
Antifungal Activity and Chemical Composition of Seven Essential Oils to Control the Main Seedborne Fungi of Cucurbits


 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
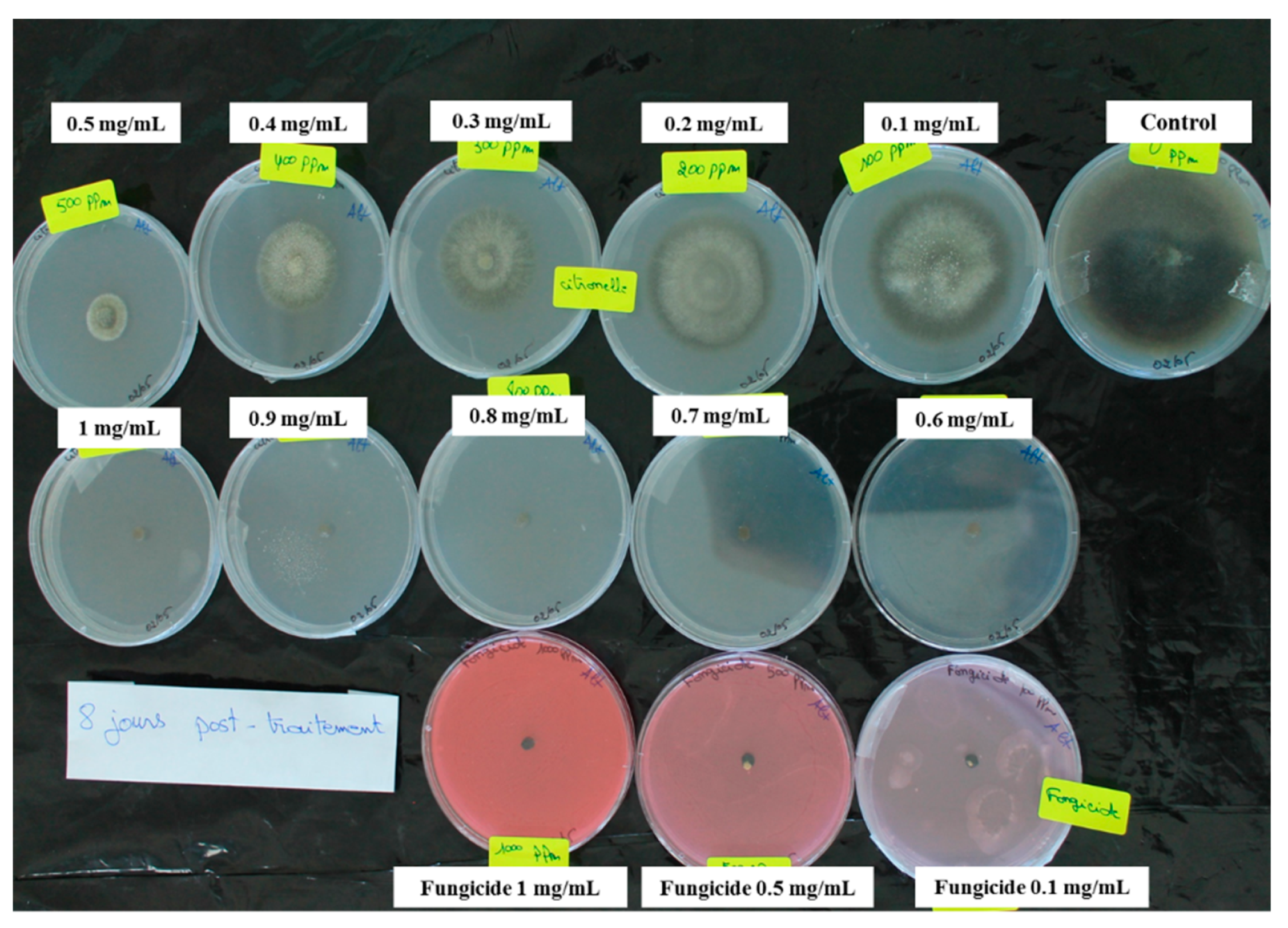
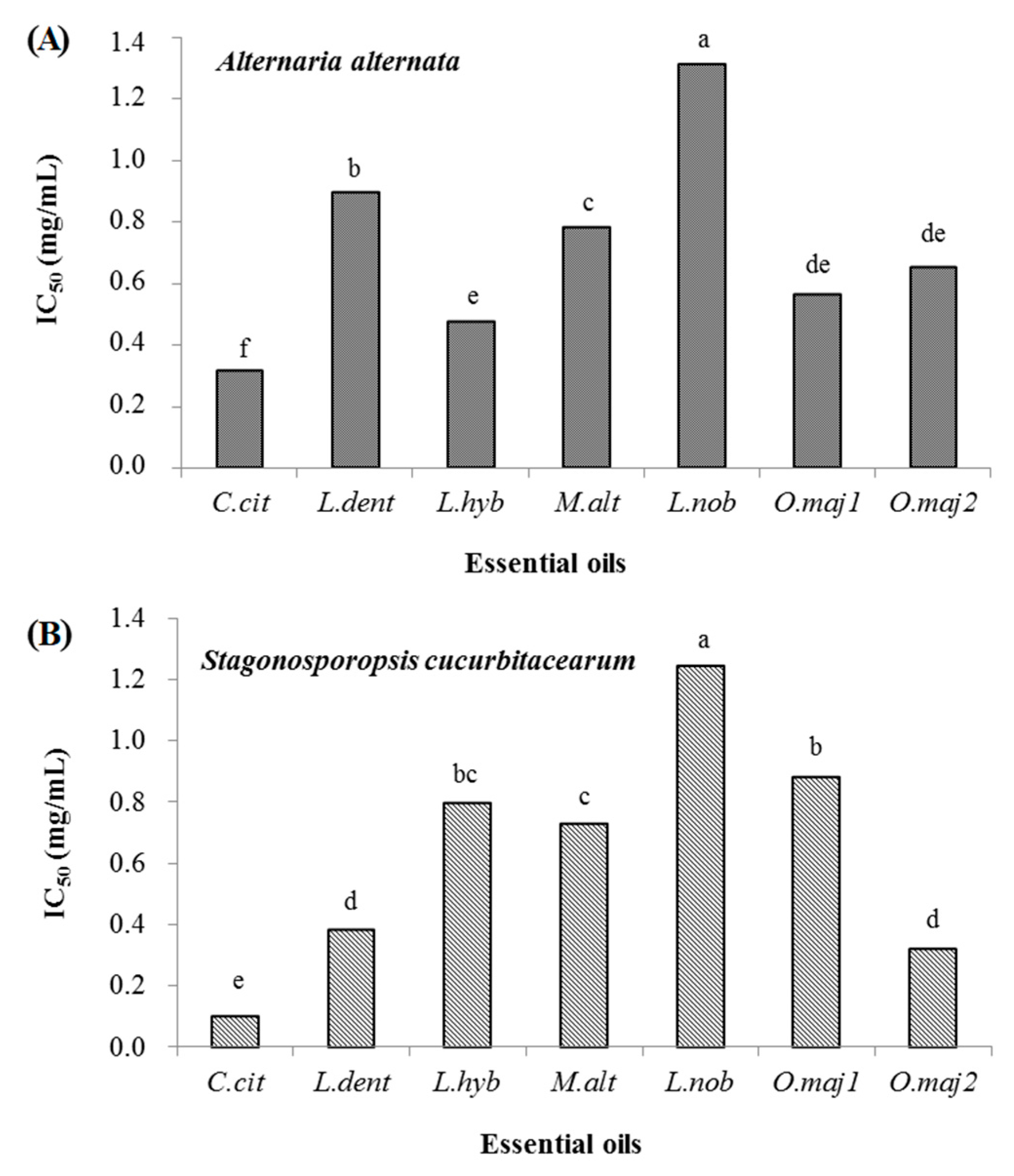
2.1. In Vitro Inhibition of Fungal Growth by the Seven Essential Oils
2.2. Chemical Profiles of the Essential Oils
3. Discussion
4. Materials and Methods
4.1. Origin of the Essential Oils
4.2. Fungal Strains
4.3. In Vitro Antifungal Activities on Mycelial Growth
4.4. Gas Chromatography-Mass Spectrometry Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT Data. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 27 October 2020).
- Moumni, M.; Allagui, M.B.; Mancini, V.; Murolo, S.; Tarchoun, N.; Romanazzi, G. Morphological and molecular identification of seedborne fungi in squash (Cucurbita maxima, Cucurbita moschata). Plant Dis. 2020, 104, 1335–1350. [Google Scholar] [CrossRef] [PubMed]
- Moumni, M.; Mancini, V.; Allagui, M.B.; Murolo, S.; Romanazzi, G. Black rot of squash (Cucurbita moschata Duchesne) caused by Stagonosporopsis cucurbitacearum reported in Italy. Phytopathol. Mediterr. 2019, 58, 381–385. [Google Scholar] [CrossRef]
- Yao, X.; Li, P.; Xu, J.; Zhang, M.; Ren, R.; Liu, G.; Yang, X. Rapid and sensitive detection of Didymella bryoniae by visual loop-mediated isothermal amplification assay. Front. Microbiol. 2016, 7, 1372. [Google Scholar] [CrossRef] [PubMed]
- Nuangmek, W.; Aiduang, W.; Suwannarach, N.; Kumla, J.; Lumyong, S. First report of gummy stem blight caused by Stagonosporopsis cucurbitacearum on cantaloupe in Thailand. Can. J. Plant Pathol. 2018, 40, 306–311. [Google Scholar] [CrossRef]
- Töfoli, J.G.; Domingues, R.J. Alternarioses in vegetables: Symptoms, etiology, and integrated management. Biológico 2004, 66, 23–33. [Google Scholar] [CrossRef]
- Farrag, E.S.H.; Moharam, M.H.A. Pathogenic fungi transmitted through cucumber seeds and safely elimination by application of peppermint extract and oil. Not. Sci. Biol. 2012, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Boughalleb, N.; El Mahjoub, M. In vitro determination of Fusarium spp. infection on watermelon seeds and their localization. Plant Pathol. J. 2006, 5, 178–182. [Google Scholar] [CrossRef]
- Gannibal, P.B. Alternaria cucumerina causing leaf spot of pumpkin newly reported in North Caucasus (Russia). New Dis. Rep. 2011, 23, 36. [Google Scholar] [CrossRef] [Green Version]
- Fish, W.W.; Bruton, B.D.; Popham, T.W. Cucurbit host range of Myrothecium roridum isolated from watermelon. Am. J. Plant Sci. 2012, 3, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Sultana, N.; Ghaffar, A. Pathogenesis and control of Myrothecium spp., the cause of leaf spot on bitter gourd (Momordica charantia Linn.). Pak. J. Bot. 2009, 1, 429–433. [Google Scholar]
- Boughalleb, N.; Tarchoun, N.; Dallagi, W. Effect of fungicides on in vitro infestation level of radish, carrot and pepper seeds. Plant Pathol. J. 2006, 3, 388–392. [Google Scholar] [CrossRef]
- Yates, I.E.; Arnold, J.W.; Hinton, D.M.; Basinger, W.; Walcott, R.R. Fusarium verticillioides induction of maize seed rot and its control. Can. J. Bot. 2003, 81, 422–428. [Google Scholar] [CrossRef]
