Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress

 , , , ,
, , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms and Maintenance
2.2. Preparation of Postbiotics from L. plantarum Strains
2.3. Animals Housing and Experimental Design
2.4. Sample Collection
2.5. Growth Performance Measurements and Mortality
2.6. Carcass Characteristics
2.7. Intestinal Morphology
2.8. Caecum Microbial Population
2.9. Plasma Immunoglobulin Concentration
2.10. RNA Extraction and RT-PCR of GHR and IGF-1 Genes
2.11. Statistical Analysis
3. Results
3.1. Growth Performance and Mortality
3.2. Carcass Yield
3.3. Intestinal Histomorphology
3.4. Caecum Microbial Population and pH
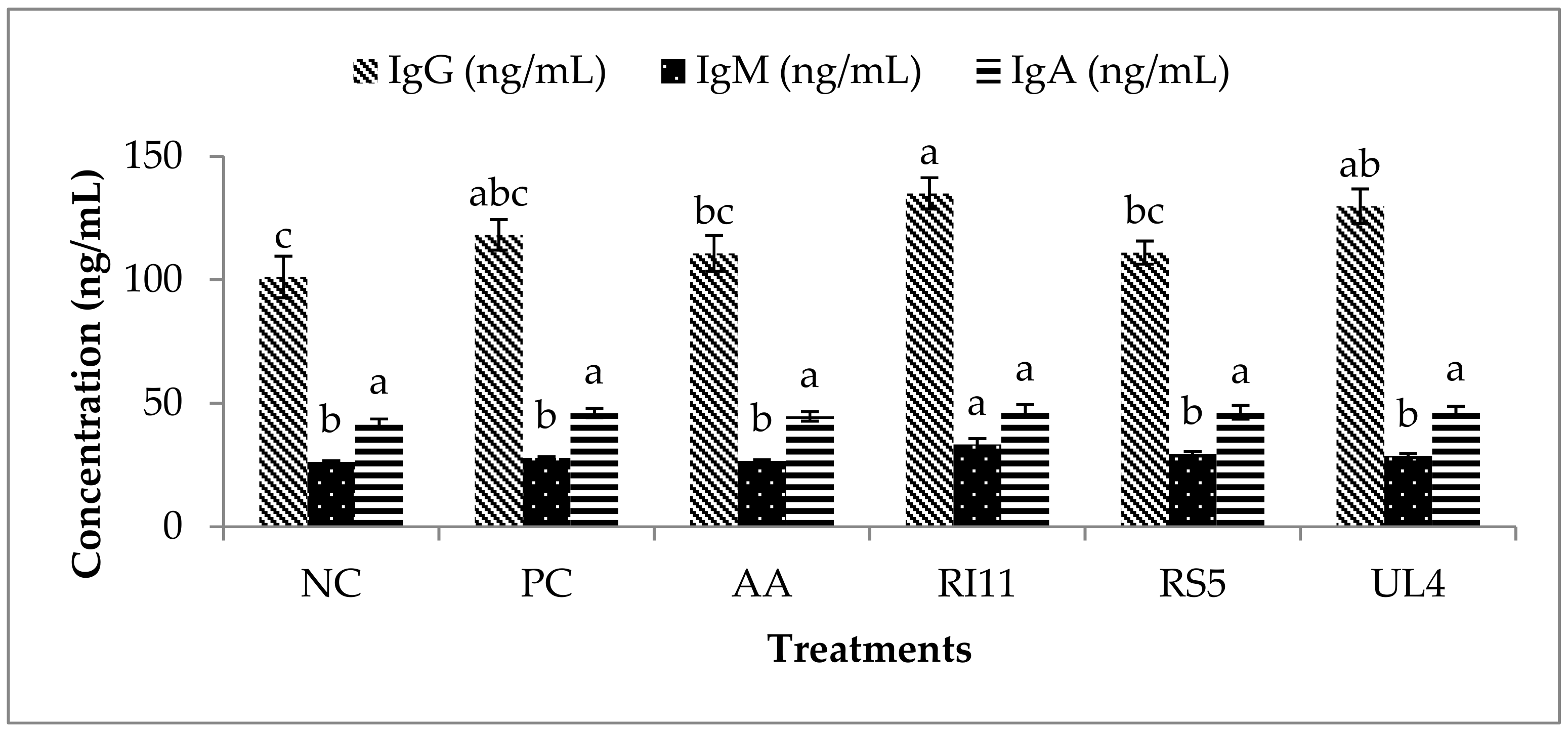
3.5. Plasma IgG, IgM and IgA Concentration
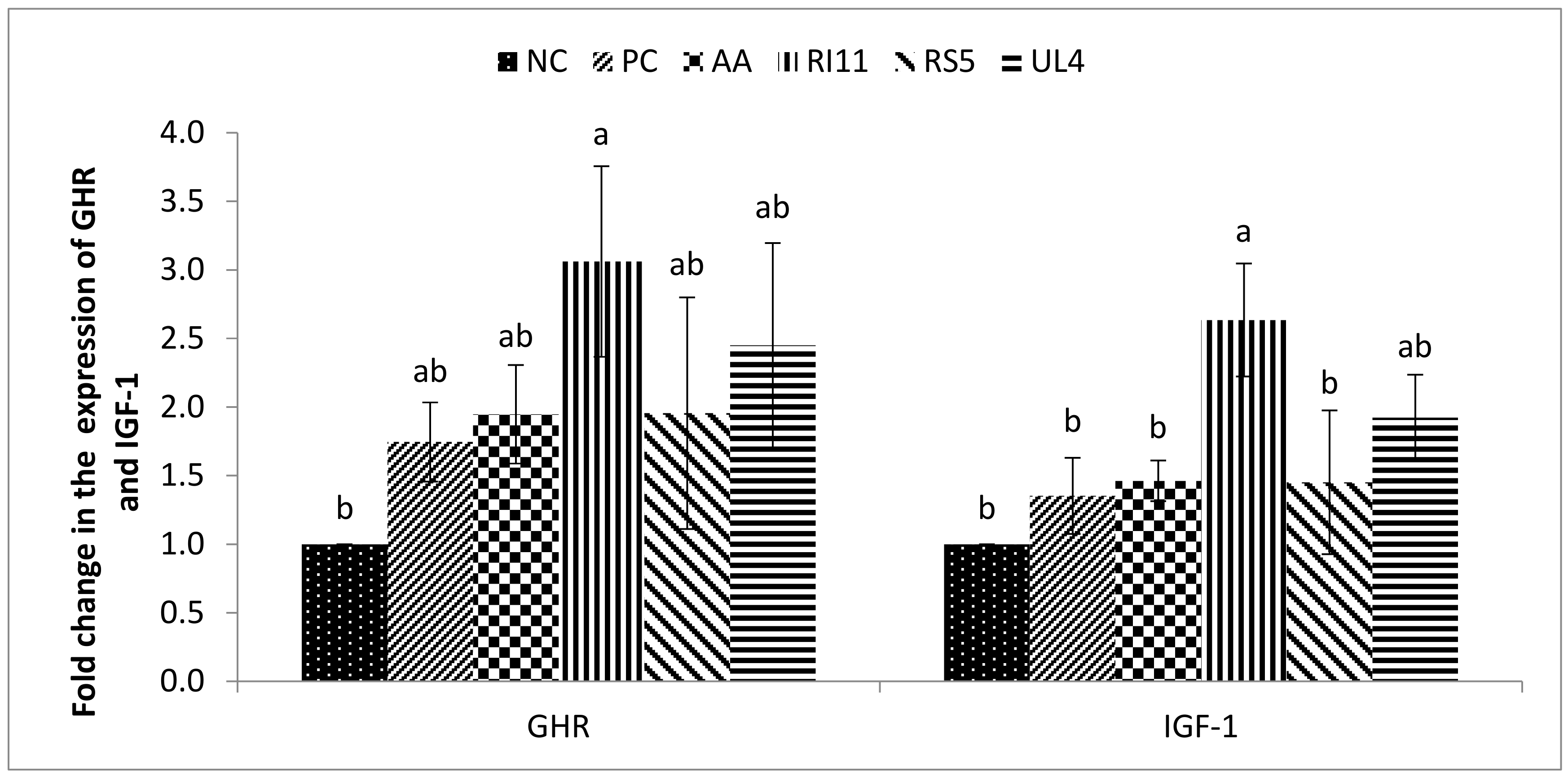
3.6. Hepatic GHR and IGF-1 Gene Expression Levels
4. Discussion
4.1. Growth Performance, Mortality and Carcass Yield
4.2. Intestinal Histomorphology
4.3. Caecum Microbial Population and pH
4.4. Plasma IgG, IgM and IgA Concentrations
4.5. Hepatic IGF-1 and GHR Gene Expression
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Najafi, P.; Zulkifli, I.; Soleimani, A.F.; Kashiani, P. The effect of different degrees of feed restriction on heat shock protein 70, acute phase proteins, and other blood parameters in female broiler breeders. Poult. Sci. 2015, 94, 2322–2329. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lu, Z.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Effects of chronic heat exposure on growth performance, intestinal epithelial histology, appetite–related hormones and genes expression in broilers. J. Sci. Food Agric. 2018, 98, 4471–4478. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, K.Z.; Edens, F.W.; Eisen, E.J.; Havenstein, G.B. Effect of ascorbic acid and acute heat exposure on heat shock protein 70 expression by young white Leghorn chickens. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 136, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, S.; Lasagna, E.; Sarti, F.M.; Marelli, S.P.; Cozzi, M.C.; Bernardini, G.; Terova, G. Expression profile of six stress-related genes and productive performances of fast and slow growing broiler strains reared under heat stress conditions. Meta Gene 2015, 6, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.; Kikusato, M.; Maekawa, T.; Shirakawa, H.; Toyomizu, M. Metabolic characteristics and oxidative damage to skeletal muscle in broiler chickens exposed to chronic heat stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 155, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Al-Aqil, A.; Zulkifli, I.; Sazili, A.; Omar, A.; Rajion, M. The effects of the hot, humid tropical climate and early age feed restriction on stress and fear responses, and performance in broiler chickens. Asian-Australas J. Anim. Sci. 2009, 22, 1581–1586. [Google Scholar] [CrossRef]
- Syafwan, S.; Kwakkel, R.; Verstegen, M. Heat stress and feeding strategies in meat-type chickens. Worlds Poult. Sci. J. 2011, 67, 653–674. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, S.; Geraert, P.; Lessire, M.; Carre, B.; Guillaumin, S. Effect of high ambient temperature on feed digestibility in broilers. Poult. Sci. 1997, 76, 857–863. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Tufarelli, V.; Dario, M.; Laudadio, V. Effect of xylanase supplementation and particle-size on performance of guinea fowl broilers fed wheat-based diets. Int. J. Poult. Sci. 2007, 4, 302–307. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sakai, M.; Sá, L.R.M.d.; Ferreira, A.J.P.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Khaksefidi, A. A comparison between the effects of a probiotic (Bioplus 2B) and an antibiotic (virginiamycin) on the performance of broiler chickens under heat stress condition. Iran. J. Vet. Res. 2006, 7, 23–28. [Google Scholar]
