
Inhibition of LpxC Increases the Activity of Iron Chelators and Gallium Nitrate in Multidrug-Resistant Acinetobacter baumannii


Abstract
:1. Introduction
2. Results
2.1. Checkerboard Assays with A. baumannii Reference Strains
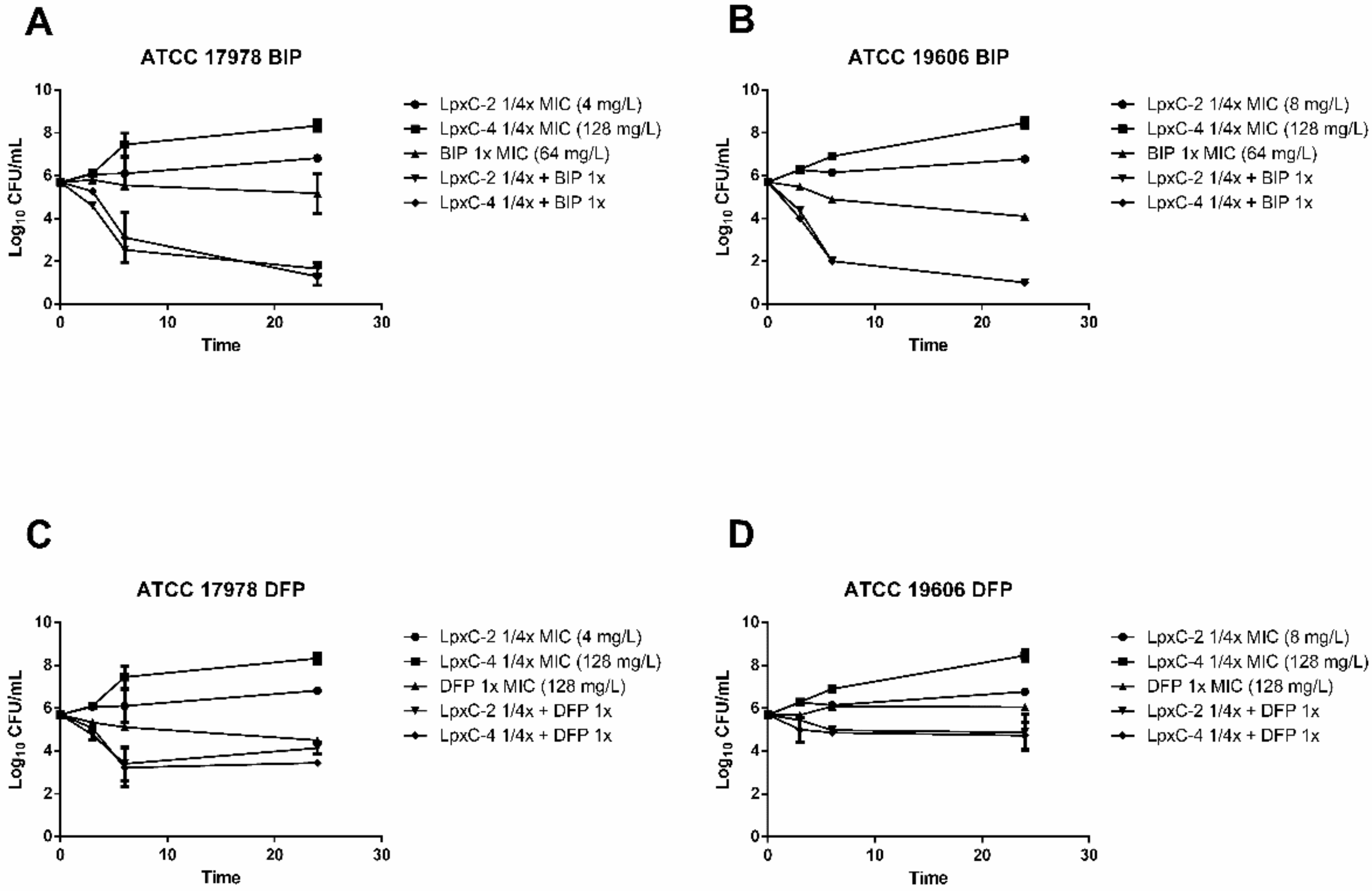
2.2. Time–Kill Analysis of Combinations of LpxC Inhibitors and Iron Chelators with A. baumannii Reference Strains
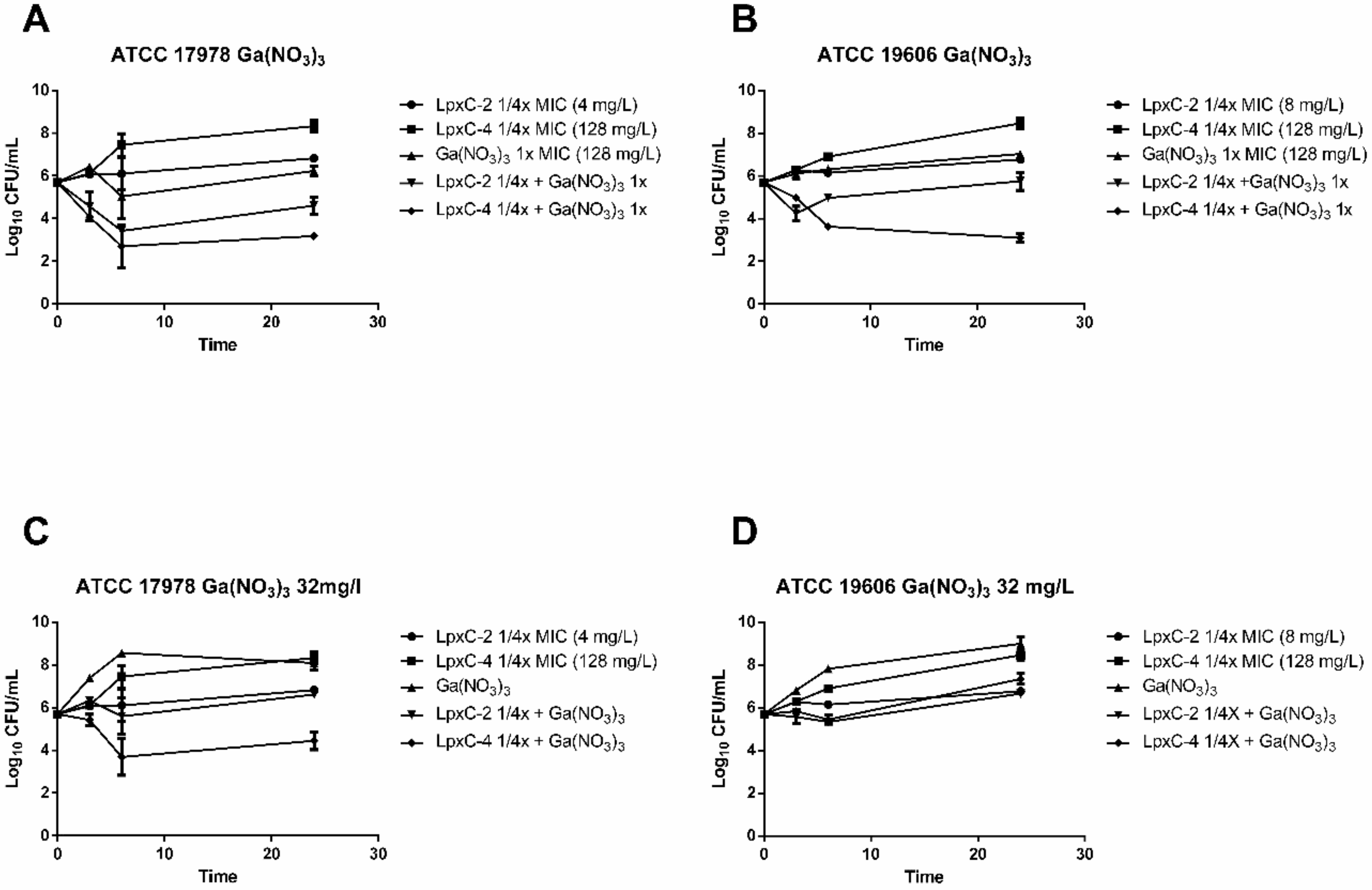
2.3. Time–Kill Analysis of Combinations of LpxC Inhibitors and Ga(NO3)3 with A. baumannii Reference Strains
2.4. Activity of LpxC Inhibitors in Combination with Iron Chelators and Gallium Nitrate in A. baumannii Clinical Isolates
3. Discussion
4. Materials and Methods
4.1. A. baumannii Strains Used in This Study
4.2. LpxC Inhibitors, Iron Chelators and Gallium Nitrate
4.3. Checkerboard Assays
4.4. Time–Kill Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McConnell, M.J.; Actis, L.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Ma, S. Recent Process in the Inhibitors of UDP-3-O-(R-3-hydroxyacyl)-Nacetylglucosamine Deacetylase (LpxC) Against Gram-Negative Bacteria. Mini Rev. Med. Chem. 2018, 18, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Baquir, B.; Tomaras, A.; Montgomery, J.I.; Reilly, U.; Barbacci, E.G.; Hujer, K.; et al. Inhibition of LpxC protects mice from resistant Acinetobacter baumannii by modulating inflammation and enhancing phagocytosis. mBio 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, J.I.; Brown, M.F.; Reilly, U.; Price, L.M.; Abramite, J.A.; Arcari, J.; Barham, R.; Che, Y.; Chen, J.M.; Chung, S.W.; et al. Pyridone methylsulfone hydroxamate LpxC inhibitors for the treatment of serious gram-negative infections. J. Med. Chem. 2012, 55, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, A.P.; McPherson, C.J.; Kuhn, M.; Carifa, A.; Mullins, L.; George, D.; Desbonnet, C.; Eidem, T.M.; Montgomery, J.I.; Brown, M.F.; et al. LpxC inhibitors as new antibacterial agents and tools for studying regulation of lipid A biosynthesis in Gram-negative pathogens. mBio 2014, 5, e01551-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St