- Keinath, A.P. Effect of protectant fungicide application schedules on gummy stem blight epidemics and marketable yield of watermelon. Plant Dis. 2000, 84, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Keinath, A.P.; DuBose, V.B.; May, M.H.; Latin, R.X. Comparison of seven fungicides intervals to control gummy stem blight in a fall watermelon crop. Fungic. Nematic. Tests 1998, 53, 268. [Google Scholar]
- Johnson, C.E.; Payne, J.T.; Buckley, J.B. Evaluation of fungicides for gummy stem blight control on watermelon. Fungic. Nematic. Tests 1995, 50, 184. [Google Scholar]
- Sudisha, J.; Niranjana, S.R.; Umesha, S.; Prakash, H.S.; Shetty, H.S. Transmission of seed-borne infection of muskmelon by Didymella bryoniae and effect of seed treatments on disease incidence and fruit yield. Biol. Control 2006, 37, 196–205. [Google Scholar] [CrossRef]
- Barnard, M.; Padgitt, M.; Uri, N.D. Pesticide use and its measurement. Int. Pest Control 1997, 39, 161–164. [Google Scholar]
- Misra, G.; Pavlostathis, S.G. Biodegradation kinetics of monoterpenes in liquid and soil-slurry systems. Appl. Microbiol. Biotechnol. 1997, 47, 572–577. [Google Scholar] [CrossRef]
- Slamet, A.S.; Nakayasu, A.; Bai, H. The determinants of organic vegetable purchasing in Jabodetabek region, Indonesia. Foods 2016, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Antunes, M.D.C.; Cavaco, A.M. The use of essential oils for postharvest decay control. Flavour Fragr. J. 2010, 25, 351–366. [Google Scholar] [CrossRef]
- Shahi, S.K.; Patra, M.; Shukla, A.C.; Dikshit, A. Use of essential oil as botanical-pesticide against postharvest spoilage in Malus pumilo fruits. Biocontrol 2003, 48, 223–232. [Google Scholar] [CrossRef]
- Djioua, T.; Charles, F.; Freire, M., Jr.; Filgueiras, H.; Ducamp-Collin, M.; Sallanon, H. Combined effects of postharvest heat treatment and chitosan coating on quality of fresh-cut mangoes (Mangifera indica L.). Int. J. Food Sci. Technol. 2010, 45, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Mancini, V.; Romanazzi, G. Seed treatments to control seedborne fungal pathogens of vegetable crops. Pest Manage. Sci. 2014, 70, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Kishore, G.K.; Pande, S. Natural fungicides for management of phytopathogenic fungi. Annu. Rev. Plant Pathol. 2004, 3, 331–356. [Google Scholar]
- Terzi, V.; Morcia, C.; Faccioli, P.; Vale, G.; Tacconi, G.; Malnati, M. In vitro antifungal activity of the tea tree (Melaleuca alternifolia) essential oil and its major components against plant pathogens. Lett. Appl. Microbiol. 2007, 44, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, L.; Orzali, L. Activity of tea tree (Melaleuca alternifolia, Cheel) and thyme (Thymus vulgaris, Linnaeus.) essential oils against some pathogenic seed borne fungi. J. Essent. Oil Res. 2011, 23, 43–47. [Google Scholar] [CrossRef]
- Seixas, P.T.L.; Castro, H.G.; Cardoso, D.P.; Junior, A.F.C.; do Nascimento, I.R. Bioactivity of essential oils on the fungus Didymella bryoniae of the cucumber culture. Appl. Res. Agrotechnol. 2012, 5, 61–66. [Google Scholar] [CrossRef]
- Shafique, S.; Majeed, R.A.; Shafique, S. Cymbopogon citrates: A remedy to control selected Alternaria species. J. Med. Plant Res. 2012, 6, 1879–1885. [Google Scholar] [CrossRef]
- Fiori, A.C.G.; Schwan-Estrada, K.R.F.; Stangarlin, J.R.; Vida, J.B.; Scapim, C.A.; Cruz, M.E.S.; Pascholati, S.F. Antifungal activity of leaf extracts and essential oils of some medicinal plants against Didymella bryoniae. J. Phytopathol. 2000, 148, 483–487. [Google Scholar] [CrossRef]