- Zulkifli, I.; Abdullah, N.; Azrin, N.M.; Ho, Y. Growth performance and immune response of two commercial broiler strains fed diets containing Lactobacillus cultures and oxytetracycline under heat stress conditions. Br. Poult. Sci. 2000, 41, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Odore, R.; De Marco, M.; Gasco, L.; Rotolo, L.; Meucci, V.; Palatucci, A.; Rubino, V.; Ruggiero, G.; Canello, S.; Guidetti, G. Cytotoxic effects of oxytetracycline residues in the bones of broiler chickens following therapeutic oral administration of a water formulation. Poult. Sci. 2015, 94, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Shazali, N.; Foo, H.L.; Loh, T.C.; Choe, D.W.; Rahim, R.A. Prevalence of antibiotic resistance in lactic acid bacteria isolated from the faeces of broiler chicken in Malaysia. Gut Pathog. 2014, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Regulation, E. No 1831/2003 of the European Parliament and Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Commun. 2003, 268, 29–43. [Google Scholar]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verghese, R.J.; Ramya, S.; Kanungo, R. In vitro Antibacterial Activity of Vitamin C and in Combination with Ciprofloxacin against Uropathogenic Escherichia coli. J. Clin. Diagn. Res. 2017, 11, 1–5. [Google Scholar] [CrossRef]
- Tajkarimi, M.; Ibrahim, S.A. Antimicrobial activity of ascorbic acid alone or in combination with lactic acid on Escherichia coli O157: H7 in laboratory medium and carrot juice. Food Control 2011, 22, 801–804. [Google Scholar] [CrossRef]
- Njoku, P. Effect of dietary ascorbic acid (vitamin C) supplementation on the performance of broiler chickens in a tropical environment. Anim. Feed Sci. Technol. 1986, 16, 17–24. [Google Scholar] [CrossRef]
- Kadim, I.; Al-Qamshui, B.; Mahgoub, O.; Al-Marzooqi, W.; Johnson, E. Effect of seasonal temperatures and ascorbic acid supplementation on performance of broiler chickens maintained in closed and open-sided houses. Int. J. Poult. Sci. 2008, 7, 655–660. [Google Scholar] [CrossRef]
- Kutlu, H.; Forbes, J. Changes in growth and blood parameters in heat-stressed broiler chicks in response to dietary ascorbic acid. Livest. Prod. Sci. 1993, 36, 335–350. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Kucuk, O. Effects of chromium, and ascorbic acid supplementation on growth, carcass traits, serum metabolites, and antioxidant status of broiler chickens reared at a high ambient temperature (32°C). Nutr. Res. 2003, 23, 225–238. [Google Scholar] [CrossRef]
- Ferreira, I.; Matos Junior, J.; Sgavioli, S.; Vicentini, T.; Morita, V.; Boleli, I. Vitamin C prevents the effects of high rearing temperatures on the quality of broiler thigh meat. Poult. Sci. 2015, 94, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Yudiarti, T.; Isroli, I.; Widiastuti, E.; Kusumanti, E. Dietary supplementation of probiotics in poultry exposed to heat stress—A review. Ann. Anim. Sci. 2017, 17, 591–604. [Google Scholar] [CrossRef]
- Gibson, G.; Saavedra, J.; MacFarlane, S.; MacFarlane, G. Probiotics and intestinal infections. In Probiotics 2; Springer: Berlin/Heidelberg, Germany, 1997; pp. 10–39. [Google Scholar]
