
Molecular Epidemiology of Fosfomycin Resistant E. coli from a Pigeon Farm in China

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation, Fosfomycin-Resistant Gene Identification, and Antimicrobial Susceptibility Testing
2.2. Antimicrobial Susceptibility Testing
2.3. Molecular Analysis of Fosfomycin-Resistant E. coli
2.4. Whole-Genome Sequencing and Phylogenetic Analysis of fosA3-Positive Isolates
2.5. Data Availability
2.6. Ethical Statement
3. Results
3.1. Identification of fos Genes in Samples
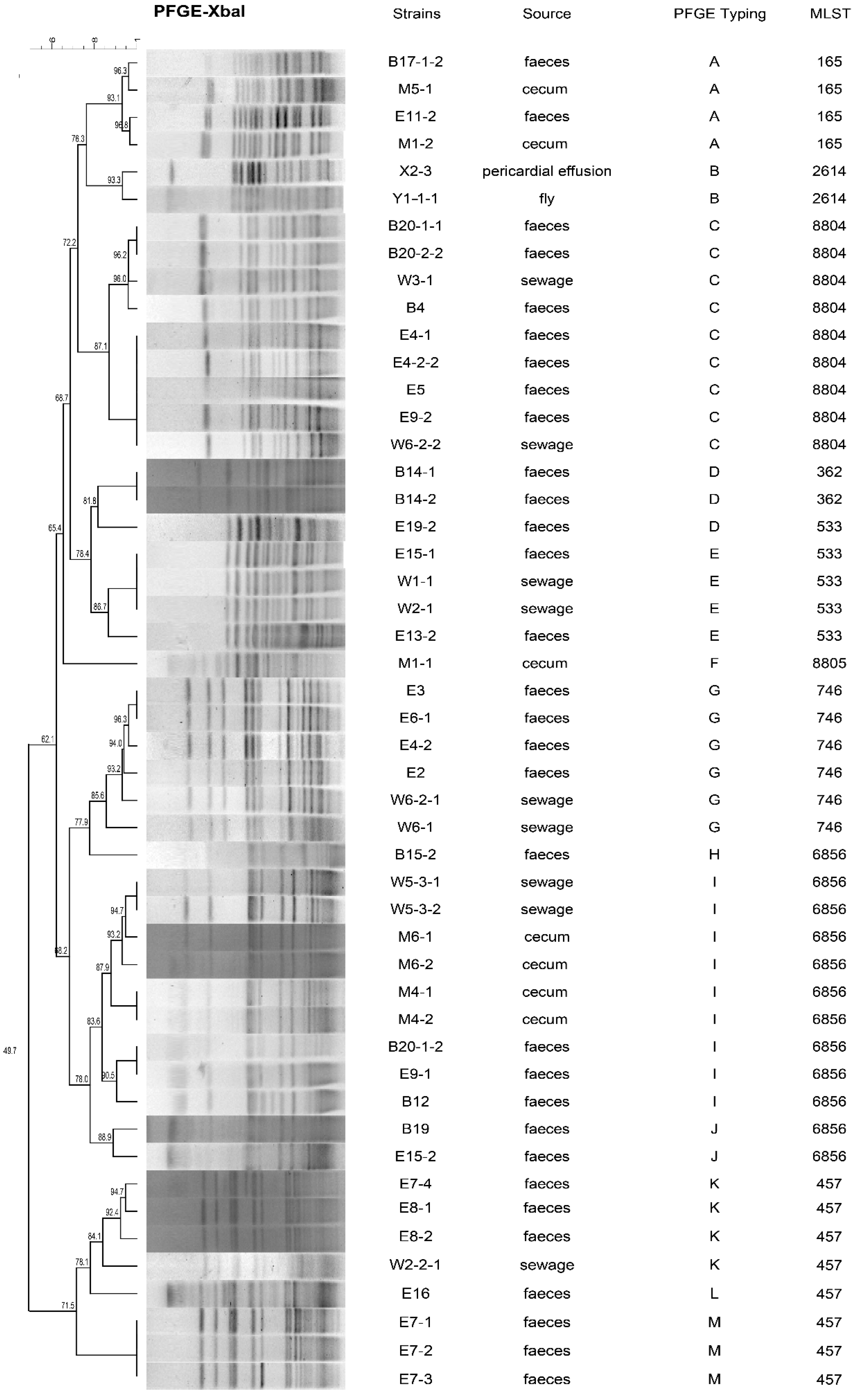
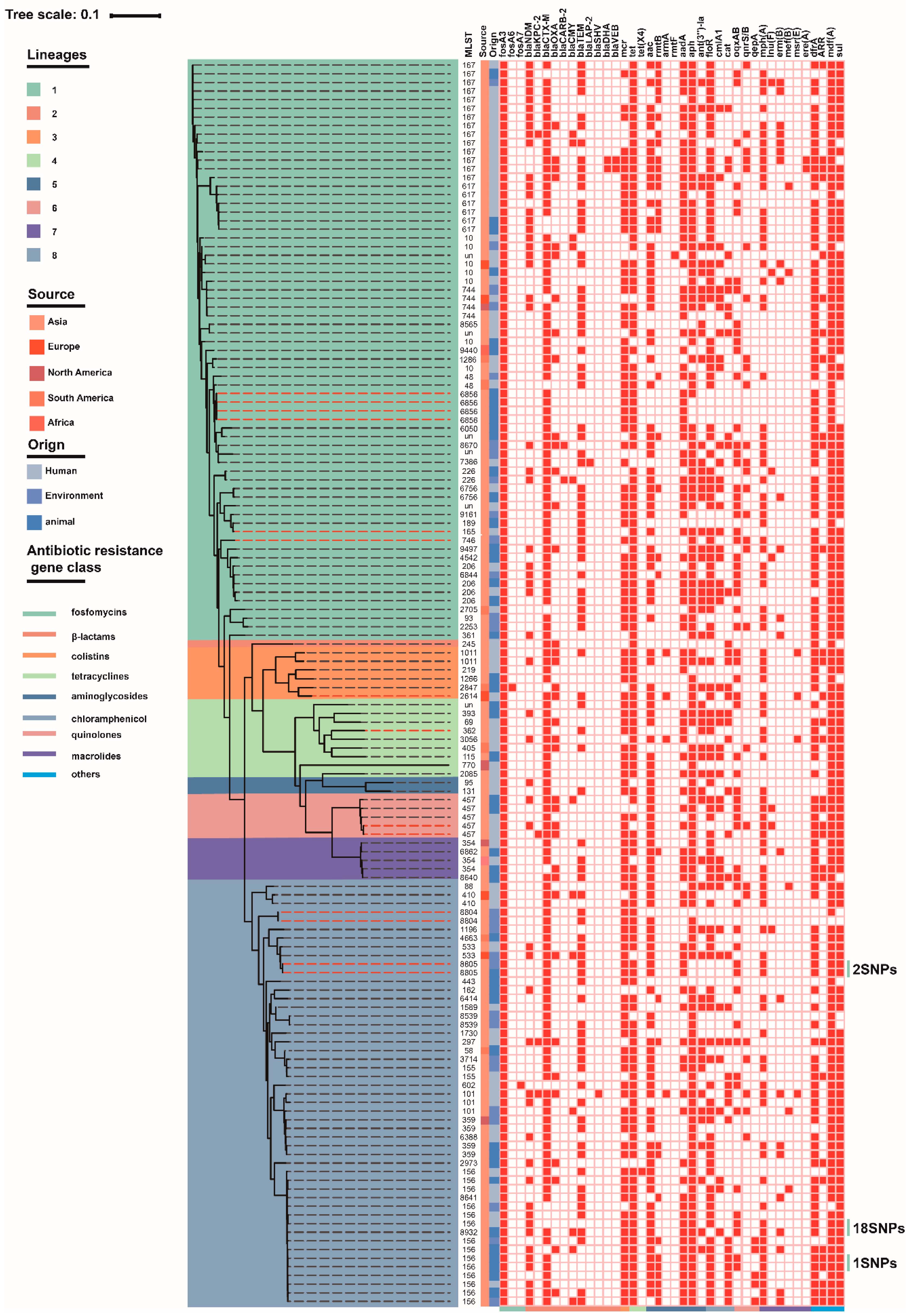
3.2. Whole-Genome Sequencing and Phylogenetic Analysis of fosA3-Positive E. coli Isolates
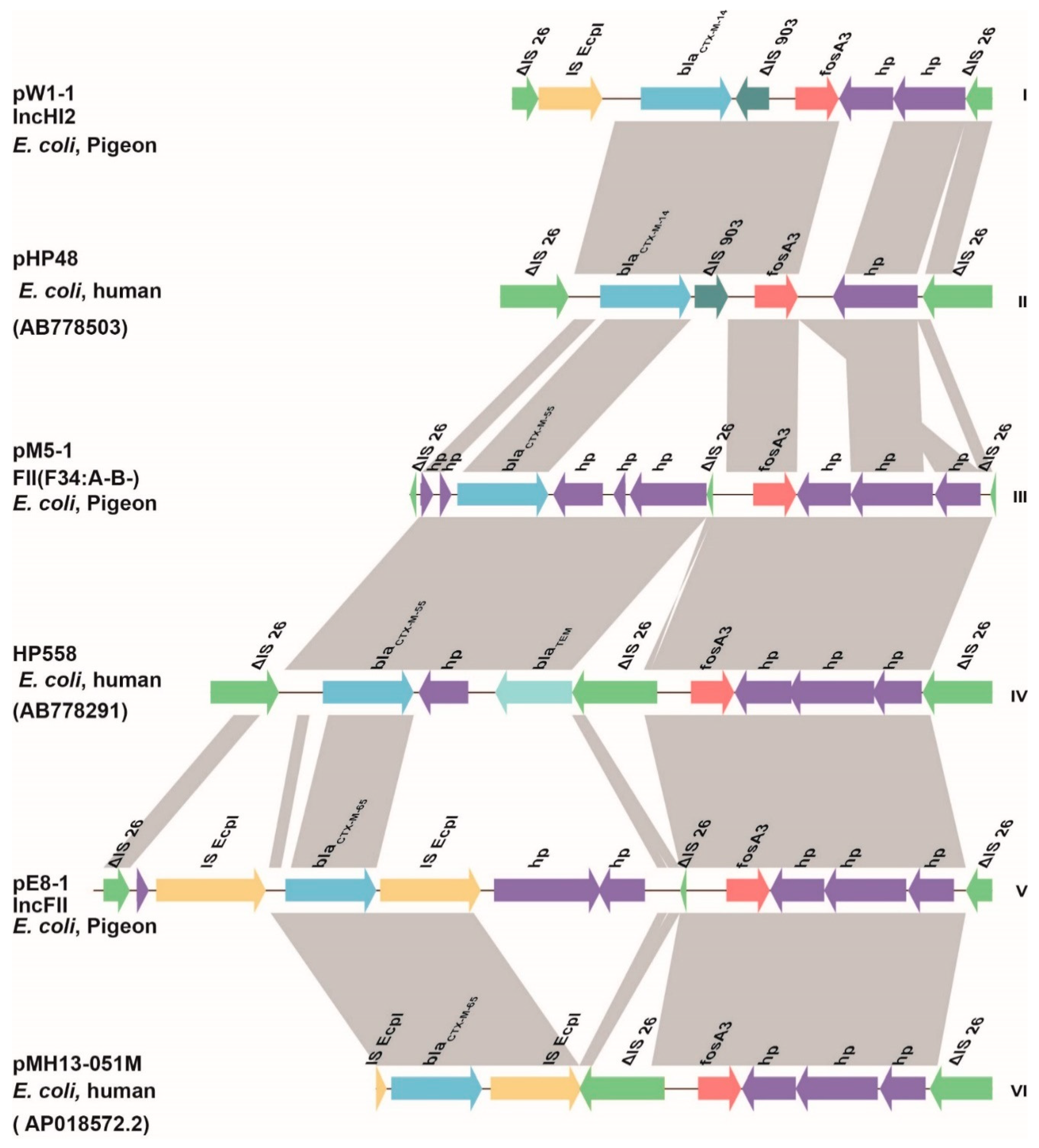
3.3. Genetic Environments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lletí, J.R.R. Fosfomycin in infections caused by multidrugresistant Gram-negative pathogens. Soc. Esp. Quimioter. 2019, 32, 45–54. [Google Scholar]