- Jie, Z.; ChaoYing, Z.; ZhanFang, G.; Lei, Y. Inhibititory activity of Cymbopogon citratus essential oil against nine phytopathogens. J. Shanghai Jiaotong Univ.-Agric. Sci. 2011, 29, 72–74. [Google Scholar]
- Guimarães, L.G.L.; dasGraças Cardoso, M.; Souza, P.E.; de Andrade, J.; Vieira, S.S. Antioxidant and fungitoxic activities of the lemongrass essential oil and citral. Rev. Cienc. Agron. 2011, 42, 464. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Ramirez, L.A.; Gutiérrez-Pacheco, M.M.; Vargas-Arispuro, I.; González-Aguilar, G.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Inhibition of glucosyltransferase activity and glucan production as an antibiofilm mechanism of lemongrass essential oil against Escherichia coli O157: H7. Antibiotics 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plata-Rueda, A.; Martínez, L.C.; da Silva Rolim, G.; Coelho, R.P.; Santos, M.H.; de Souza Tavares, W.; Zanuncio, J.C.; Serrão, J.E. Insecticidal and repellent activities of Cymbopogon citratus (Poaceae) essential oil and its terpenoids (citral and geranyl acetate) against Ulomoides dermestoides. Crop Prot. 2020, 137, 105299. [Google Scholar] [CrossRef]
- Bermúdez-Vásquez, M.J.; Granados-Chinchilla, F.; Molina, A. Composición química y actividad antimicrobiana del aceite esencial de Psidium guajava y Cymbopogon citratus. Agron. Mesoam. 2019, 30, 147–163. [Google Scholar] [CrossRef]
- Dègnon, R.G.; Allagbé, A.C.; Adjou, E.S.; Dahouenon-Ahoussi, E. Antifungal activities of Cymbopogon citratus essential oil against Aspergillus species isolated from fermented fish products of Southern Benin. J. Food Qual. Hazards Control 2019, 6, 53–57. [Google Scholar] [CrossRef]
- Supardan, M.D.; Misran, E.; Mustapha, W.A.W. Effect of material length on kinetics of essential oil hydrodistillation from lemongrass (Cymbopogon citratus). J. Eng. Sci. Technol. 2019, 14, 810–819. [Google Scholar]
- Brügger, B.P.; Martínez, L.C.; Plata-Rueda, A.; de Castro e Castro, B.M.; Soares, M.A.; Wilcken, C.F.; Carvalho, A.G.; Serrão, J.E.; Zanuncio, J.C. Bioactivity of the Cymbopogon citratus (Poaceae) essential oil and its terpenoid constituents on the predatory bug, Podisus nigrispinus (Heteroptera: Pentatomidae). Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Macedo, I.T.F.; de Oliveira, L.M.B.; André, W.P.P.; de Araújo Filho, J.V.; dos Santos, J.M.L.; Rondon, F.C.M.; Ribeiro, W.L.C.; Camurça-Vasconcelos, A.L.F.; de Oliveira, E.F.; de Paula, H.C.B.; et al. Anthelmintic effect of Cymbopogon citratus essential oil and its nanoemulsion on sheep gastrointestinal nematodes. Rev. Bras. Parasitol. Vet. 2019, 28, 522–527. [Google Scholar] [CrossRef]
- Sonker, N.; Pandey, A.K.; Singh, P.; Tripathi, N.N. Assessment of Cymbopogon citratus (DC.) stapf essential oil as herbal preservatives based on antifungal, antiaflatoxin, and antiochratoxin activities and in vivo efficacy during storage. J. Food Sci. 2014, 79, M628–M634. [Google Scholar] [CrossRef]
- Dalcin, M.S.; Cafee-Filho, A.C.; de Almeida Sarmento, R.; do Nascimento, I.R.; de Souza Ferreira, T.P.; de Sousa Aguiar, R.W.; dos Santos, G.R. Evaluation of essential oils for preventive or curative management of melon gummy stem blight and plant toxicity. J. Med. Plant Res. 2017, 11, 426–432. [Google Scholar] [CrossRef] [Green Version]