- Jahromi, M.F.; Altaher, Y.W.; Shokryazdan, P.; Ebrahimi, R.; Ebrahimi, M.; Idrus, Z.; Tufarelli, V.; Liang, J.B. Dietary supplementation of a mixture of Lactobacillus strains enhances performance of broiler chickens raised under heat stress conditions. Int. J. Biometeorol. 2016, 60, 1099–1110. [Google Scholar] [CrossRef]
- Egervärn, M.; Roos, S.; Lindmark, H. Identification and characterization of antibiotic resistance genes in Lactobacillus reuteri and Lactobacillus plantarum. J. Appl. Microbiol. 2009, 107, 1658–1668. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; de los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Marteau, P.; Shanahan, F. Basic aspects and pharmacology of probiotics: An overview of pharmacokinetics, mechanisms of action and side-effects. Best Pract. Res. Clin. Gastroenterol. 2003, 17, 725–740. [Google Scholar] [CrossRef]
- Thanh, N.T.; Loh, T.C.; Foo, H.L.; Hair-Bejo, M.; Azhar, B.K. Effects of feeding metabolite combinations produced by Lactobacillus plantarum on growth performance, faecal microbial population, small intestine villus height and faecal volatile fatty acids in broilers. Br. Poult. Sci. 2009, 50, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.; Garcia-Varela, R.; Garcia, H.; Mata-Haro, V.; González-Córdova, A.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Ling, F.H.; Chwen, L.T.; Foong, O.M.; Asmara, S.A. Inhibitory activity of postbiotic produced by strains of Lactobacillus plantarum using reconstituted media supplemented with inulin. Gut Pathog. 2014, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Choe, D.W.; Foo, H.L.; Loh, T.C.; Hair-Bejo, M.; Awis, Q.S. Inhibitory property of metabolite combinations produced from Lactobacillus plantarum strains. Pertanika J. Trop. Agric. Sci. 2013, 36, 79–88. [Google Scholar]
- Thanh, N.T.; Chwen, L.T.; Foo, H.L.; Hair-Bejo, M.; Kasim, A.B. Inhibitory activity of metabolites produced by strains of Lactobacillus plantarum isolated from Malaysian fermented food. Int. J. Probiotics Prebiotics 2010, 5, 37. [Google Scholar]
- Van Thu, T.; Foo, H.L.; Loh, T.C.; Bejo, M.H. Inhibitory activity and organic acid concentrations of metabolite combinations produced by various strains of Lactobacillus plantarum. Afr. J. Biotechnol. 2011, 10, 1359–1363. [Google Scholar]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Akit, H.; Samsudin, A.A. Effects of dietary postbiotic and inulin on growth performance, IGF1 and GHR mRNA expression, faecal microbiota and volatile fatty acids in broilers. BMC Vet. Res. 2016, 12, 163. [Google Scholar] [CrossRef]
- Loh, T.C.; Thanh, N.T.; Foo, H.L.; Hair-Bejo, M.; Azhar, B.K. Feeding of different levels of metabolite combinations produced by Lactobacillus plantarum on growth performance, fecal microflora, volatile fatty acids and villi height in broilers. Anim. Sci. J. 2010, 81, 205–214. [Google Scholar] [CrossRef]
- Rosyidah, M.; Loh, T.; Foo, H.; Cheng, X.; Bejo, M. Effect of feeding metabolites and acidifier on growth performance, faecal characteristics and microflora in broiler chickens. J. Anim. Vet. Adv. 2011, 10, 2758–2764. [Google Scholar]
- Loh, T.C.; Choe, D.W.; Foo, H.L.; Sazili, A.Q.; Bejo, M.H. Effects of feeding different postbiotic metabolite combinations produced by Lactobacillus plantarum strains on egg quality and production performance, faecal parameters and plasma cholesterol in laying hens. BMC Vet. Res. 2014, 10, 149. [Google Scholar] [CrossRef]
