
Evaluation of a Biocide Used in the Biological Isolation and Containment Unit of a Veterinary Teaching Hospital

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility Testing
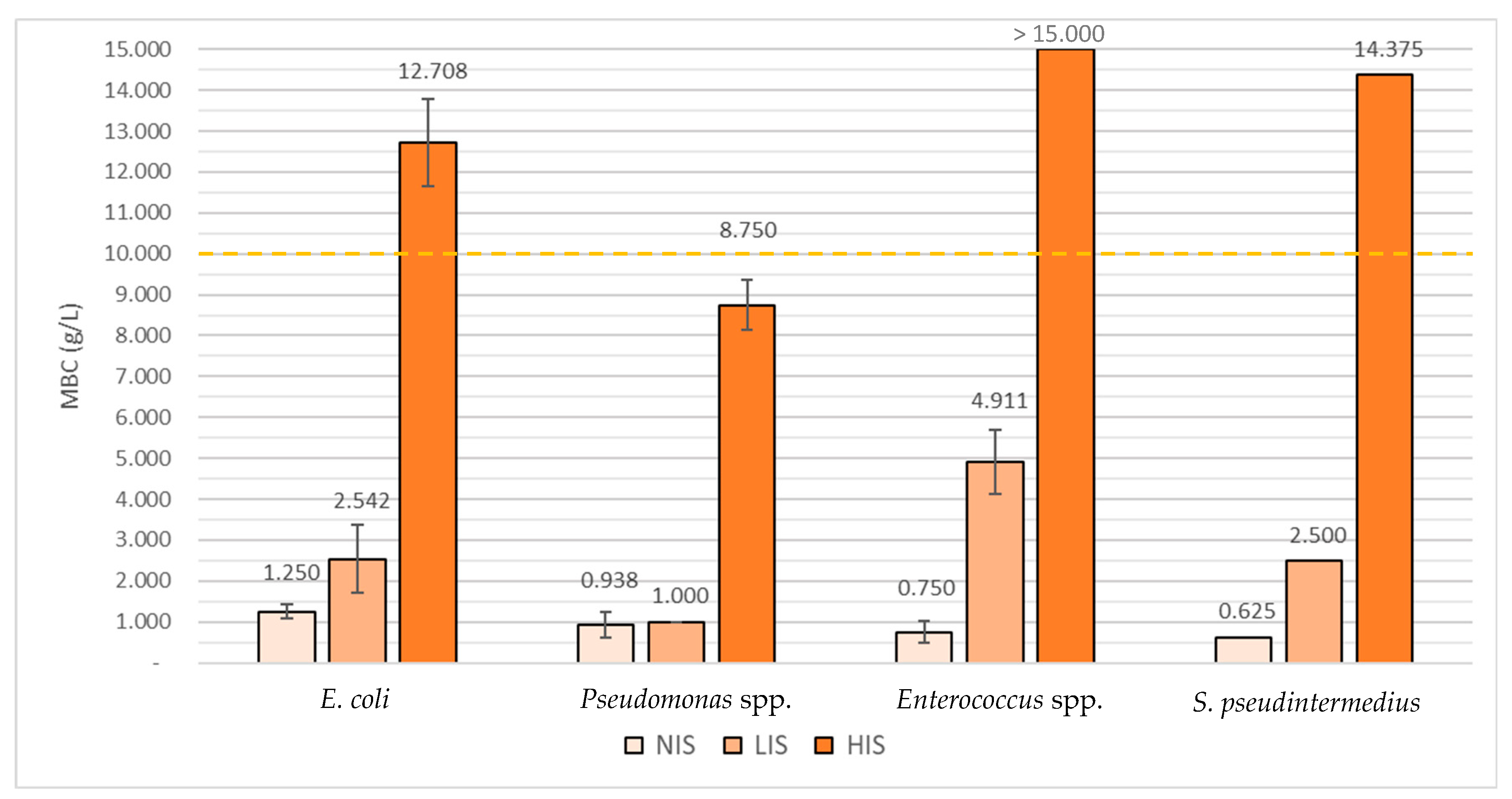
2.2. Determination of Virkon™ S’s Minimal Inhibitory Concentrations (MICs) and Minimal Bactericidal Concentrations (MBCs)
2.3. Influence of Bacterial Exposure to Sub-Lethal Concentrations of Virkon™ S
3. Discussion
3.1. Antimicrobial Susceptibility Testing
3.2. Determination of Virkon™ S’s Minimal Inhibitory Concentrations (MICs) and Minimal Bactericidal Concentrations (MBCs)
3.3. Influence of Bacterial Exposure to Sub-Lethal Concentrations of Virkon™ S
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. Determination of Minimal Inhibitory Concentrations (MICs) and Minimal Bactericidal Concentrations (MBCs)
4.4. Influence of Virkon™ S Sub-MBC Values on Isolates Biocide Resistance Ability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guardabassi, L. Veterinary hospital-acquired infections: The challenge of MRSA and other multidrug-resistant bacterial infections in veterinary medicine. Vet. J. 2012, 193, 307–308. [Google Scholar] [CrossRef] [PubMed]
- Stull, J.W.; Weese, J.S. Hospital-Associated Infections in Small Animal Practice. Vet. Clin. N. Am. Small Anim. Pract. 2015, 45, 217–233. [Google Scholar] [CrossRef]
- Weese, J.S. Infection control in veterinary practice; the time is now. J. Small Anim. Pract. 2011, 52, 507–508. [Google Scholar] [CrossRef]
- Benedict, K.M.; Morley, P.S.; Van Metre, D.C. Characteristics of biosecurity and infection control programs at veterinary teaching hospitals. J. Am. Vet. Med. Assoc. 2008, 233, 767–773. [Google Scholar] [CrossRef] [Green Version]
- Burgess, B.A.; Morley, P.S.; Hyatt, D.R. Environmental surveillance for Salmonella enterica in a veterinary teaching hospital. J. Am. Vet. Med. Assoc. 2004, 225, 1344–1348. [Google Scholar] [CrossRef]
