
16S rRNA Sequencing Analysis of the Gut Microbiota in Broiler Chickens Prophylactically Administered with Antimicrobial Agents

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
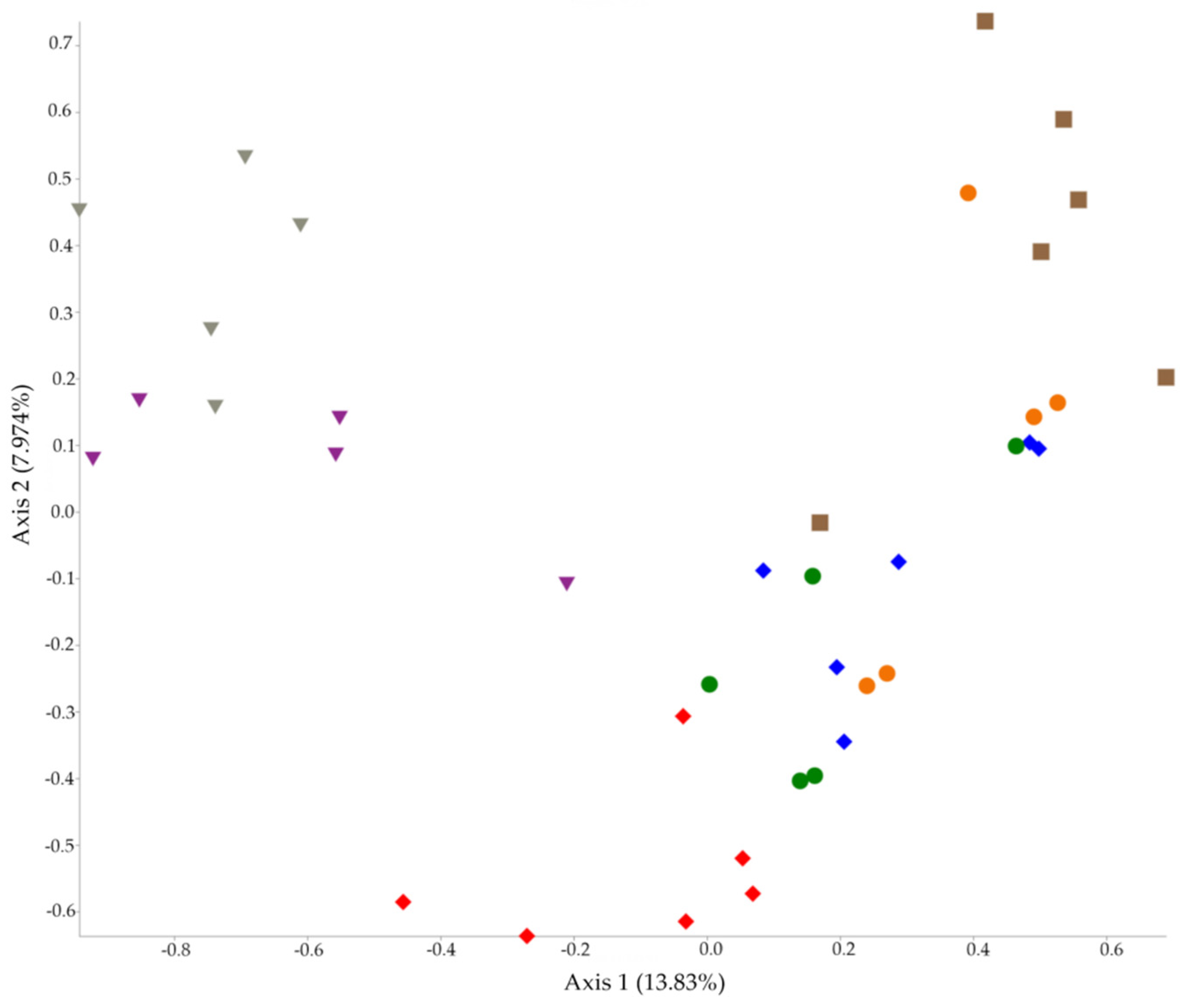
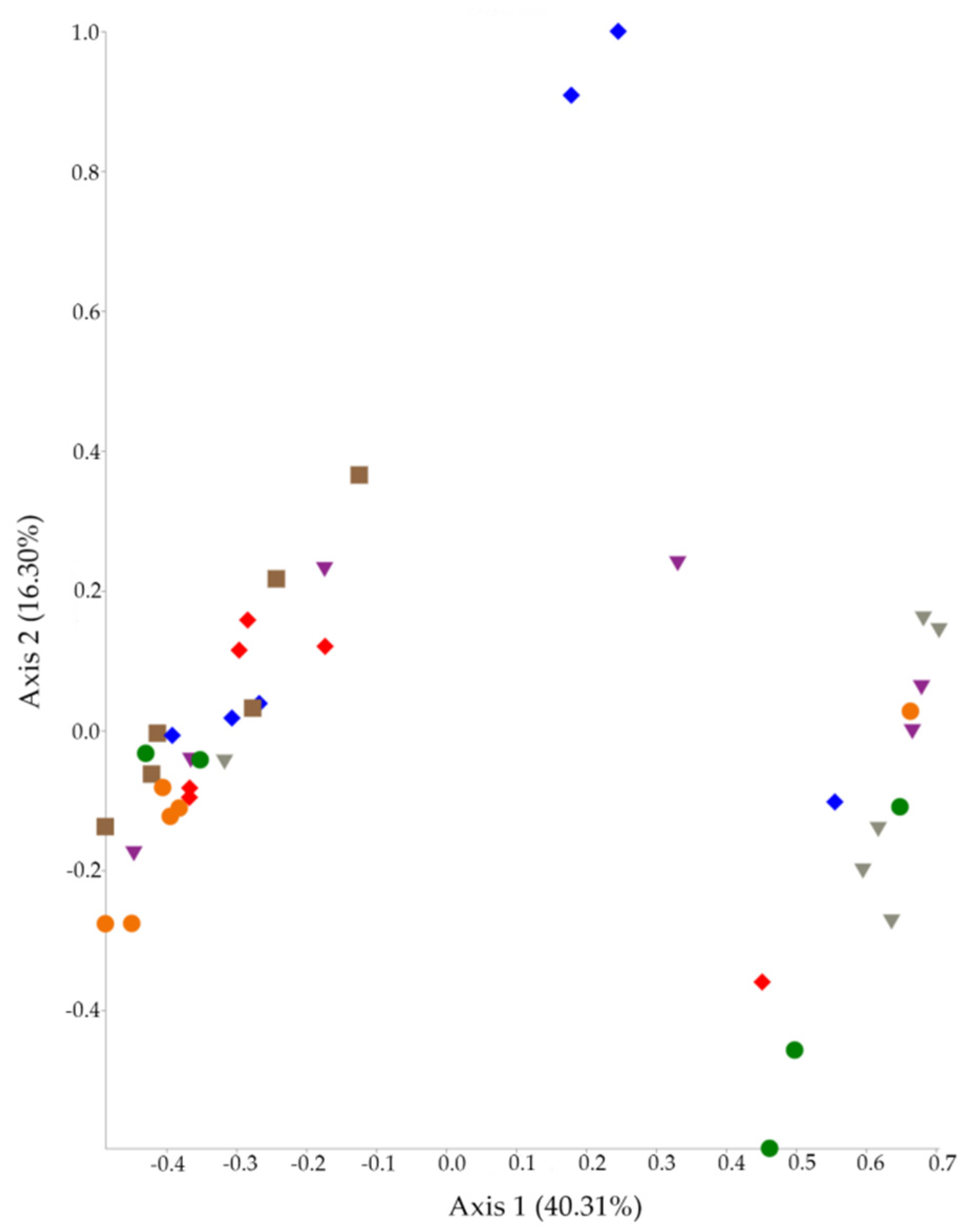
2.1. 16S rRNA Diversity Analysis
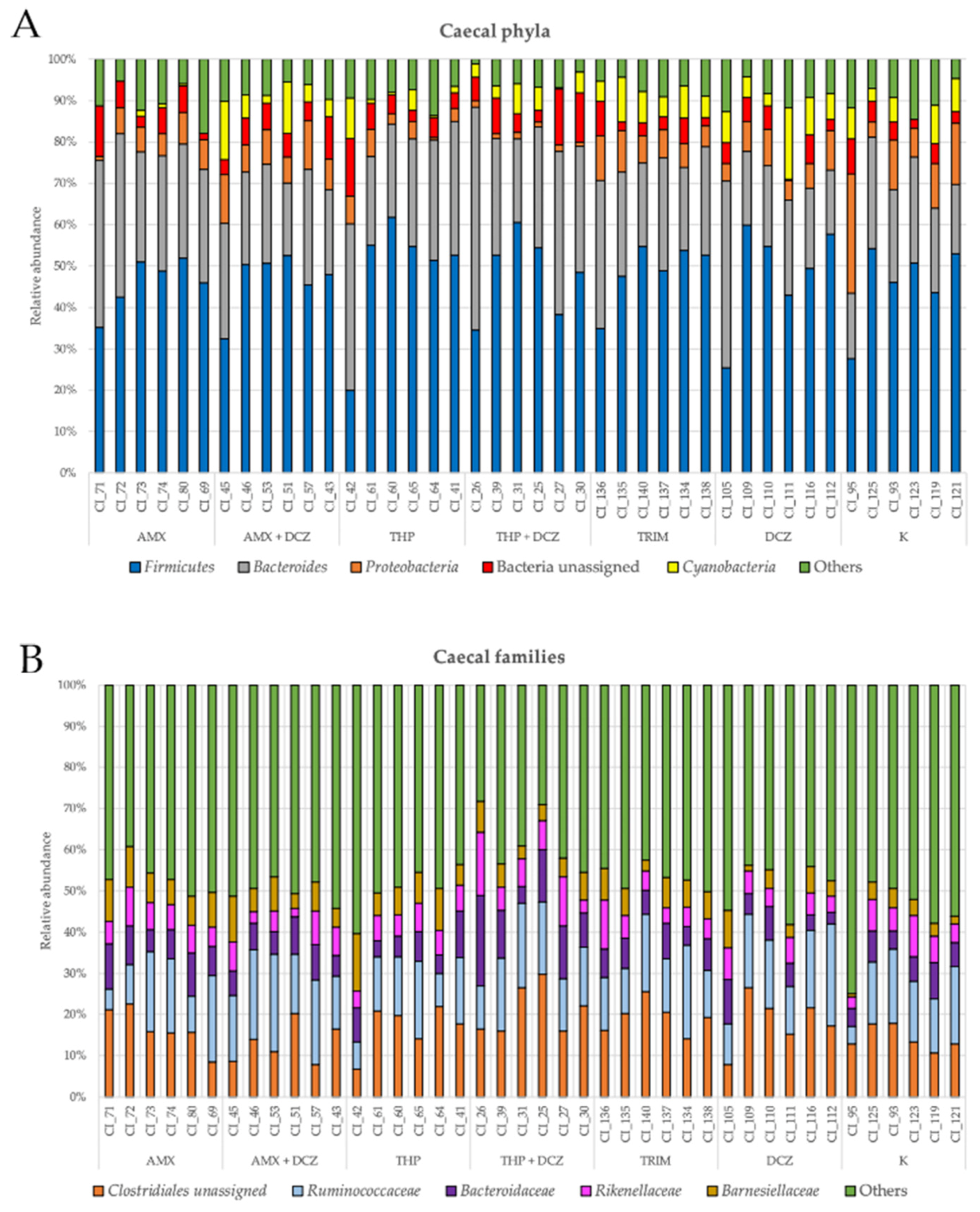
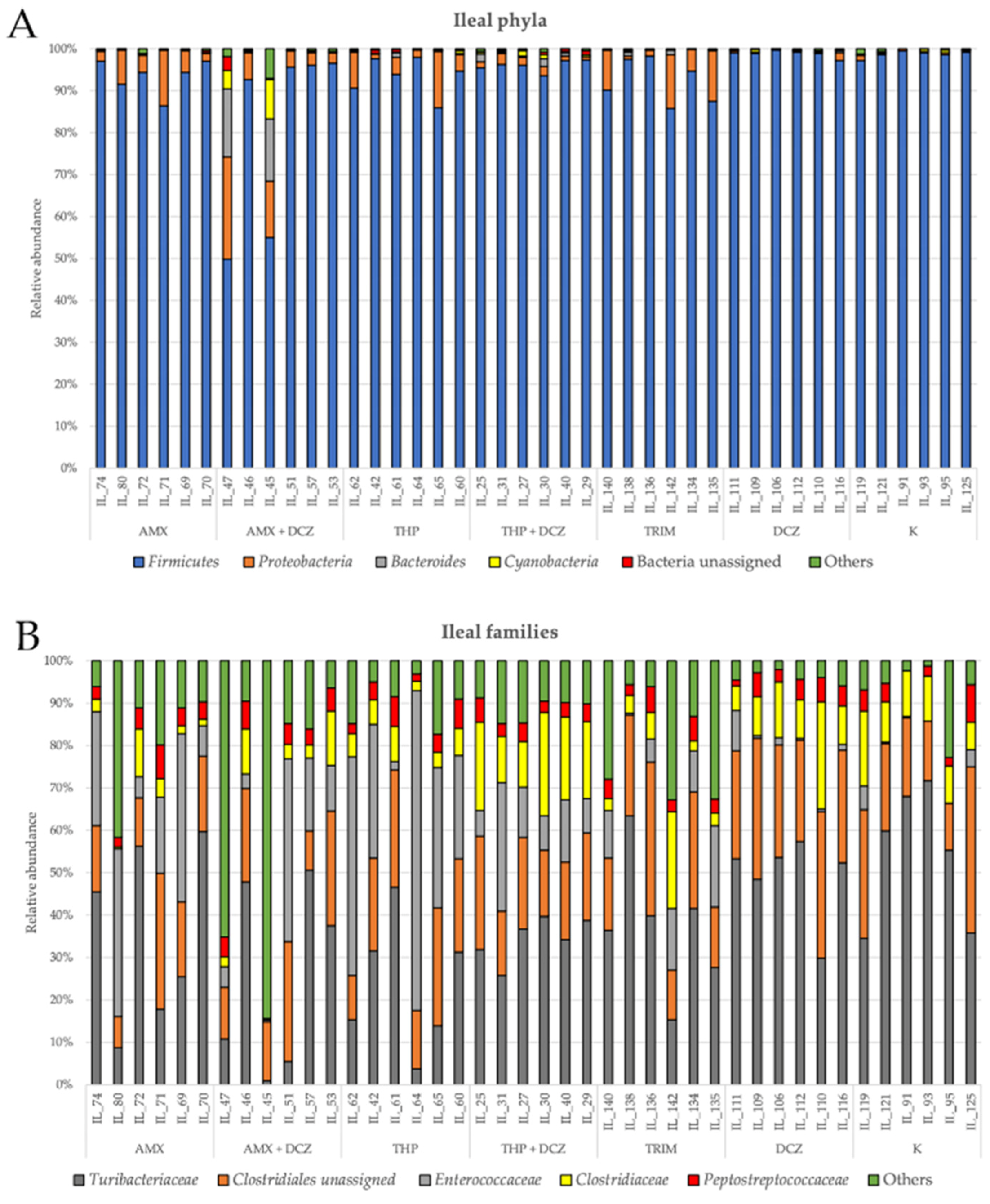
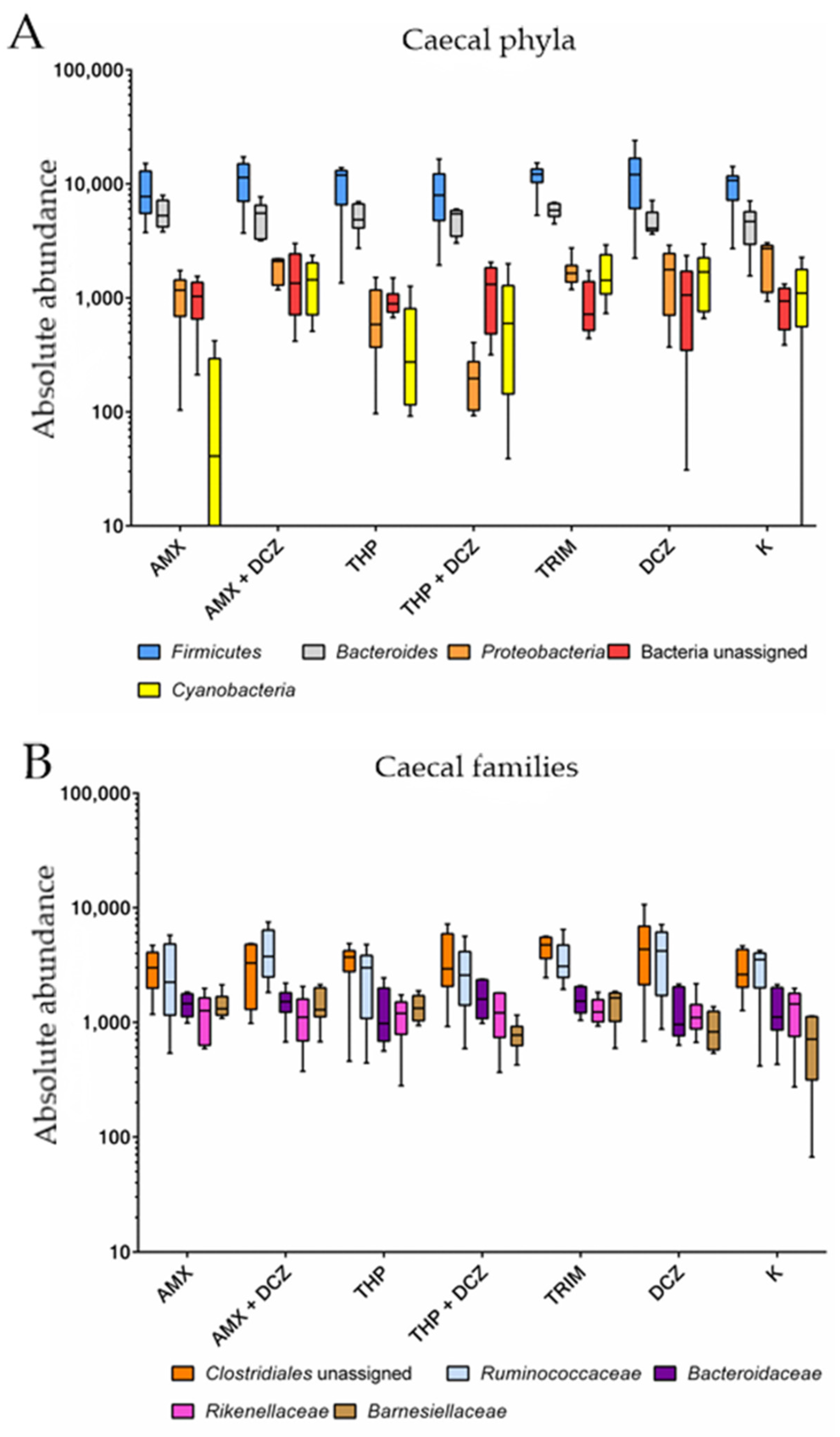
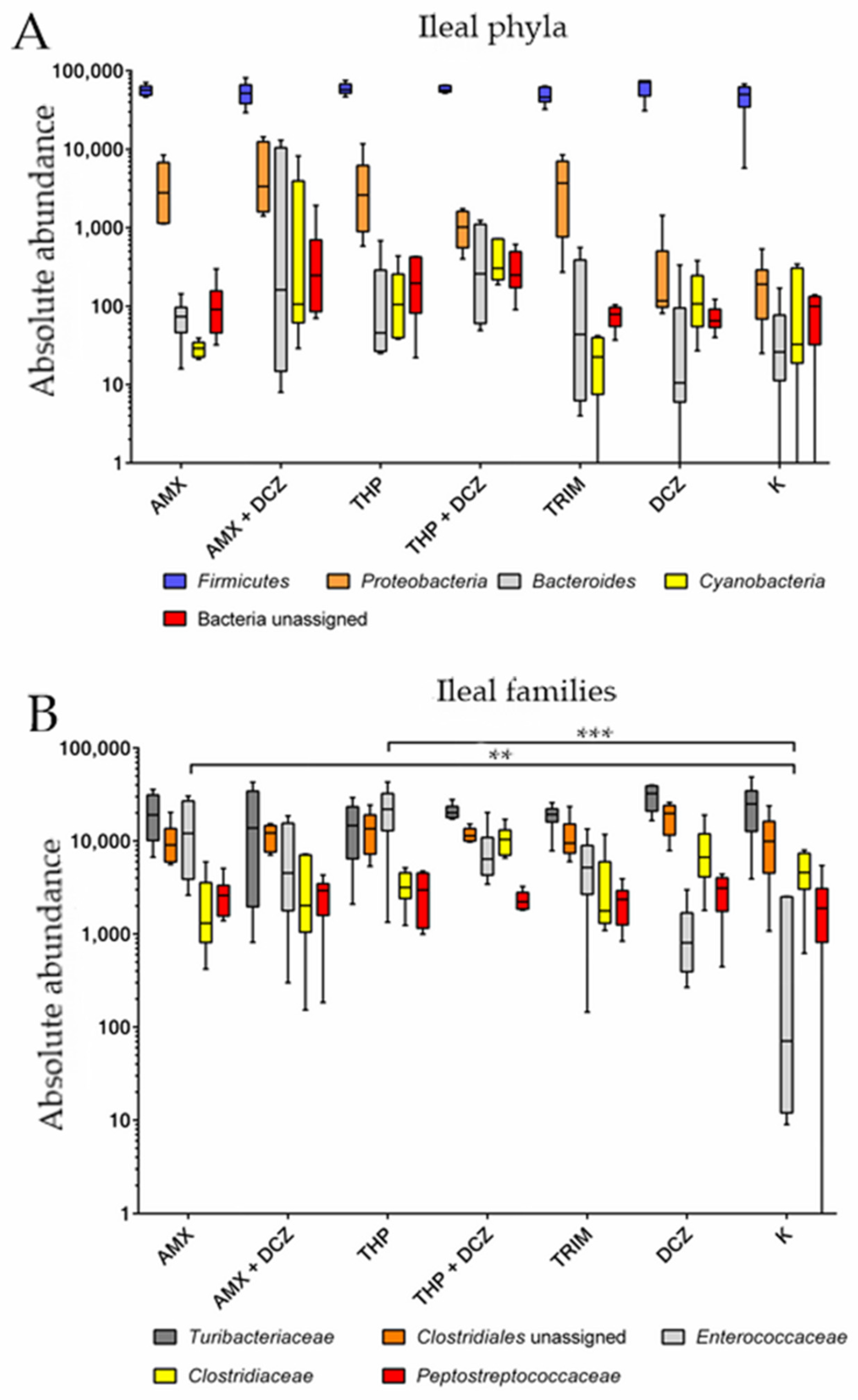
2.2. 16S rRNA Taxonomy Analysis
3. Discussion
4. Materials and Methods
4.1. Animals and Samples Collection
4.2. DNA Extraction and 16S rRNA Sequencing
4.3. Bioinformatic Analyses
4.4. Taxonomy Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Roy, C.I.; Woodward, M.J.; Ellis, R.J.; La Ragione, R.M.; Claus, S.P. Antibiotic treatment triggers gut dysbiosis and modulates metabolism in a chicken model of gastro-intestinal infection. BMC Vet. Res. 2019, 15, 1–13. [Google Scholar] [CrossRef]
- Iacob, S.; Iacob, D.G. Infectious Threats, the Intestinal Barrier, and Its Trojan Horse: Dysbiosis. Front. Microbiol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and environmental factors affecting