- Kishore, G.K.; Pande, S.; Harish, S. Evaluation of essential oils and their components for broad-spectrum antifungal activity and control of late leaf spot and crown rot diseases in peanut. Plant Dis. 2007, 91, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farias, P.K.S.; Silva, J.C.R.L.; Souza, C.N.D.; Fonseca, F.S.A.D.; Brandi, I.V.; Martins, E.R.; Azevedo, A.M.; Almeida, A.C.D. Antioxidant activity of essential oils from condiment plants and their effect on lactic cultures and pathogenic bacteria. Cienc. Rural 2019, 49, e20180140. [Google Scholar] [CrossRef] [Green Version]
- Kurita, N.; Makoto, M.; Kurane, R.; Takahara, Y.; Ichimura, K. Antifungal activity of components of essential oils. Agric. Biol. Chem. 1981, 45, 945–952. [Google Scholar] [CrossRef]
- de Paula Martins, R.; da Silva Gomes, R.A.; Malpass, A.C.G.; Okura, M.H. Chemical characterization of Lavandula dentata L. essential oils grown in Uberaba-MG. Cienc. Rural 2019, 49. [Google Scholar] [CrossRef] [Green Version]
- Bajalan, I.; Rouzbahani, R.; Pirbalouti, A.G.; Maggi, F. Chemical composition and antibacterial activity of Iranian lavandula × hybrida. Chem. Biodivers. 2017, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dias, I.J.; Trajano, E.R.I.S.; Castro, R.D.; Ferreira, G.L.S.; Medeiros, H.C.M.; Gomes, D.Q.C. Antifungal activity of linalool in cases of Candida spp. isolated from individuals with oral candidiasis. Braz. J. Biol. 2018, 78, 368–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitarokili, D.; Couladis, M.; Petsikos-Panayotarou, N.; Tzakou, O. Composition and antifungal activity on soil-borne pathogens of the essential oil of Salvia sclarea from Greece. J. Agr. Food Chem. 2002, 50, 6688–6691. [Google Scholar] [CrossRef] [PubMed]
- Chaves, R.D.S.B.; Martins, R.L.; Rodrigues, A.B.L.; Rabelo, É.D.M.; Farias, A.L.F.; Brandão, L.B.; Santos, L.L.; Galardo, A.K.R.; de Almeida, S.S.M.D.S. Evaluation of larvicidal potential against larvae of Aedes aegypti (Linnaeus, 1762) and of the antimicrobial activity of essential oil obtained from the leaves of Origanum majorana L. PLoS ONE 2020, 15, e0235740. [Google Scholar] [CrossRef]
- Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in Cilento (Southern Italy). Molecules 2019, 24, 2576. [Google Scholar] [CrossRef] [Green Version]
- Khadhri, A.; Bouali, I.; Aouadhi, C.; Lagel, M.C.; Masson, E.; Pizzi, A. Determination of phenolic compounds by MALDI–TOF and essential oil composition by GC–MS during three development stages of Origanum majorana L. Biomed. Chromatogr. 2019, 33, 1–13. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Aouni, M.; Gharsallah, N.; Kadri, A. Chemical composition and in vitro evaluation of antioxidant, antimicrobial, cytotoxicity and anti-acetylcholinesterase properties of Tunisian Origanum majorana L. essential oil. Microb. Pathog. 2016, 95, 86–94. [Google Scholar] [CrossRef]
- Busatta, C.; Barbosa, J.; Cardoso, R.I.; Paroul, N.; Rodrigues, M.; de Oliveira, D.; de Oliveira, J.V.; Cansian, R.L. Chemical profiles of essential oils of marjoram (Origanum majorana) and oregano (Origanum vulgare) obtained by hydrodistillation and supercritical CO2. J. Essent. Oil Res. 2017, 29, 367–374. [Google Scholar] [CrossRef]