- Hamilton, E.; Kaneene, J.B.; May, J.K.; Kruger, J.M.; Schall, W.; Beal, M.W.; Hauptman, J.G.; De Camp, C.E. Prevalence and antimicrobial resistance of Enterococcus spp and Staphylococcus spp isolated from surfaces in a veterinary teaching hospital. J. Am. Vet. Med. Assoc. 2012, 240, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Weese, J.S.; Dacosta, T.; Button, L.; Goth, K.; Ethier, M.; Boehnke, K. Isolation of Methicillin-Resistant Staphylococcus aureus from the Environment in a Veterinary Teaching Hospital. J. Vet. Intern. Med. 2004, 18, 468–470. [Google Scholar] [CrossRef]
- Weese, J.; Dick, H.; Willey, B.; McGeer, A.; Kreiswirth, B.; Innis, B.; Low, D. Suspected transmission of methicillin-resistant Staphylococcus aureus between domestic pets and humans in veterinary clinics and in the household. Vet. Microbiol. 2006, 115, 148–155. [Google Scholar] [CrossRef]
- Schreiber, P.W.; Sax, H.; Wolfensberger, A.; Clack, L.; Kuster, S.P.; Swissnoso. The preventable proportion of healthcare-associated infections 2005–2016: Systematic review and meta-analysis. Infect. Control Hosp. Epidemiol. 2018, 39, 1277–1295. [Google Scholar] [CrossRef] [Green Version]
- Canadian Committee on Antibiotic Resistance (CCAR). Infection Prevention and Control Best Practices: For Small Animal Veterinary Clinics. Available online: https://www.wormsandgermsblog.com/files/2008/04/CCAR-Guidelines-Final2.pdf (accessed on 23 October 2019).
- Mount, R.; Schick, A.E.; Lewis, T.P.; Newton, H.M. Evaluation of Bacterial Contamination of Clipper Blades in Small Animal Private Practice. J. Am. Anim. Hosp. Assoc. 2016, 52, 95–101. [Google Scholar] [CrossRef]
- Stull, J.W.; Bjorvik, E.; Bub, J.; Dvorak, G.; Petersen, C.; Troyer, H.L. 2018 AAHA Infection Control, Prevention, and Biosecurity Guidelines. J. Am. Anim. Hosp. Assoc. 2018, 54, 297–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regulation (UE), No. 528/2012 of 22 of May 2012; Official Journal of the European Union—L167/1; European Parliament and Council: Brussels, Belgium, 2012.