- Choe, D.W.; Loh, T.C.; Foo, H.L.; Hair-Bejo, M.; Awis, Q.S. Egg production, faecal pH and microbial population, small intestine morphology, and plasma and yolk cholesterol in laying hens given liquid metabolites produced by Lactobacillus plantarum strains. Br. Poult. Sci. 2012, 53, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Loh, T.C.; Thu, T.V.; Foo, H.L.; Bejo, M.H. Effects of different levels of metabolite combination produced by Lactobacillus plantarum on growth performance, diarrhoea, gut environment and digestibility of postweaning piglets. J. Appl. Anim. Res. 2013, 41, 200–207. [Google Scholar] [CrossRef]
- Thu, T.V.; Loh, T.C.; Foo, H.L.; Yaakub, H.; Bejo, M.H. Effects of liquid metabolite combinations produced by Lactobacillus plantarum on growth performance, faeces characteristics, intestinal morphology and diarrhoea incidence in postweaning piglets. Trop. Anim. Health Prod. 2011, 43, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Izuddin, W.I.; Loh, T.C.; Samsudin, A.A.; Foo, H.L. In vitro study of postbiotics from Lactobacillus plantarum RG14 on rumen fermentation and microbial population. Rev. Bras. Zootec. 2018, 47. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Humam, A.M. Postbiotic Lactobacillus plantarum RG14 improves ruminal epithelium growth, immune status and upregulates the intestinal barrier function in post-weaning lambs. Sci. Rep. 2019, 9, 9938. [Google Scholar] [CrossRef] [PubMed]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Asmara, S.A.; Akit, H. Influence of postbiotic RG14 and inulin combination on cecal microbiota, organic acid concentration, and cytokine expression in broiler chickens. Poult. Sci. 2017, 96, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Jang, N.Y.; Kim, Y.T. Isolation of Lactic Acid Bacteria Showing Antioxidative and Probiotic Activities from Kimchi and Infant Feces. J. Microbiol. Biotechnol. 2015, 25, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Wang, X.; Li, G.; Zhao, Y.; Zhang, J.; Niu, C.; Zhang, L.; Zhang, X.; Ying, D.; Li, S. Antioxidant activity of prebiotic ginseng polysaccharides combined with potential probiotic Lactobacillus plantarum C88. Int. J. Food Sci. Technol. 2015, 50, 1673–1682. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef]
- Moghadam, M.S.; Foo, H.L.; Leow, T.C.; Rahim, R.A.; Loh, T.C. Novel Bacteriocinogenic Lactobacillus plantarum Strains and Their Differentiation by Sequence Analysis of 16 S rDNA, 16 S-23 S and 23 S-5 S Intergenic Spacer Regions and Randomly Amplified Polymorphic DNA Analysis. Food Technol. Biotechnol. 2010, 48, 476–483. [Google Scholar]
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J. Anim. Sci. Biotechnol. 2019, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Foo, H.; Loh, T.; Law, F.; Lim, Y.; Kuflin, C.; Rusul, G. Effect of feeding L. plantarum I-UL4 isolated from Malaysian Tempeh on growth performance, fecla flora and lactic acid bacteria and plasma cholesterol concentrations in post weaning rats. J. Food Sci. Biotechnol. 2003, 12, 403–408. [Google Scholar]
- Loh, T.; Chong, S.; Foo, H.; Law, F. Effects on growth performance, faecal microflora and plasma cholesterol after supplementation of spray-dried metabolite to postweaning rats. Czech J. Anim. Sci. 2009, 54, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Cobb-Vantress, S.S. Cobb 500 Broiler Performance and Nutrition Supplement; Siloam Springs: Benton County, Arkansas, USA, 2008; pp. 1–14. Available online: cobb-vantress.com (accessed on 26 July 2019).