- Fikry, S.; Khalil, N.; Salama, O. Chemical profiling, biostatic and biocidal dynamics of Origanum vulgare L. essential oil. AMB Express 2019, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Mir, J.I.; Shafi, W.; Faktoo, S.Z.; Singh, D.B.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Divergence in tissue-specific expression patterns of genes associated with the terpeniod biosynthesis in two oregano species Origanum vulgare L., and Origanum majorana. Ind. Crops Prod. 2018, 123, 546–555. [Google Scholar] [CrossRef]
- Nardoni, S.; Pisseri, F.; Pistelli, L.; Najar, B.; Luini, M.; Mancianti, F. In vitro activity of 30 essential oils against bovine clinical isolates of Prototheca zopfii and Prototheca blaschkeae. Vet. Sci. 2018, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Semiz, G.; Semiz, A.; Mercan-Doğan, N. Essential oil composition, total phenolic content, antioxidant and antibiofilm activities of four Origanum species from southeastern turkey. Int. J. Food Prop. 2018, 21, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Sahab, A.F.; Aly, S.; Hathout, A.S.; Ziedan, E.S.H.; Sabry, B.A. Application of some plant essential oils to control Fusarium isolates associated with freshly harvested maize in Egypt. J. Essent. Oil Bear. Plants 2014, 17, 1146–1155. [Google Scholar] [CrossRef]
- Szczerbanik, M.; Jobling, J.; Morris, S.; Holford, P. Essential oil vapours control some common postharvest fungal pathogens. Aust. J. Exp. Agric. 2007, 47, 103–109. [Google Scholar] [CrossRef]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. 2012, 29 Part A, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Ebani, V.V.; Najar, B.; Bertelloni, F.; Pistelli, L.; Mancianti, F.; Nardoni, S. Chemical composition and in vitro antimicrobial efficacy of sixteen essential oils against Escherichia coli and Aspergillus fumigatus isolated from poultry. Vet. Sci. 2018, 5, 62. [Google Scholar] [CrossRef] [Green Version]
- Elmi, A.; Ventrella, D.; Barone, F.; Carnevali, G.; Filippini, G.; Pisi, A.; Benvenuti, S.; Scozzoli, M.; Bacci, M.L. In vitro effects of tea tree oil (Melaleuca alternifolia essential oil) and its principal component terpinen-4-ol on swine spermatozoa. Molecules 2019, 24, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Flores, P.G.; Pérez-López, L.A.; Rivas-Galindo, V.M.; Paniagua-Vega, D.; Galindo-Rodríguez, S.A.; Álvarez-Román, R. Simultaneous GC-FID quantification of main components of Rosmarinus officinalis L. and Lavandula dentata essential oils in polymeric nanocapsules for antioxidant application. J. Anal. Meth. Chem. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhifi, W.; Bellili, S.; Jazi, S.; Nasr, S.B.; El Beyrouthy, M.; Mnif, W. Phytochemical composition and antioxidant activity of Tunisian Laurus nobilis. Pak. J. Pharm. Sci. 2018, 31, 2397–2402. [Google Scholar]
- Alves, F.M.; De, F.; França, K.R.D.S.; Araújo, I.G.D.; Nóbrega, L.P.D.; Xavier, A.L.; Dos, S.; Lima, T.S.; Rodrigues, A.P.M.; Júnior, A.F.; et al. Control of Alternaria alternata using melaleuca essential oil (Melaleuca alternifolia). J. Exp. Agric. Int. 2019, 40, 1–10. [Google Scholar] [CrossRef]
- Black-Solis, J.; Ventura-Aguilar, R.I.; Correa-Pacheco, Z.; Corona-Rangel, M.L.; Bautista-Baños, S. Preharvest use of biodegradable polyester nets added with cinnamon essential oil and the effect on the storage life of tomatoes and the development of Alternaria alternata. Sci. Hortic. 2019, 245, 65–73. [Google Scholar] [CrossRef]
- Soylu, E.M.; Kose, F. Antifungal activities of essential oils against citrus black rot disease agent Alternaria alternata. J. Essent. Oil-Bear. Plants 2015, 18, 894–903. [Google Scholar] [CrossRef]