- Hernández-Navarrete, M.J.; Celorrio-Pascual, J.M.; Moros, C.L.; Bernad, V.M.S. Principles of antisepsis, disinfection and sterilization. Enferm. Infect. Microbiol. Clin. 2014, 32, 681–688. [Google Scholar] [CrossRef]
- Sheldon, A.T. Antiseptic “Resistance”: Real or Perceived Threat? Clin. Infect. Dis. 2005, 40, 1650–1656. [Google Scholar] [CrossRef] [PubMed]
- Maillard, J.-Y. Factors Affecting the Activities of Microbicides. In Russell, Hugo & Ayliffe’s Principles and Practice of Disinfection, Preservation & Sterilization; Fraise, A.P., Lambert, P.A., Maillard, J.-Y., Eds.; Blackwell Publishing: London, UK, 2013; pp. 71–86. [Google Scholar]
- Maillard, J.-Y. Antimicrobial biocides in the healthcare environment: Efficacy, usage, policies, and perceived problems. Ther. Clin. Risk Manag. 2005, 1, 307–320. [Google Scholar]
- Buffet-Bataillon, S.; Tattevin, P.; Maillard, J.-Y.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Efflux pump induction by quaternary ammonium compounds and fluoroquinolone resistance in bacteria. Future Microbiol. 2016, 11, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Grande Burgos, M.J.; Fernández Márquez, M.L.; Pérez Pulido, R.; Gálvez, A.; Lucas López, R. Virulence factors and antimicrobial resistance in Escherichia coli strains isolated from hen eggshells. Int. J. Food Microbiol. 2016, 238, 89–95. [Google Scholar] [CrossRef]
- Kim, M.; Hatt, J.K.; Weigand, M.R.; Krishnan, R.; Pavlostathis, S.G.; Konstantinidis, K.T. Genomic and Transcriptomic Insights into How Bacteria Withstand High Concentrations of Benzalkonium Chloride Biocides. Appl. Environ. Microbiol. 2018, 84, e00197-18. [Google Scholar] [CrossRef] [Green Version]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes Strains Selected on Ciprofloxacin or the Disinfectant Benzalkonium Chloride Exhibit Reduced Susceptibility to Ciprofloxacin, Gentamicin, Benzalkonium Chloride, and Other Toxic Compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [Green Version]
- Gadea, R.; Glibota, N.; Pérez Pulido, R.; Gálvez, A.; Ortega, E. Adaptation to Biocides Cetrimide and Chlorhexidine in Bacteria from Organic Foods: Association with Tolerance to Other Antimicrobials and Physical Stresses. J. Agric. Food Chem. 2017, 65, 1758–1770. [Google Scholar] [CrossRef]
- Tattawasart, U.; Maillard, J.-Y.; Furr, J.R.; Russell, A.D. Outer membrane changes in Pseudomonas stutzeri resistant to chlorhexidine diacetate and cetylpyridinium chloride. Int. J. Antimicrob. Agents 2000, 16, 233–238. [Google Scholar] [CrossRef]
- Lanxess. Virkon™ S Disinfectant and Virucide. Available online: https://syndel.com/wp-content/uploads/2019/01/Virkon-S-Information-Page-Lanxess.pdf (accessed on 8 July 2020).