- Malaysian Standard, M. Halal Food—Production, Preparation, Handling and Storage—General Guideline; Department of Standards Malaysia: Cyberjaya, Malaysia, 2009; pp. 1–26. Available online: http://www.standardsmalaysia.gov.my (accessed on 26 July 2019).
- Touchette, K.; Carroll, J.; Allee, G.; Matteri, R.; Dyer, C.; Beausang, L.; Zannelli, M. Effect of spray-dried plasma and lipopolysaccharide exposure on weaned pigs: I. Effects on the immune axis of weaned pigs. J. Anim. Sci. 2002, 80, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Navidshad, B.; Liang, J.B.; Jahromi, M.F. Correlation coefficients between different methods of expressing bacterial quantification using real time PCR. Int. J. Mol. Sci. 2012, 13, 2119–2132. [Google Scholar] [CrossRef]
- Jahromi, M.F.; Liang, J.B.; Ebrahimi, R.; Soleimani, A.F.; Rezaeizadeh, A.; Abdullah, N.; Shokryazdan, P. Protective potential of Lactobacillus species in lead toxicity model in broiler chickens. Animal 2017, 11, 755–761. [Google Scholar] [CrossRef]
- Bartosch, S.; Fite, A.; Macfarlane, G.T.; McMurdo, M.E. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl. Environ. Microbiol. 2004, 70, 3575–3581. [Google Scholar] [CrossRef]
- Nam, H.-M.; Srinivasan, V.; Gillespie, B.E.; Murinda, S.E.; Oliver, S.P. Application of SYBR green real-time PCR assay for specific detection of Salmonella spp. in dairy farm environmental samples. Int. J. Food Microbiol. 2005, 102, 161–171. [Google Scholar] [CrossRef]
- Del Vesco, A.; Gasparino, E.; Oliveira Neto, A.; Guimaraes, S.; Voltolini, D. Dietary methionine effects on IGF-I and GHR mRNA expression in broilers. Genet. Mol. Res. 2013, 12, 6414–6423. [Google Scholar] [CrossRef]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Moeini, H.; Ideris, A.; Bejo, M.H.; Alitheen, N.B.M.; Kaiser, P.; Omar, A.R. Alteration in lymphocyte responses, cytokine and chemokine profiles in chickens infected with genotype VII and VIII velogenic Newcastle disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 11–21. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mookiah, S.; Sieo, C.C.; Ramasamy, K.; Abdullah, N.; Ho, Y.W. Effects of dietary prebiotics, probiotic and synbiotics on performance, caecal bacterial populations and caecal fermentation concentrations of broiler chickens. J. Sci. Food Agric. 2014, 94, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Kalavathy, R.; Abdullah, N.; Jalaludin, S.; Ho, Y.W. Effects of Lactobacillus cultures on growth performance, abdominal fat deposition, serum lipids and weight of organs of broiler chickens. Br. Poult. Sci. 2003, 44, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.U.; Hume, M.E.; Byrd, J.A.; Nisbet, D.J.; Ijaz, A.; Sohail, A.; Shabbir, M.Z.; Rehman, H. Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth performance of broilers subjected to chronic heat stress. Poult. Sci. 2012, 91, 2235–2240. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.S.; Ko, Y.H.; Moon, Y.S.; Sohn, S.H. Effects of Vitamin C or E on the Pro-inflammatory Cytokines, Heat Shock Protein 70 and Antioxidant Status in Broiler Chicks under Summer Conditions. Asian-Australas J. Anim. Sci. 2014, 27, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; El, A.E.-H.E.A.; Abedalla, A.A.; Berika, M.A.; Al-Harthi, M.A.; Kucuk, O.; Sahin, K.; Abou-Shehema, B.M. Laying performance, digestibility and plasma hormones in laying hens exposed to chronic heat stress as affected by betaine, vitamin C, and/or vitamin E supplementation. SpringerPlus 2016, 5, 1619. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Asmara, S.A.; Akit, H.; Abdulla, N.R.; Ooi, M.F. Carcass, meat and bone quality of broiler chickens fed with postbiotic and prebiotic combinations. Int. J. Probiotics Prebiotics 2015, 10, 23. [Google Scholar]
- Shimokomakillal, A.O.M. The effects of biotic additives on growth performance and meat qualities in broiler chickens. Int. J. Poult. Sci. 2012, 11, 599–604. [Google Scholar] [CrossRef]