the intestinal microbiota in chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Yeoman, C.J.; Chia, N.; Jeraldo, P.; Sipos, M.; Goldenfeld, N.D.; White, B.A. The microbiome of the chicken gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Xiao, Y.; Xiang, Y.; Zhou, W.; Chen, J.; Li, K.; Yang, H. Microbial community mapping in intestinal tract of broiler chicken. Poult. Sci. 2017, 96, 1387–1393. [Google Scholar] [CrossRef]
- Fletcher, O.J.; Mansell, R.; Martin, M.P.; Borst, L.B.; Barnes, H.J.; Gonzalez, L.M. Gross morphometry, histomorphometry, and immunohistochemistry confirm early and persistent jejunal crypt hyperplasia in poults with enteritis and depressed growth. Avian Dis. 2018, 62, 163–170. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.; Wang, Y.; Sun, Y.; Ma, L.; Zeng, Q.; Jiang, X.; Li, A.; Zeng, Z.; Zhang, T. Antibiotic-mediated changes in the fecal microbiome of broiler chickens define the incidence of antibiotic resistance genes. Microbiome 2018, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mehdi, Y.; Létourneau-Montminy, M.P.; Gaucher, M.L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Côté, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef]
- Morgun, A.; Dzutsev, A.; Dong, X.; Greer, R.L.; Sexton, D.J.; Ravel, J.; Schuster, M.; Hsiao, W.; Matzinger, P.; Shulzhenko, N. Uncovering effects of antibiotics on the host and microbiota using transkingdom gene networks. Gut 2015, 64, 1732–1743. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Lee, K.; Kim, D.W.; Kil, D.Y.; Kim, G.B.; Cha, C.J. Influence of dietary avilamycin on ileal and cecal microbiota in broiler chickens. Poult. Sci. 2018, 97, 970–979. [Google Scholar] [CrossRef]
- Gadde, U.D.; Oh, S.; Lillehoj, H.S.; Lillehoj, E.P. Antibiotic growth promoters virginiamycin and bacitracin methylene disalicylate alter the chicken intestinal metabolome. Sci. Rep. 2018, 8, 3592. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.C.; Bessegatto, J.A.; Alfieri, A.A.; Weese, J.S.; Filho, J.A.B.; Oba, A. Different antibiotic growth promoters induce specific changes in the cecal microbiota membership of broiler chicken. PLoS ONE 2017, 12, e171642. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Liu, F.; Cao, J.; Lv, N.; Zhu, B.; Zhang, G.; Fu Gao, G. Integrated metagenomic and metatranscriptomic profiling reveals differentially expressed resistomes in human, chicken, and pig gut microbiomes. Environ. Int. 2020, 138, 105649. [Google Scholar] [CrossRef]