- Gabbert, L.R.; Smith, J.D.; Neilan, J.G.; Ferman, G.S.; Rasmussen, M.V. Smart Card Decontamination in a High-Containment Laboratory. Health Secur. 2018, 16, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Saklou, N.T.; Burgess, B.A.; Van Metre, D.C.; Hornig, K.; Morley, P.S.; Byers, S.R. Comparison of disinfectant efficacy when using high-volume directed mist application of accelerated hydrogen peroxide and peroxymonosulfate disinfectants in a large animal hospital. Equine Vet. J. 2015, 48, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.M.; Rogers, A.T.; Jacob, M.E. Susceptibility of Escherichia coli O157:H7 to Disinfectants In Vitro and in Simulated Footbaths Amended with Manure. Foodborne Pathog. Dis. 2018, 15, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Sylte, M.J.; Chandra, L.C.; Looft, T. Evaluation of disinfectants and antiseptics to eliminate bacteria from the surface of turkey eggs and hatch gnotobiotic poults. Poult. Sci. 2017, 96, 2412–2420. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Selden, D.; Fooks, A.R.; Banyard, A. Inactivation of rabies virus. J. Virol. Methods 2017, 243, 109–112. [Google Scholar] [CrossRef]
- Dunowska, M.; Morley, P.S.; Hyatt, D.R. The effect of Virkon® S fogging on survival of Salmonella enterica and Staphylococcus aureus on surfaces in a veterinary teaching hospital. Vet. Microbiol. 2005, 105, 281–289. [Google Scholar] [CrossRef]
- McCormick, L.; Maheshwari, G. Inactivation of adenovirus types 5 and 6 by Virkon® S. Antivir. Res. 2004, 64, 27–33. [Google Scholar] [CrossRef]
- Lanxess. Safety Data Sheet. Available online: https://virkon.us/wp-content/uploads/sites/15/2017/11/VirkonTM-S-USA.pdf (accessed on 8 July 2020).
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; CLSI Document VET01-A4; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2013. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals; CLSI Supplement VET01-S2; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2013. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2016. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [Green Version]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection-Treatment and Antibiotic Resistance. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Eds.; Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 6, 185–227. [Google Scholar] [CrossRef]
- Iseppi, R.; Messi, P.; Anacarso, I.; Bondi, M.; Sabia, C.; Condò, C.; De Niederhausern, S. Antimicrobial resistance and virulence traits in Enterococcus strains isolated from dogs and cats. New Microbiol. 2015, 38, 369–378. [Google Scholar]
- Jackson, C.; Cray, P.; Davis, J.; Barrett, J.; Frye, J. Prevalence, species distribution and antimicrobial resistance of enterococci isolated from dogs and cats in the United States. J. Appl. Microbiol. 2009, 107, 1269–1278. [Google Scholar] [CrossRef]
- Awosile, B.B.; McClure, J.T.; Saab, M.E.; Heider, L.C. Antimicrobial resistance in bacteria isolated from cats and dogs from the Atlantic Provinces, Canada from 1994–2013. Can. Vet. J. 2018, 59, 885–893. [Google Scholar]
- Delgado, M.; Neto, I.; Correia, J.H.D.; Pomba, C. Antimicrobial resistance and evaluation of susceptibility testing among pathogenic enterococci isolated from dogs and cats. Int. J. Antimicrob. Agents 2007, 30, 98–100. [Google Scholar] [CrossRef]