- Abdel-Raheem, S.M.; Abd-Allah, S.M. The effect of single or combined dietary supplementation of mannan oligosacharide and probiotics on performance and slaughter characteristics of broilers. Int. J. Poult. Sci. 2011, 10, 854–862. [Google Scholar]
- Pelicano, E.R.L.; De Souza, P.; De Souza, H.; Oba, A.; Norkus, E.; Kodawara, L.; De Lima, T. Effect of different probiotics on broiler carcass and meat quality. Braz. J. Poult. Sci. 2003, 5, 207–214. [Google Scholar] [CrossRef] [Green Version]
- de Souza, L.F.; Araújo, D.N.; Stefani, L.M.; Giometti, I.C.; Cruz-Polycarpo, V.C.; Polycarpo, G.; Burbarelli, M.F. Probiotics on performance, intestinal morphology and carcass characteristics of broiler chickens raised with lower or higher environmental challenge. Austral. J. Vet. Sci. 2018, 50, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, Ö.; Köksal, B.H.; Tatlı, O.; Sevim, Ö.; Ahsan, U.; Üner, A.G.; Ulutaş, P.A.; Beyaz, D.; Büyükyörük, S.; Yakan, A. Effect of dietary probiotic and high stocking density on the performance, carcass yield, gut microflora, and stress indicators of broilers. Poult. Sci. 2015, 94, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Uni, Z.; Noy, Y.; Sklan, D. Posthatch changes in morphology and function of the small intestines in heavy- and light-strain chicks. Poult. Sci. 1995, 74, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Caspary, W.F. Physiology and pathophysiology of intestinal absorption. Am. J. Clin. Nutr. 1992, 55, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Hu, C.; Xia, M.; Zhan, X.; Wang, M. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult. Sci. 2003, 82, 1030–1036. [Google Scholar] [CrossRef]
- Fan, Y.; Croom, J.; Christensen, V.; Black, B.; Bird, A.; Daniel, L.; McBride, B.; Eisen, E. Jejunal glucose uptake and oxygen consumption in turkey poults selected for rapid growth. Poult. Sci. 1997, 76, 1738–1745. [Google Scholar] [CrossRef]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503. [Google Scholar] [CrossRef]
- Suzuki, K.; Harasawa, R.; Yoshitake, Y.; Mitsuoka, T. Effects of crowding and heat stress on intestinal flora, body weight gain, and feed efficiency of growing rats and chicks. Nihon juigaku zasshi. J. Vet. Med. Sci. 1983, 45, 331. [Google Scholar] [CrossRef]
- Loh, T.; Thanh, N.; Foo, H.; Hair-Bejo, M. Effects of feeding metabolite combinations from lactobacillus plantarum on plasma and breast meat lipids in Broiler Chickens. Rev. Bras. Cienc. Avic. 2013, 15, 307–316. [Google Scholar] [CrossRef]
- Wang, J.; Han, M.; Zhang, G.; Qiao, S.; Li, D.; Ma, X. The signal pathway of antibiotic alternatives on intestinal microbiota and immune function. Curr. Protein Pept. Sci. 2016, 17, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Steer, T.; Carpenter, H.; Tuohy, K.; Gibson, G.R. Perspectives on the role of the human gut microbiota and its modulation by pro-and prebiotics. Nutr. Res. Rev. 2000, 13, 229–254. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, J.; Zhang, S.; Yang, F.; Thacker, P.A.; Zhang, G.; Qiao, S.; Ma, X. Oral administration of Lactobacillus fermentum I5007 favors intestinal development and alters the intestinal microbiota in formula-fed piglets. J. Agric. Food Chem. 2014, 62, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Gil De Los Santos, J.; Storch, O.; Gil-Turnes, C. Bacillus cereus var. toyoii and Saccharomyces boulardii increased feed efficiency in broilers infected with Salmonella enteritidis. Br. Poult. Sci. 2005, 46, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS ONE 2013, 8, e70215. [Google Scholar] [CrossRef]
- Burkholder, K.; Thompson, K.; Einstein, M.; Applegate, T.; Patterson, J. Influence of stressors on normal intestinal microbiota, intestinal morphology, and susceptibility to Salmonella enteritidis colonization in broilers. Poult. Sci. 2008, 87, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Ehrenstein, M.R.; Cook, H.T.; Neuberger, M.S. Deficiency in serum immunoglobulin (Ig)M predisposes to development of IgG autoantibodies. J. Exp. Med. 2000, 191, 1253–1258. [Google Scholar] [CrossRef]