Michael, F.; Cox, A.D.; et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffatt, J.H.; Harper, M.; Adler, B.; Nation, R.L.; Li, J.; Boyce, J.D. Insertion sequence ISAba11 is involved in colistin resistance and loss of lipopolysaccharide in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3022–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carretero-Ledesma, M.; García-Quintanilla, M.; Martín-Peña, R.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Phenotypic changes associated with Colistin resistance due to Lipopolysaccharide loss in Acinetobacter baumannii. Virulence 2018, 9, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the essentiality of lipopolysaccharide to Gram-negative bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuesa, V.; McConnell, M.J. Recent Advances in Iron Chelation and Gallium-Based Therapies for Antibiotic Resistant Bacterial Infections. Int. J. Mol. Sci. 2021, 22, 2876. [Google Scholar] [CrossRef]
- Antunes, L.C.; Imperi, F.; Minandri, F.; Visca, P. In vitro and in vivo antimicrobial activities of gallium nitrate against multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 5961–5970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.R.; Britigan, B.E.; Narayanasamy, P. Dual Inhibition of Klebsiella pneumoniae and Pseudomonas aeruginosa Iron Metabolism Using Gallium Porphyrin and Gallium Nitrate. ACS Infect. Dis. 2019, 5, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Hijazi, S.; Visaggio, D.; Pirolo, M.; Frangipani, E.; Bernstein, L.; Visca, P. Antimicrobial Activity of Gallium Compounds on ESKAPE Pathogens. Front. Cell Infect. Microbiol. 2018, 8, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goss, C.H.; Kaneko, Y.; Khuu, L.; Anderson, G.D.; Ravishankar, S.; Aitken, M.L.; Lechtzin, N.; Zhou, G.; Czyz, D.M.; McLean, K.; et al. Gallium disrupts bacterial iron metabolism and has therapeutic effects in mice and humans with lung infections. Sci. Transl. Med. 2018, 10, eaat7520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, P.J.; Labthavikul, P.; Jones, C.H.; Bradford, P.A. In vitro antibacterial activities of tigecycline in combination with other antimicrobial agents determined by chequerboard and time-kill kinetic analysis. J. Antimicrob. Chemother. 2006, 57, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, L.R. Mechanisms of therapeutic activity for gallium. Pharmacol. Rev. 1998, 50, 665–682. [Google Scholar] [PubMed]