- Ghosh, A.; Dowd, S.E.; Zurek, L. Dogs Leaving the ICU Carry a Very Large Multi-Drug Resistant Enterococcal Population with Capacity for Biofilm Formation and Horizontal Gene Transfer. PLoS ONE 2011, 6, e22451. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, Y.; Ito, C.; Kawashima, A.; Ishii, M.; Yamashiro, S.; Harada, K.; Ochi, H.; Sawada, T. Identification and Antimicrobial Susceptibility of Enterococci Isolated from Dogs and Cats Subjected to Differing Antibiotic Pressures. J. Vet. Med Sci. 2013, 75, 749–753. [Google Scholar] [CrossRef] [Green Version]
- Leite-Martins, L.; Mahú, M.I.; Costa, A.L.; Bessa, L.J.; Vaz-Pires, P.; Loureiro, L.; Ribeiro, J.N.; de Matos, A.J.F.; da Costa, P.M. Prevalence of antimicrobial resistance in faecal enterococci from vet-visiting pets and assessment of risk factors. Vet. Rec. 2015, 176, 674. [Google Scholar] [CrossRef] [PubMed]
- Ossiprandi, M.C.; Bottarelli, E.; Cattabiani, F.; Bianchi, E. Susceptibility to vancomycin and other antibiotics of 165 Enterococcus strains isolated from dogs in Italy. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 1–9. [Google Scholar] [CrossRef]
- Rodrigues, J.; Poeta, P.; Martins, A.; Costa, D. The Importance of Pets as Reservoirs of Resistant Enterococcus Strains, with Special Reference to Vancomycin. J. Vet. Med. Ser. B 2002, 49, 278–280. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Weese, J.S.; Blondeau, J.; Boothe, D.; Guardabassi, L.G.; Gumley, N.; Papich, M.; Jessen, L.R.; Lappin, M.; Rankin, S.; Westropp, J.L.; et al. International Society for Companion Animal Infectious Diseases (ISCAID) guidelines for the diagnosis and management of bacterial urinary tract infections in dogs and cats. Vet. J. 2019, 247, 8–25. [Google Scholar] [CrossRef]
- Brandl, K.; Plitas, G.; Mihu, C.N.; Ubeda, C.; Jia, T.; Fleisher, M.; Schnabl, B.; DeMatteo, R.P.; Pamer, E.G. Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits. Nat. Cell Biol. 2008, 455, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Donskey, C.J. The Role of the Intestinal Tract as a Reservoir and Source for Transmission of Nosocomial Pathogens. Clin. Infect. Dis. 2004, 39, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donskey, C.J.; Helfand, M.S.; Pultz, N.J.; Rice, L.B. Effect of Parenteral Fluoroquinolone Administration on Persistence of Vancomycin-Resistant Enterococcus faecium in the Mouse Gastrointestinal Tract. Antimicrob. Agents Chemother. 2004, 48, 326–328. [Google Scholar] [CrossRef] [Green Version]
- Donskey, C.J.; Hanrahan, J.A.; Hutton, R.A.; Rice, L.B. Effect of Parenteral Antibiotic Administration on the Establishment of Colonization with Vancomycin-Resistant Enterococcus faecium in the Mouse Gastrointestinal Tract. J. Infect. Dis. 2000, 181, 1830–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donskey, C.J.; Hanrahan, J.A.; Hutton, R.A.; Rice, L.B. Effect of Parenteral Antibiotic Administration on Persistence of Vancomycin-Resistant Enterococcus faecium in the Mouse Gastrointestinal Tract. J. Infect. Dis. 1999, 180, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-Term Antibiotic Treatment Has Differing Long-Term Impacts on the Human Throat and Gut Microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef] [Green Version]
- Ubeda, C.; Taur, Y.; Jenq, R.R.; Equinda, M.J.; Son, T.; Samstein, M.; Viale, A.; Socci, N.D.; Brink, M.R.V.D.; Kamboj, M.; et al. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J. Clin. Investig. 2010, 120, 4332–4341. [Google Scholar] [CrossRef]
- Machado, I.C.; Cunha, E.; Gomes, J.; Tavares, L.; Almeida, V.; Gil, S. Survival Rates of Dogs with Infectious Diseases in a Veterinary Hospital Isolation Unit; International Society for Companion Animal Infectious Diseases (ISCAID): Portland, OR, USA, 2018. [Google Scholar]