- Yang, T.Y.; Lu, P.L.; Tseng, S.P. Update on fosfomycin-modified genes in Enterobacteriaceae. J. Microbiol. Immunol. Infect. 2017, 52, 9–21. [Google Scholar] [CrossRef]
- He, D.; Liu, L.; Guo, B.; Wu, S.; Chen, X.; Wang, J.; Zeng, Z.; Liu, J. Chromosomal location of the fosA3 and blaCTX-M genes in Proteus mirabilis and clonal spread of Escherichia coli ST117 carrying fosA3-positive IncHI2/ST3 or F2: A-: B-plasmids in a chicken farm. Int. J. Antimicrob. Agents 2017, 49, 443–448. [Google Scholar] [CrossRef]
- Yao, H.; Wu, D.; Lei, L.; Shen, Z.; Wang, Y.; Liao, K. The detection of fosfomycin resistance genes in Enterobacteriaceae from pets and their owners. Vet. Microbiol. 2016, 193, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gu, X.; Zhang, J.; Yang, L.; Lu, Y.; Fang, L.; Jiang, H. Characterization of a fosA3 carrying IncC–IncN plasmid from a multidrug-resistant ST17 Salmonella Indiana isolate. Front. Microbiol. 2020, 11, 1582. [Google Scholar] [CrossRef]