- Fernández-Cuenca, F.; Pascual, A.; Ribera, A.; Vila, J.; Bou, G.; Cisneros, J.M.; Rodríguez-Baño, J.; Pachón, J.; Martínez-Martínez, L.; Hospitalaria, G.d.E.d.I. Clonal diversity and antimicrobial susceptibility of Acinetobacter baumannii isolated in Spain. A nationwide multicenter study: GEIH-Ab project (2000). Enferm. Infecc. Microbiol. Clin. 2004, 22, 267–271. [Google Scholar] [CrossRef]
- Fernández-Cuenca, F.; Tomás-Carmona, M.; Caballero-Moyano, F.; Bou, G.; Martínez-Martínez, L.; Vila, J.; Pachón, J.; Cisneros, J.M.; Rodríguez-Baño, J.; Pascual, A.; et al. In vitro activity of 18 antimicrobial agents against clinical isolates of Acinetobacter spp.: Multicenter national study GEIH-REIPI-Ab 2010. Enferm. Infecc. Microbiol. Clin. 2013, 31, 4–9. [Google Scholar] [CrossRef] [PubMed]
- García-Quintanilla, M.; Caro-Vega, J.M.; Pulido, M.R.; Moreno-Martínez, P.; Pachón, J.; McConnell, M.J. Inhibition of LpxC Increases Antibiotic Susceptibility in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 5076–5079. [Google Scholar] [CrossRef] [Green Version]
- Minandri, F.; Bonchi, C.; Frangipani, E.; Imperi, F.; Visca, P. Promises and failures of gallium as an antibacterial agent. Future Microbiol. 2014, 9, 379–397. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Strain | MIC (mg/L) | ||||
|---|---|---|---|---|---|
| BIP | DFP | Ga(NO3)3 | LpxC-2 | LpxC-4 | |
| ATCC 17978 | 64 | 128 | 128 | 16 | 512 |
| ATCC 19606 | 64 | 128 | 128 | 32 | 512 |
| Ab-1 | 64 | ND | 128 | 16 | >512 |
| Ab-66 | 64 | ND | 128 | 32 | >512 |
| Ab-84 | 64 | ND | 128 | 16 | >512 |
| Ab-108 | 64 | ND | 32 | 16 | >512 |
| Ab-154 | 32 | ND | 64 | 16 | 512 |
| Ab-167 | 64 | ND | 128 | 128 | >512 |
| Ab-176 | 64 | ND | 64 | 16 | >512 |
| Ab-208 | 64 | ND | 128 | 8 | >512 |
| Strain | BIP | DFP | Ga(NO3)3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LpxC-2 | LpxC-4 | LpxC-2 | LpxC-4 | LpxC-2 | LpxC-4 | |||||||