- Machado, I.C.; Cunha, E.; Gomes, J.; Tavares, L.; Almeida, V.; Gil, S. Survival Rates of Cats with Infectious Diseases in a Veterinary Hospital Isolation Unit; International Society for Companion Animal Infectious Diseases (ISCAID): Portland, OR, USA, 2018. [Google Scholar]
- Damborg, P.; Top, J.; Hendrickx, A.P.A.; Dawson, S.; Willems, R.J.L.; Guardabassi, L. Dogs Are a Reservoir of Ampicillin-Resistant Enterococcus faecium Lineages Associated with Human Infections. Appl. Environ. Microbiol. 2009, 75, 2360–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederhäusern, S.; Sabia, C.; Messi, P.; Guerrieri, E.; Manicardi, G.; Bondi, M. Glycopeptide-resistance transferability from vancomycin-resistant enterococci of human and animal source to Listeria spp. Lett. Appl. Microbiol. 2004, 39, 483–489. [Google Scholar] [CrossRef]
- Limbago, B.M.; Kallen, A.J.; Zhu, W.; Eggers, P.; McDougal, L.K.; Albrecht, V.S. Report of the 13th Vancomycin-Resistant Staphylococcus aureus Isolate from the United States. J. Clin. Microbiol. 2014, 52, 998–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigel, L.M.; Clewell, D.B.; Gill, S.R.; Clark, N.C.; McDougal, L.K.; Flannagan, S.E.; Kolonay, J.F.; Shetty, J.; Killgore, G.E.; Tenover, F.C. Genetic Analysis of a High-Level Vancomycin-Resistant Isolate of Staphylococcus aureus. Science 2003, 302, 1569–1571. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Clark, N.C.; McDougal, L.K.; Hageman, J.; McDonald, L.C.; Patel, J.B. Vancomycin-Resistant Staphylococcus aureus Isolates Associated with Inc18-Like vanA Plasmids in Michigan. Antimicrob. Agents Chemother. 2007, 52, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maillard, J.-Y. Resistance of Bacteria to Biocides. Microbiol. Spectr. 2018, 6, 109–126. [Google Scholar]
- Humayoun, S.B.; Hiott, L.M.; Gupta, S.K.; Barrett, J.B.; Woodley, T.A.; Johnston, J.J.; Jackson, C.R.; Frye, J.G. An assay for determining the susceptibility of Salmonella isolates to commercial and household biocides. PLoS ONE 2018, 13, e0209072. [Google Scholar] [CrossRef] [Green Version]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Kalchayanand, N.; Koohmaraie, M.; Wheeler, T.L. Effect of Exposure Time and Organic Matter on Efficacy of Antimicrobial Compounds against Shiga Toxin–Producing Escherichia coli and Salmonella. J. Food Prot. 2016, 79, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalli, A.; Marinaro, M.; Desario, C.; Corrente, M.; Camero, M.; Buonavoglia, C. In vitro virucidal activity of sodium hypochlorite against canine parvovirus type 2. Epidemiol. Infect. 2018, 146, 2010–2013. [Google Scholar] [CrossRef] [Green Version]
- Chandler-Bostock, R.; Mellits, K.H. Efficacy of disinfectants against porcine rotavirus in the presence and absence of organic matter. Lett. Appl. Microbiol. 2015, 61, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Alotaibi, S.M.I.; Ayibiekea, A.; Pedersen, A.F.; Jakobsen, L.; Pinholt, M.; Gumpert, H.; Hammerum, A.M.; Westh, H.; Ingmer, H. Susceptibility of vancomycin-resistant and -sensitive Enterococcus faecium obtained from Danish hospitals to benzalkonium chloride, chlorhexidine and hydrogen peroxide biocides. J. Med. Microbiol. 2017, 66, 1744–1751. [Google Scholar] [CrossRef]