- Guardabassi, L.; Apley, M.; Olsen, J.E.; Toutain, P.-L.; Weese, S. Optimization of Antimicrobial Treatment to Minimize Resistance Selection. Microbiol. Spectr. 2018, 6, 637–673. [Google Scholar] [CrossRef]
- Kers, J.G.; Fischer, E.A.J.; Stegeman, J.A.; Smidt, H.; Velkers, F.C. Comparison of different invasive and non-invasive methods to characterize intestinal microbiota throughout a production cycle of broiler chickens. Microorganisms 2019, 7, 431. [Google Scholar] [CrossRef] [Green Version]
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Lemos, M.P.L.; Saraiva, M.M.S.; Leite, E.L.; Silva, N.M.V.; Vasconcelos, P.C.; Giachetto, P.F.; Freitas Neto, O.C.; Givisiez, P.E.N.; Gebreyes, W.A.; Oliveira, C.J.B. The posthatch prophylactic use of ceftiofur affects the cecal microbiota similar to the dietary sanguinarine supplementation in broilers. Poult. Sci. 2020, 99, 6013–6021. [Google Scholar] [CrossRef]
- Wagner, B.D.; Grunwald, G.K.; Zerbe, G.O.; Mikulich-Gilbertson, S.K.; Robertson, C.E.; Zemanick, E.T.; Harris, J.K. On the use of diversity measures in longitudinal sequencing studies of microbial communities. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Ngunjiri, J.; Taylor, K.; Abundo, M.; Jang, H.; Elaish, M.; Mahesh, K.; Ghorbani, A.; Wijeratne, S.; Weber, B.; Johnson, T.; et al. Farm Stage, Bird Age, and Body Site Dominantly Affect the Quantity, Taxonomic Composition, and Dynamics of Respiratory and Gut Microbiota of Commercial Layer Chickens. Appl. Environ. Microbiol. 2019, 85, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Glendinning, L.; Watson, K.A.; Watson, M. Development of the duodenal, ileal, jejunal and caecal microbiota in chickens. Anim. Microbiome 2019, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mohd Shaufi, M.A.; Sieo, C.C.; Chong, C.W.; Gan, H.M.; Ho, Y.W. Deciphering chicken gut microbial dynamics based on high-throughput 16S rRNA metagenomics analyses. Gut Pathog. 2015, 7, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Kalkut, G. Sulfonamides and trimethoprim. Cancer Investig. 1998, 16, 612–615. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Beta-Lactams and beta-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Saliu, E.M.; Vahjen, W.; Zentek, J. Types and prevalence of extended-spectrum beta-lactamase producing Enterobacteriaceae in poultry. Anim. Health Res. Rev. 2017, 18, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Apostolakos, I.; Mughini-Gras, L.; Fasolato, L.; Piccirillo, A. Assessing the occurrence and transfer dynamics of ESBL/pAmpC-producing Escherichia coli across the broiler production pyramid. PLoS ONE 2019, 14, e217174. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 6, 185–227. [Google Scholar] [CrossRef]
- Gagetti, P.; Bonofiglio, L.; García Gabarrot, G.; Kaufman, S.; Mollerach, M.; Vigliarolo, L.; von Specht, M.; Toresani, I.; Lopardo, H.A. Resistance to β-lactams in enterococci. Rev. Argent. Microbiol. 2019, 51, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Kohler, V.; Vaishampayan, A.; Grohmann, E. Broad-host-range Inc18 plasmids: Occurrence, spread and transfer mechanisms. Plasmid 2018, 99, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Giannuzzi, D.; Biolatti, B.; Longato, E.; Divari, S.; Starvaggi Cucuzza, L.; Pregel, P.; Scaglione, F.E.; Rinaldi, A.; Chiesa, L.M.; Cannizzo, F.T. Application of RNA-sequencing to identify biomarkers in broiler chickens prophylactic administered with antimicrobial agents. Animal 2020, in press. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Elokil, A.A.; Abouelezz, K.F.M.; Ahmad, H.I.; Pan, Y.; Li, S. Investigation of the Impacts of Antibiotic Exposure on the Diversity of the Gut Microbiota in Chicks. Animals 2020, 10, 896. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Values | ||||
|---|---|---|---|---|
| Source of Variation | Phyla | Families | ||
| Caecum | Ileum | Caecum | Ileum | |
| Interaction | 0.92 | 0.31 | 0.32 | <0.0001 |
| Groups | 0.64 | 0.32 | 0.81 | 0.17 |
| Taxa | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Group | AMs | Dosing Protocols per os | Time-Lapse of Treatments (Rearing Days) | Withdrawal (Days) |
|---|---|---|---|---|
| AMX | Amoxicillin | 30 mg·kg−1 b.w. twice/day | 20–22/53–56 | 1 |
| AMX + DCZ | Amoxicillin Diclazuril | 30 mg·kg−1 b.w. twice/day 1 mg/kg | 20–22/53–56 0–52 | 1 5 |
| THP | Thiamphenicol | 65 mg·kg−1 b.w./day | 23–25/47–51 | 6 |
| THP + DCZ | Thiamphenicol Diclazuril | 65 mg·kg−1 b.w./day 1 mg/kg | 23–25/47–51 0–52 | 6 5 |