- Bienenstock, J.; Gauldie, J.; Perey, D.Y. Synthesis of IgG, IgA, IgM by chicken tissues: Immunofluorescent and 14C amino acid incorporation studies. J. Immunol. 1973, 111, 1112–1118. [Google Scholar]
- Shamblott, M.J.; Cheng, C.M.; Bolt, D.; Chen, T.T. Appearance of insulin-like growth factor mRNA in the liver and pyloric ceca of a teleost in response to exogenous growth hormone. Proc. Natl. Acad. Sci. USA 1995, 92, 6943–6946. [Google Scholar] [CrossRef]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Natl. Acad. Sci. USA 2016, 113, E7554–E7563. [Google Scholar] [CrossRef]
- Yan, J.; Charles, J.F. Gut Microbiota and IGF-1. Calcif. Tissue Int. 2018, 102, 406–414. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients% | Treatments Diets 1 | |||||
|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |
| Corn | 50 | 50 | 50 | 50 | 50 | 50 |
| Soybean meal 48% | 34.5 | 34.5 | 34.5 | 34.5 | 34.5 | 34.5 |
| Palm oil | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 | 6.3 |
| Wheat pollard | 4 | 3.98 | 3.98 | 3.7 | 3.7 | 3.7 |
| DCP 18% 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Calcium carbonate | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 | 1.8 |
| Choline chloride | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Salt | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 |
| DL-Methionine | 0.32 | 0.32 | 0.32 | 0.32 | 0.32 | 0.32 |
| L-Lysine | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| L-Threonine | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Vitamin premix 3 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Mineral premix 4 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Antioxidant 5 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 |
| Toxin binder 6 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Oxytetracycline 7 | 0 | 0.02 | 0 | 0 | 0 | 0 |
| Ascorbic acid 8 | 0 | 0 | 0.02 | 0 | 0 | 0 |
| RI11 | 0 | 0 | 0 | 0.3 | 0 | 0 |
| RS5 | 0 | 0 | 0 | 0 | 0.3 | 0 |
| UL4 | 0 | 0 | 0 | 0 | 0 | 0.3 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 |
| Calculated nutrient level 9 | ||||||
| Metabolizable energy kcal/kg | 3140.58 | 3140.57 | 3140.57 | 3140.57 | 3140.57 | 3140.57 |
| Protein % | 19.68 | 19.68 | 19.68 | 19.68 | 19.68 | 19.68 |
| Fat % | 8.319 | 8.319 | 8.319 | 8.319 | 8.319 | 8.319 |
| Fiber % | 3.973 | 3.973 | 3.973 | 3.973 | 3.973 | 3.973 |
| Calcium % | 1.097 | 1.097 | 1.097 | 1.097 | 1.097 | 1.097 |
| Total phosphorus % | 0.848 | 0.848 | 0.848 | 0.848 | 0.848 | 0.848 |
| Available phosphorus for poultry % | 0.49 | 0.49 | 0.49 | 0.49 | 0.49 | 0.49 |
| Target Microbes | Primer Sequence 5′-3′ | Product Size (bp) | References |
|---|---|---|---|
| Total bacteria | F—CGGCAACGAGCGCAACCC | 145 | [58] |
| R—CCATTGTAGCACGTGTGTAGCC | |||
| Lactobacillus | F—CATCCAGTGCAAACCTAAGAG | 341 | [59] |
| R—GATCCGCTTGCCTTCGCA | |||
| Bifidobacterium | F—GGGTGGTAATGCCGGATG | 278 | [60] |
| R—TAAGCCATGGACTTTCACACC | |||
| Enterococcus genus | F—CCCTTATTGTTAGTTGCCATCATT | 144 | [58] |
| R—ACTCGTTGTACTTCCCATTGT | |||
| Enterobacteriaceae | F—CATTGACGTTACCCGCAGAAGAAGC | 195 | [59] |
| R—CTCTACGAGACTCAAGCTTGC | |||
| Escherichia coli | F—GTGTGATATCTACCCGCTTCGC | 82 | [59] |
| R—AGAACGCTTTGTGGTTAATCAGGA | |||
| Salmonella | F—TCGTCATTCCATTACCTACC | 119 | [61] |
| R—AAACGTTGAAAAACTGAGGA |
| Target Gene | Primer Sequence 5′-3′ | Product Size (bp) | Reference |
|---|---|---|---|
| GHR | F—AACACAGATACCCAACAGCC | 145 | [62] |
| R—AGAAGTCAGTGTTTGTCAGGG | |||
| IGF-1 | F—CACCTAAATCTGCACGCT | 140 | [62] |
| R—CTTGTGGATGGCATGATCT | |||
| GAPDH | F—CTGGCAAAGTCCAAGTGGTG | 312 | [63] |
| R—AGCACCACCCTTCAGATGAG |
| Parameters | Dietary Treatments 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| IBW (g) | 1020.31 | 1027.5 | 1001.29 | 1014 | 1004.48 | 1006.81 | 5.77 | 0.77 |
| FBW (g) | 2705.11 b | 2717.52 b | 2735.52 b | 2951.75 a | 2826.03 a,b | 2834.00 a,b | 18.92 | 0.001 |
| CWG (g) | 1704.21 b | 1746.54 b | 1759.34 b | 1944.94 a | 1802.77 b | 1805.39 b | 16.44 | 0.001 |
| ADG (g) | 81.15 b | 83.16 b | 83.77 b | 92.61 a | 85.84 b | 85.97 b | 0.78 | 0.001 |
| CFI (g) | 2932.1 | 3032.1 | 2970.8 | 3109.9 | 3004 | 3009 | 33.56 | 0.77 |