- Márquez, M.L.F.; Burgos, M.J.G.; Pulido, R.P.; Gálvez, A.; López, R.L. Biocide Tolerance and Antibiotic Resistance in Salmonella Isolates from Hen Eggshells. Foodborne Pathog. Dis. 2017, 14, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Lerma, L.L.; Benomar, N.; Muñoz, M.D.C.C.; Galvez, A.; Abriouel, H. Correlation between antibiotic and biocide resistance in mesophilic and psychrotrophic Pseudomonas spp. isolated from slaughterhouse surfaces throughout meat chain production. Food Microbiol. 2015, 51, 33–44. [Google Scholar] [CrossRef]
- Wieland, N.; Boss, J.; Lettmann, S.; Fritz, B.; Schwaiger, K.; Bauer, J.; Hölzel, C.S. Susceptibility to disinfectants in antimicrobial-resistant and -susceptible isolates of Escherichia coli, Enterococcus faecalis and Enterococcus faecium from poultry-ESBL/AmpC-phenotype of E. coli is not associated with resistance to a quaternary ammonium. J. Appl. Microbiol. 2017, 122, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Cole, E.; Addison, R.; Rubino, J.; Leese, K.; Dulaney, P.; Newell, M.; Wilkins, J.; Gaber, D.; Wineinger, T.; Criger, D. Investigation of antibiotic and antibacterial agent cross-resistance in target bacteria from homes of antibacterial product users and nonusers. J. Appl. Microbiol. 2003, 95, 664–676. [Google Scholar] [CrossRef]
- Ghanem, B.; Haddadin, R.N. Multiple drug resistance and biocide resistance in Escherichia coli environmental isolates from hospital and household settings. Antimicrob. Resist. Infect. Control 2018, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide-Tolerant Listeria monocytogenes Isolates from German Food Production Plants Do Not Show Cross-Resistance to Clinically Relevant Antibiotics. Appl. Environ. Microbiol. 2019, 85, 85. [Google Scholar] [CrossRef] [Green Version]
- Russell, A.D. Similarities and differences in the responses of microorganisms to biocides. J. Antimicrob. Chemother. 2003, 52, 750–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdial, C.S.S. Prevenção de Infeções Nosocomiais: Controlo Bacteriológico de Superfícies Hospitalares da Unidade de Isolamento e Contenção Biológica do Hospital Escolar da FMV-ULisboa. Master’s Thesis, FMV-ULisboa, Lisbon, Portugal, 2020. [Google Scholar]
- European Committee for Standardization (CEN). Chemical Disinfection and Antiseptics: Quantitative Suspension Tests for the Evaluation of Bactericidal Activity of Chemical Disinfectants and Antiseptics Used in the Veterinary Area—Test Method and Requirements (Phase 2, Step 1); European Standard (EN) Document 1656; European Committee for Standardization: Brussels, Belgium, 2009. [Google Scholar]


{kind=link}
| E. coli (n = 3) | Pseudomonas spp. (n = 2) | Enterococcus spp. (n = 23) | S. pseudintermedius (n = 1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n = S% | n = I% | n = R% | n = S% | n = I% | n = R% | n = S% | n = I% | n = R% | n = S% | n = I% | n = R% | |
| AMC | 100 | 0 | 0 | 0 | 0 | 100 | 26.1 | 0 | 73.9 | 100 | 0 | 0 |
| AMP | 33.3 | 0 | 66.7 | - | - | - | 21.7 | 0 | 78.3 | 100 | 0 | 0 |
| CL | 100 | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 100 | 100 | 0 | 0 |
| CTX | 100 | 0 | 0 | 0 | 50 | 50 | 0 | 0 | 100 | 100 | 0 | 0 |
| CIP | 100 | 0 | 0 | 100 | 0 | 0 | 0 | 65.2 | 34.8 | 100 | 0 | 0 |
| ENR | 66.7 | 0 | 33.3 | 0 | 50 | 50 | 0 | 47.8 | 52.2 | 100 | 0 | 0 |
| CN | 100 | 0 | 0 | 100 | 0 | 0 | 0 | 38.1 | 61.9 | 100 | 0 | 0 |
| SXT | 100 | 0 | 0 | 50 | 0 | 50 | - | - | - | 0 | 0 | 100 |
| TE | 33.3 | 33.3 | 33.3 | 0 | 0 | 100 | 0 | 0.4 | 99.6 | 0 | 0 | 100 |
| Isolate Code | MDR | MIC (g/L) | MBC (g/L) | ||||