- Najar, B.; Marchioni, I.; Ruffoni, B.; Copetta, A.; Pistelli, L.; Pistelli, L. Volatilomic analysis of four edible flowers from Agastache genus. Molecules 2019, 24, 4480. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 1902. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavor and Fragrance Volatiles by Glass Capillary Gas Chromatography, Food/Nahrung; Academic Press: New York, NY, USA; London, UK; Sydney, Australia; Toronto, ON, Canada; San Francisco, CA, USA, 1982. [Google Scholar]
- Masada, Y. Analysis of Essential Oils by Gas Chromatography and Mass Spectrometry; John Wiley & Sons, Inc.: New York, NY, USA, 1976. [Google Scholar]
- Stenhagen, E.; Abrahamsson, S.; McLafferty, F.W. Registry of Mass Spectral Data; Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Swigar, A.A.; Silverstein, R.M. Monoterpenes, Aldrich Chemical Company; Aldrich Chemical Company: Milwaukee, WI, USA, 1981. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Code | Species | Family | Common Name | Source |
|---|---|---|---|---|
| C.cit | Cymbopogon citratus (DC.) Stapf | Poaceae | Lemongrass | Biopesticides Laboratory, Regional Centre for Research in Horticulture and Organic Agriculture (CRRHAB), Tunisia |
| L.dent | Lavandula dentata L. | Lamiaceae | Lavender | CRRHAB, Tunisia |
| L.hyb | Lavandula hybrida E.Rev. ex Briq | Lamiaceae | Lavandin | FLORA s.r.l. (Batch N° 161808) |
| M.alt | Melaleuca alternifolia (Maiden & Betche) Cheel | Myrtaceae | Tea tree | FLORA s.r.l. (Batch N° 161960) |
| L.nob | Laurus nobilis L. | Lauraceae | Bay laurel | Medicinal Plants Laboratory, National Institute of Agronomy of Tunisia (INAT) |
| O.maj1 | Origanum majorana L. | Lamiaceae | Marjoram | INAT |
| O.maj2 | Origanum majorana L. | Lamiaceae | Marjoram | CRRHAB, Tunisia |
| Essential Oil | Inhibition of Mycelial Growth of Alternaria alternata (%) at Increasing Essential Oil Concentrations (mg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | 1 | |
| C.cit | 0.00 | 20.28 ± 4.32 | 25.03 ± 1.56 | 43.21 ± 2.33 | 65.92 ± 6.29 | 79.14 ± 10.97 | 100 | 100 | 100 | 100 | 100 |
| L.dent | 0.00 | 9.95 ± 2.30 | 20.07 ± 3.63 | 29.99 ± 4.97 | 44.44 ± 3.13 | 43.82 ± 1.84 | 43.00 ± 3.18 | 37.63 ± 1.65 | 44.44 ± 0.21 | 43.00 ± 1.89 | 54.98 ± 0.83 |
| L.hyb | 0.00 | 31.23 ± 4.40 | 32.88 ± 4.72 | 42.38 ± 2.18 | 50.64 ± 2.89 | 51.05 ± 2.50 | 59.31 ± 3.76 | 63.44 ± 1.64 | 65.72 ± 1.49 | 67.37 ± 0.55 | 71.50 ± 0.95 |
| M.alt | 0.00 | 13.47 ± 5.36 | 15.74 ± 3.06 | 21.52 ± 10.17 | 21.73 ± 5.38 | 24.00 ± 1.09 | 25.44 ± 4.42 | 29.37 ± 3.45 | 62.00 ± 1.49 | 67.99 ± 2.33 | 68.19 ± 2.98 |
| L.nob | 0.00 | 0.58 ± 0.29 | 0.58 ± 0.29 | 1.28 ± 0.41 | 8.72 ± 2.51 | 12.02 ± 1.56 | 23.17 ± 2.48 | 24.00 ± 1.65 | 25.03 ± 3.41 | 33.91 ± 4.96 | 36.08 ± 2.44 |
| O.maj1 | 0.00 | 20.07 ± 2.18 | 37.84 ± 9.68 | 42.59 ± 2.30 | 45.48 ± 4.13 | 47.13 ± 1.80 | 47.75 ± 1.09 | 53.12 ± 7.10 | 60.97 ± 3.41 | 64.27 ± 3.25 | 74.18 ± 1.45 |
| O.maj2 | 0.00 | 27.10 ± 0.55 | 31.85 ± 1.99 | 32.47 ± 1.99 | 33.91 ± 0.55 | 37.01 ± 0.41 | 42.38 ± 1.24 | 42.59 ± 0.83 | 46.92 ± 0.55 | 63.86 ± 1.61 | 90.50 ± 9.50 |
| Fungicides a | 0.00 | 100 | - | - | - | 100 | - | - | - | - | 100 |
| Essential Oil | Inhibition of Mycelial Growth of Stagonosporopsis cucurbitacearum (%) at Increasing Essential Oil Concentrations (mg/mL) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 | 1 | |
