of Heat Stress on Egg Production, Steroid Hormone Synthesis, and Related Gene Expression in Chicken Preovulatory Follicular Granulosa Cells" Animals 12, no. 11: 1467. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12111467