|---|---|---|---|---|---|---|---|
| NIS | LIS | HIS | NIS | LIS | HIS | ||
| C1 | - | 1.000 | 1.375 | 11.250 | 1.000 | 1.375 | 11.250 |
| C2 | - | 1.000 | 3.125 | 13.125 | 1.375 | 3.125 | 13.125 |
| C3 | - | 1.375 | 3.125 | 13.750 | 1.375 | 3.125 | 13.750 |
| 1.125 | 2.542 | 12.708 | 1.250 | 2.542 | 12.708 | ||
| σ | 0.177 | 0.825 | 1.062 | 0.177 | 0.825 | 1.062 | |
| P1 | - | 0.875 | 1.000 | 9.375 | 1.250 | 1.000 | 9.375 |
| P3 | - | 0.500 | 1.000 | 8.125 | 0.625 | 1.000 | 8.125 |
| 0.688 | 1.000 | 8.750 | 0.938 | 1.000 | 8.750 | ||
| σ | 0.188 | 0.000 | 0.625 | 0.313 | 0.000 | 0.625 | |
| E2 | - | 0.750 | 5.000 | >15.000 | 0.750 | 5.625 | >15.000 |
| E3 | ✓ | 0.500 | 5.000 | >15.000 | 0.625 | 5.000 | >15.000 |
| E5 | ✓ | 1.000 | 3.125 | >15.000 | 1.375 | 3.125 | >15.000 |
| E14 | - | 0.625 | 5.000 | >15.000 | 0.625 | 5.000 | >15.000 |
| E16 | ✓ | 0.625 | 5.000 | >15.000 | 0.750 | 5.000 | >15.000 |
| E17 | ✓ | 0.625 | 5.000 | >15.000 | 0.625 | 5.000 | >15.000 |
| E19 | ✓ | 0.500 | 5.625 | >15.000 | 0.500 | 5.625 | >15.000 |
| 0.661 | 4.821 | >15.000 | 0.750 | 4.911 | >15.000 | ||
| σ | 0.160 | 0.725 | - | 0.267 | 0.778 | - | |
| S3 | - | 0.625 | 2.500 | 14.375 | 0.625 | 2.500 | 14.375 |
| 0.625 | 2.500 | 14.375 | 0.625 | 2.500 | 14.375 | ||
| σ | - | - | - | - | - | - | |
| Isolate Code | MDR | Assay | MIC (g/L) | MBC (g/L) | ||||
|---|---|---|---|---|---|---|---|---|
| NIS | LIS | HIS | NIS | LIS | HIS | |||
| E2 | - | A | 0.750 | 5.000 | >15.000 | 0.750 | 5.625 | >15.000 |
| B | 0.625 | 7.500 | >15.000 | 0.625 | 7.500 | >15.000 | ||
| E3 | ✓ | A | 0.500 | 5.000 | >15.000 | 0.650 | 5.000 | >15.000 |
| B | 0.625 | 6.250 | >15.000 | 0.625 | 6.250 | >15.000 | ||
| E5 | ✓ | A | 1.000 | 3.125 | >15.000 | 1.375 | 3.125 | >15.000 |
| B | 0.500 | 6.875 | >15.000 | 0.500 | 7.500 | >15.000 | ||
| E14 | - | A | 0.625 | 5.000 | >15.000 | 0.625 | 5.000 | >15.000 |
| B | 0.500 | 6.875 | 14.125 | 0.500 | 6.875 | 14.125 | ||
| E16 | ✓ | A | 0.625 | 5.000 | >15.000 | 0.750 | 5.000 | >15.000 |
| B | 0.500 | 8.750 | >15.000 | 0.500 | 8.750 | >15.000 | ||
| E17 | ✓ | A | 0.625 | 5.000 | >15.000 | 0.625 | 5.000 | >15.000 |
| B | 0.500 | 8.750 | >15.000 | 0.500 | 8.750 | >15.000 | ||
| E19 | ✓ | A | 0.500 | 5.625 | >15.000 | 0.500 | 5.625 | >15.000 |
| B | 0.500 | 7.500 | >15.000 | 0.500 | 7.500 | >15.000 | ||
| A | 0.661 | 4.821 | >15.000 | 0.750 | 4.911 | >15.000 | ||
| B | 0.536 | 7.500 | >15.000 | 0.536 | 7.589 | >15.000 | ||
| σ | A | 0.160 | 0.725 | - | 0.267 | 0.778 | - | |
| B | 0.056 | 0.884 | - | 0.056 | 0.847 | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geraldes, C.; Verdial, C.; Cunha, E.; Almeida, V.; Tavares, L.; Oliveira, M.; Gil, S. Evaluation of a Biocide Used in the Biological Isolation and Containment Unit of a Veterinary Teaching Hospital. Antibiotics 2021, 10, 639. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060639
Geraldes C, Verdial C, Cunha E, Almeida V, Tavares L, Oliveira M, Gil S. Evaluation of a Biocide Used in the Biological Isolation and Containment Unit of a Veterinary Teaching Hospital. Antibiotics. 2021; 10(6):639. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060639
Chicago/Turabian StyleGeraldes, Catarina, Cláudia Verdial, Eva Cunha, Virgílio Almeida, Luís Tavares, Manuela Oliveira, and Solange Gil. 2021. "Evaluation of a Biocide Used in the Biological Isolation and Containment Unit of a Veterinary Teaching Hospital" Antibiotics 10, no. 6: 639. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10060639