| C.cit | 0.00 | 51.76 ± 3.86 | 53.53 ± 1.22 | 58.24 ± 0.59 | 67.25 ± 1.37 | 74.51 ± 3.08 | 85.49 ± 7.45 | 82.16 ± 9.00 | 87.06 ± 6.51 | 100 | 100 |
| L.dent | 0.00 | 29.41 ± 0.00 | 41.18 ± 6.79 | 47.06 ± 1.36 | 51.76 ± 0.68 | 61.37 ± 0.71 | 62.75 ± 0.71 | 66.27 ± 0.39 | 71.37 ± 0.78 | 73.33 ± 0.39 | 73.53 ± 0.59 |
| L.hyb | 0.00 | 5.49 ± 1.68 | 6.27 ± 2.18 | 10.39 ± 2.05 | 12.75 ± 1.04 | 21.57 ± 5.85 | 22.75 ± 4.43 | 27.65 ± 4.75 | 53.92 ± 0.52 | 63.73 ± 0.85 | 73.92 ± 3.63 |
| M.alt | 0.00 | 3.92 ± 1.96 | 22.16 ± 1.04 | 20.78 ± 3.08 | 20.59 ± 3.02 | 30.78 ± 3.16 | 31.37 ± 4.11 | 41.96 ± 3.35 | 60.20 ± 0.71 | 63.92 ± 0.20 | 73.73 ± 0.85 |
| L.nob | 0.00 | 0.98 ± 0.71 | 3.73 ± 3.73 | 5.88 ± 2.38 | 5.88 ± 2.37 | 5.69 ± 0.52 | 6.67 ± 0.71 | 11.37 ± 4.37 | 15.29 ± 4.57 | 29.22 ± 8.35 | 65.29 ± 3.83 |
| O.maj1 | 0.00 | 0.00 | 5.69 ± 1.87 | 8.24 ± 1.22 | 9.02 ± 1.19 | 10.98 ± 1.37 | 33.73 ± 4.31 | 33.73 ± 3.74 | 47.45 ± 6.97 | 54.12 ± 2.96 | 60.00 ± 1.56 |
| O.maj2 | 0.00 | 34.12 ± 4.42 | 49.22 ± 1.41 | 54.31 ± 4.27 | 56.47 ± 1.96 | 57.25 ± 1.56 | 58.82 ± 1.04 | 59.61 ± 1.80 | 61.57 ± 1.19 | 63.53 ± 0.34 | 67.25 ± 1.74 |
| Fungicides a | 0.00 | 75.69 ± 0.00 | - | - | - | 84.90 ± 0.68 | - | - | - | - | 86.67 ± 0.39 |
| Fungus | Fungicidal and Fungistatic Activities of Cymbopogon citratus Essential Oil at Increasing Concentrations (mg/mL) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0.6 | 0.7 | 0.8 | 0.9 | 1 | ||||||
| Fungicidal | Fungistatic | Fungicidal | Fungistatic | Fungicidal | Fungistatic | Fungicidal | Fungistatic | Fungicidal | Fungistatic | |
| Alternaria alternata | NO | Yes | NO | Yes | Yes | NO | Yes | NO | Yes | NO |
| Stagonosporopsis cucurbitacearum | - | - | - | - | - | - | Yes | NO | Yes | NO |
| Compound a | Class | Linear Retention Index | Relative amount (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (n-alkane) b | (Adams, 2007) c | Poaceae | Lamiaceae | Myrtaceae | Lauraceae | ||||||
| C.cit | L.dent | L.hyb | O.maj1 | O.maj2 | M.alt | L.nob | |||||
| 1 | α-Pinene | mh d | 937 | 932 | 0.1 | 0.6 | 0.5 | 0.6 | 0.3 | 2.7 | 5.6 |
| 2 | Sabinene | mh | 974 | 969 | 0.1 | - | 0.1 | 4.5 | 2.2 | - | 6.7 |
| 3 | β-Pinene | mh | 979 | 974 | - | 3.1 | 0.5 | 0.4 | 0.2 | 0.7 | 5.0 |
| 4 | β-Myrcene | mh | 991 | 988 | 5.3 | - | 0.4 | 1.1 | 0.6 | 0.6 | 1.2 |
| 5 | α-Terpinene | mh | 1017 | 1014 | - | - | - | 5.7 | 0.8 | 6.1 | 0.8 |
| 6 | p-Cymene | mh | 1025 | 1020 | 0.3 | 0.9 | 0.2 | 11.3 | 17.8 | 9.3 | 0.6 |
| 7 | Limonene | mh | 1030 | 1024 | 0.4 | 1.1 | 0.8 | 3.5 | 2.4 | 1.0 | 1.7 |
| 8 | Eucalyptol | om | 1032 | 1026 | 0.6 | 63.5 | 6.5 | 0.7 | 0.2 | 2.8 | 47.9 |
| 9 | γ-Terpinene | mh | 1060 | 1054 | - | - | - | 12.6 | 3.8 | 16.0 | 1.4 |
| 10 | Terpinolene | mh | 1088 | 1086 | - | - | 0.2 | 3.4 | 1.2 | 3.0 | 0.3 |
| 11 | Linalool | om | 1099 | 1095 | 0.8 | 1.8 | 33.7 | 1.1 | 3.6 | - | 7.4 |
| 12 | cis-p-Menth-2-en-1-ol | om | 1122 | 1118 | - | - | - | 1.3 | 0.7 | 0.1 | - |
| 13 | trans-Pinocarveol | om | 1139 | 1135 | - | 2.9 | - | - | - | - | - |
| 14 | trans-p-Menth-2-en-1-ol | om | 1141 | 1136 | - | - | - | 1.2 | 0.7 | 0.1 | - |
| 15 | Camphor | om | 1145 | 1141 | - | 0.5 | 9.3 | - | 0.1 | - | - |
| 16 | γ-Terpineol | om | 1166 | 1162 | - | 1.3 | - | - | - | - | 0.3 |
| 17 | endo-Borneol | om | 1167 | 1165 | - | 0.4 | 4.4 | 0.1 | 0.2 | - | - |