| TRIM | Sulfadiazine Trimethoprim | 20 mg·kg−1 b.w./day 4 mg b.w./day | 21–25/50–54 | 3 |
| DCZ | Diclazuril | 1 mg/kg | 0–52 | 5 |
| K | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuccato, M.; Rubiola, S.; Giannuzzi, D.; Grego, E.; Pregel, P.; Divari, S.; Cannizzo, F.T. 16S rRNA Sequencing Analysis of the Gut Microbiota in Broiler Chickens Prophylactically Administered with Antimicrobial Agents. Antibiotics 2021, 10, 146. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020146
Cuccato M, Rubiola S, Giannuzzi D, Grego E, Pregel P, Divari S, Cannizzo FT. 16S rRNA Sequencing Analysis of the Gut Microbiota in Broiler Chickens Prophylactically Administered with Antimicrobial Agents. Antibiotics. 2021; 10(2):146. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020146
Chicago/Turabian StyleCuccato, Matteo, Selene Rubiola, Diana Giannuzzi, Elena Grego, Paola Pregel, Sara Divari, and Francesca Tiziana Cannizzo. 2021. "16S rRNA Sequencing Analysis of the Gut Microbiota in Broiler Chickens Prophylactically Administered with Antimicrobial Agents" Antibiotics 10, no. 2: 146. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10020146