| FCR (g/g) | 1.72 a | 1.72 a | 1.70 a,b | 1.61 c | 1.68 b | 1.67 b | 0.007 | 0.001 |
| Mortality | 14/42 | 17/42 | 13/42 | 10/42 | 12/42 | 11/42 | - | 0.636 |
| Parameters | Dietary Treatments 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| Carcass weight (g) | 1905.9 c | 1956.9 c | 2059.6 b,c | 2350.4 a | 2222.3 a,b | 2306.7 a | 78.37 | 0.001 |
| Carcass % | 72.78 | 73.74 | 74.32 | 75.67 | 74.57 | 75.85 | 1.21 | 0.56 |
| Breast % | 27.11 | 28.51 | 27.07 | 28.82 | 26.75 | 28.33 | 0.98 | 0.56 |
| Leg % | 20.05 | 18.51 | 22.18 | 20.33 | 20.54 | 21.35 | 0.75 | 0.15 |
| Wing % | 7.24 | 7.52 | 8.04 | 7.61 | 7.53 | 7.68 | 0.27 | 0.59 |
| Back % | 18.12 | 16.81 | 18.02 | 18.39 | 19.35 | 17.88 | 0.74 | 0.33 |
| Liver % | 1.97 | 1.86 | 1.84 | 1.85 | 1.86 | 1.82 | 0.06 | 0.75 |
| Gizzard % | 2.16 | 2.15 | 2.02 | 1.96 | 1.91 | 1.82 | 0.13 | 0.35 |
| Spleen % | 0.08 | 0.09 | 0.06 | 0.08 | 0.07 | 0.06 | 0.01 | 0.62 |
| Abdominal fat % | 1.28 | 1.23 | 1.15 | 1.11 | 1.15 | 1.05 | 0.13 | 0.87 |
| Heart % | 0.38 | 0.4 | 0.43 | 0.4 | 0.4 | 0.38 | 0.03 | 0.88 |
| Parameters | Dietary Treatments 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| Villi height, μm | ||||||||
| Duodenum | 1178.5 c | 1266.08 b,c | 1310.1 b | 1577.57 a | 1368.24 b | 1291.15 b | 20.99 | <0.0001 |
| Jejunum | 893.71 d | 1000.45 c | 1058.58 b,c | 1117.61 a,b | 1142.34 a | 1183.97 a | 15.5 | <0.0001 |
| Ileum | 611.43 d | 702.84 c | 839.08 b | 932.65 a | 886.69 a,b | 885.7 a,b | 17.11 | <0.0001 |
| Crypt depth, μm | ||||||||
| Duodenum | 239.76 a | 195.05 b,c | 180.67 b,c | 211.18 a,b | 174.45 c | 204.54 b,c | 5.02 | 0.001 |
| Jejunum | 143.64 | 135.51 | 142 | 147.45 | 142.18 | 145.49 | 14.49 | 0.945 |
| Ileum | 127.55 a | 100.77 b | 104.00 b | 98.19 b | 102.40 b | 111.94 a,b | 2.59 | 0.007 |
| Villi height: Crypt depth | ||||||||
| Duodenum | 4.9 c | 6.60 b | 7.40 a,b | 7.94 a | 8.16 a | 6.71 b | 0.20 | <0.0001 |
| Jejunum | 6.35 b | 7.82 a | 7.82 a | 7.72 a | 8.26 a | 8.29 a | 0.19 | 0.040 |
| Ileum | 4.95 c | 6.96 b | 8.45 a | 9.58 a | 8.71 a | 8.34 a | 0.26 | <0.0001 |
| Parameters | Dietary Treatments 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| NC | PC | AA | RI11 | RS5 | UL4 | |||
| Total bacteria | 9.65 c | 9.90b c | 10.18 a,b | 10.25 a | 10.13 a,b | 10.23 a | 0.04 | 0.004 |
| Lactobacillus | 7.79 c | 7.90b c | 8.07 b,c | 8.65 a | 8.26 b | 8.19 b | 0.06 | 0.002 |
| Bifidobacterium | 4.48 b | 5.87 a | 6.29 a | 6.56 a | 6.48 a | 6.46 a | 0.17 | 0.001 |
| Enterobacteriaceae | 7.57 a | 7.21 a,b | 6.35 c | 6.34 c | 6.70 b,c | 6.67 b,c | 0.11 | 0.001 |
| Escherichia coli | 7.14a b | 7.71 a | 6.91 b | 6.67 b | 6.82 b | 7.07 b | 0.09 | 0.008 |
| Enterococcus | 7.71 | 8.04 | 8.23 | 8.33 | 8.39 | 8.12 | 0.09 | 0.308 |
| Salmonella | 3.26 a | 2.77 a | 2.38 a,b | 1.77 b | 2.39 a,b | 2.46 a,b | 0.13 | 0.034 |
| Caecal pH | 6.14 a | 6.02 a,b | 5.91 b,c | 5.83 c | 5.88 b,c | 5.91 b,c | 0.03 | 0.009 |
| Lactobacillus | Bifidobacterium | Enterobacteriaceae | E. coli | Salmonella | Caecum pH | |
|---|---|---|---|---|---|---|
| Lactobacillus | 0.34 * | −0.33 * | −0.38 * | −0.37 * | −0.50 *** | |
| Bifidobacterium | −0.38 * | −0.23 | −0.34 * | −0.55 *** | ||
| Enterobacteriaceae | 0.69 *** | 0.59 *** | 0.45 ** | |||
| E. coli | 0.32 * | 0.33 * | ||||
| Salmonella | 0.48 *** | |||||
| Caecum pH |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Mustapha, N.M.; Zulkifli, I.; Izuddin, W.I. Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress. Animals 2019, 9, 644. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9090644
Humam AM, Loh TC, Foo HL, Samsudin AA, Mustapha NM, Zulkifli I, Izuddin WI. Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress. Animals. 2019; 9(9):644. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9090644
Chicago/Turabian StyleHumam, Ali Merzza, Teck Chwen Loh, Hooi Ling Foo, Anjas Asmara Samsudin, Noordin Mohamed Mustapha, Idrus Zulkifli, and Wan Ibrahim Izuddin. 2019. "Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress" Animals 9, no. 9: 644. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9090644