| 18 | p-Mentha-1,5-dien-8-ol | om | 1170 | 1166 | 2.5 | - | - | - | - | - | - |
| 19 | Terpinen-4-ol | om | 1177 | 1174 | 0.1 | 1.3 | 4.5 | 32.4 | 50.1 | 41.1 | 1.5 |
| 20 | p-Cymen-8-ol | om | 1183 | 1179 | 1.1 | 0.3 | - | 0.3 | 0.4 | - | - |
| 21 | Cryptone | nt | 1186 | 1183 | - | 1.3 | - | - | - | - | - |
| 22 | α-Terpineol | om | 1189 | 1186 | - | 1.8 | 1.1 | 6.0 | 5.4 | 3.7 | 1.6 |
| 23 | Myrtenal | om | 1198 | 1195 | - | 2.7 | - | - | - | - | - |
| 24 | trans-Piperitol | om | 1208 | 1207 | - | - | - | 1.0 | 0.6 | - | - |
| 25 | β-Citral | om | 1240 | 1235 | 26.0 | - | - | - | - | - | - |
| 26 | Carvone | om | 1243 | 1239 | 0.9 | 1.6 | - | 0.2 | 0.6 | - | - |
| 27 | Geraniol | om | 1253 | 1249 | 2.7 | - | - | - | - | - | - |
| 28 | Linalyl acetate | om | 1257 | 1254 | - | - | 27.7 | 2.7 | 2.2 | - | 0.2 |
| 29 | α-Citral | om | 1270 | 1264 | 51.6 | - | - | - | - | - | - |
| 30 | 2-Undecanone | nt | 1294 | 1293 | 1.2 | - | - | - | - | - | - |
| 31 | 4-Terpinyl acetate | om | 1300 | 1300 | - | - | - | 1.5 | 1.0 | - | - |
| 32 | Lavandulyl acetate | om | 1304 | 1288 | - | - | 3.0 | - | - | - | - |
| 33 | 𝛼-Terpinyl acetate | om | 1350 | 1346 | - | - | - | 0.1 | - | - | 10.2 |
| 34 | Methyleugenol | pp | 1404 | 1403 | - | - | - | - | - | - | 3.1 |
| 35 | β-Caryophyllene | sh | 1419 | 1417 | - | - | 2.0 | 2.4 | 1.6 | 0.5 | 0.5 |
| 36 | Aromandendrene | sh | 1440 | 1439 | - | - | - | - | - | 2.2 | - |
| 37 | β-Selinene | sh | 1486 | 1489 | - | 4.1 | - | - | - | 0.1 | - |
| 38 | Eremophyllene | sh | 1498 | 1492 | - | - | - | - | - | 1.5 | - |
| 39 | δ-Cadinene | sh | 1524 | 1522 | - | - | - | - | - | 1.7 | 0.1 |
| 40 | Caryophyllene oxide | os | 1583 | 1582 | 0.2 | 1.9 | 0.1 | 0.2 | 0.3 | - | - |
| 41 | β-Eudesmol | os | 1651 | 1649 | - | 2.1 | - | - | - | - | - |
| Monoterpene Hydrocarbons | mh | 6.5 | 5.9 | 3.6 | 44.3 | 29.7 | 40.4 | 24.0 | |||
| Oxygenated Monoterpenes | om | 88.5 | 81.1 | 90.8 | 49.7 | 66.8 | 48.1 | 70.3 | |||
| Sesquiterpene Hydrocarbons | sh | - | 5.5 | 4.0 | 3.3 | 2.0 | 9.3 | 1.2 | |||
| Oxygenated Sesquiterpens | os | 0.3 | 4.6 | 0.2 | 0.6 | 1.1 | 1.8 | - | |||
| Penylpropanoids | pp | - | - | - | - | - | - | 3.6 | |||
| Non-terpene Derivatives | nt | 3.1 | 2.2 | 1.4 | - | - | - | 0.4 | |||
| Total Identified | 98.4 | 99.3 | 100.0 | 97.7 | 99.6 | 99.6 | 99.5 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moumni, M.; Romanazzi, G.; Najar, B.; Pistelli, L.; Ben Amara, H.; Mezrioui, K.; Karous, O.; Chaieb, I.; Allagui, M.B. Antifungal Activity and Chemical Composition of Seven Essential Oils to Control the Main Seedborne Fungi of Cucurbits. Antibiotics 2021, 10, 104. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020104
Moumni M, Romanazzi G, Najar B, Pistelli L, Ben Amara H, Mezrioui K, Karous O, Chaieb I, Allagui MB. Antifungal Activity and Chemical Composition of Seven Essential Oils to Control the Main Seedborne Fungi of Cucurbits. Antibiotics. 2021; 10(2):104. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020104
Chicago/Turabian StyleMoumni, Marwa, Gianfranco Romanazzi, Basma Najar, Luisa Pistelli, Hajer Ben Amara, Kaies Mezrioui, Olfa Karous, Ikbal Chaieb, and Mohamed Bechir Allagui. 2021. "Antifungal Activity and Chemical Composition of Seven Essential Oils to Control the Main Seedborne Fungi of Cucurbits" Antibiotics 10, no. 2: 104. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020104