| FICI Value | Effect of Combination | FICI Value | Effect of Combination | FICI Value | Effect of Combination | FICI Value | Effect of Combination | FICI Value | Effect of Combination | FICI Value | Effect of Combination | |
| ATCC 17978 | 0.53 | Indifference | 0.09 | Synergy | 1.01 | Indifference | 1.00 | Indifference | 0.38 | Synergy | 0.38 | Synergy |
| ATCC 19606 | 0.50 | Synergy | 0.53 | Indifference | 1.00 | Indifference | 1.00 | Indifference | 0.27 | Synergy | 0.08 | Synergy |
| Ab-1 | 1.02 | Indifference | ≤0.50 | Synergy | ND | - | ND | - | 1.00 | Indifference | ≤0.19 | Synergy |
| Ab-66 | 0.38 | Synergy | ≤0.50 | Synergy | ND | - | ND | - | 0.38 | Synergy | ≤0.19 | Synergy |
| Ab-84 | 1.00 | Indifference | ≤0.63 | Indifference | ND | - | ND | - | 0.50 | Synergy | ≤0.09 | Synergy |
| Ab-108 | 0.63 | Indifference | ≤0.50 | Synergy | ND | - | ND | - | 0.75 | Indifference | ≤0.16 | Synergy |
| Ab-154 | 0.56 | Indifference | 0.75 | Indifference | ND | - | ND | - | 0.53 | Indifference | 0.31 | Synergy |
| Ab-167 | 0.50 | Synergy | ≤0.75 | Indifference | ND | - | ND | - | 0.53 | Indifference | ≤0.50 | Synergy |
| Ab-176 | 0.52 | Indifference | ≤0.38 | Synergy | ND | - | ND | - | 0.50 | Synergy | ≤0.19 | Synergy |
| Ab-208 | 0.53 | Indifference | ≤0.38 | Synergy | ND | - | ND | - | 0.75 | Indifference | ≤0.09 | Synergy |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinuesa, V.; Cruces, R.; Nonnoi, F.; McConnell, M.J. Inhibition of LpxC Increases the Activity of Iron Chelators and Gallium Nitrate in Multidrug-Resistant Acinetobacter baumannii. Antibiotics 2021, 10, 609. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050609
Vinuesa V, Cruces R, Nonnoi F, McConnell MJ. Inhibition of LpxC Increases the Activity of Iron Chelators and Gallium Nitrate in Multidrug-Resistant Acinetobacter baumannii. Antibiotics. 2021; 10(5):609. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050609
Chicago/Turabian StyleVinuesa, Víctor, Raquel Cruces, Francesca Nonnoi, and Michael J. McConnell. 2021. "Inhibition of LpxC Increases the Activity of Iron Chelators and Gallium Nitrate in Multidrug-Resistant Acinetobacter baumannii" Antibiotics 10, no. 5: 609. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050609