- Haag-Wackernagel, D. Die taube-eine erfolgsgeschichte. vom liebling der götter zur eroberung der städte. Biol. Unserer Zeit 2011, 41, 44–52. [Google Scholar] [CrossRef]
- Ye, M.; Xu, M.; Chen, C.; He, Y.; Ding, M.; Ding, X.; Wei, W.; Yang, S.; Zhou, B. Expression analyses of candidate genes related to meat quality traits in squabs from two breeds of meat-type pigeon. J. Anim. Physiol. Anim. Nutr. 2017, 102, 727–735. [Google Scholar] [CrossRef]
- Teske, L.; Ryll, M.; Rubbenstroth, D.; Hänel, I.; Hartmann, M.; Kreienbrock, L.; Rautenschlein, S. Epidemiological investigations on the possible risk of distribution of zoonotic bacteria through apparently healthy homing pigeons. Avian Pathol. 2013, 42, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukerji, S.; Gunasekera, S.; Dunlop, J.N.; Stegger, M.; Jordan, D.; Laird, T.; Abraham, R.J.; Barton, M.; O’Dea, M.; Abraham, S. Implications of foraging and interspecies interactions of birds for carriage of escherichia coli strains resistant to critically important antimicrobials. Appl. Environ. Microbiol. 2020, 86, e01610–e01620. [Google Scholar] [CrossRef]
- Zhang, R.-M.; Liu, X.; Wang, S.-L.; Fang, L.-X.; Sun, J.; Liu, Y.-H.; Liao, X.-P. Distribution patterns of antibiotic resistance genes and their bacterial hosts in pig farm wastewater treatment systems and soil fertilized with pig manure. Sci. Total Environ. 2020, 758, 143654. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef]
- Walsh, F.; Duffy, B. The culturable soil antibiotic resistome: A community of multi-drug resistant bacteria. PLoS ONE 2013, 8, e65567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seng, P.; Drancourt, M.; Gouriet, F.; La Scola, B.; Fournier, P.; Rolain, J.M.; Raoult, D. Ongoing revolution in bacteriology: Routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Infect. Dis. 2009, 49, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Kawamura, K.; Nakane, K.; Wachino, J.; Arakawa, Y. First detection of fosfomycin resistance gene fosA3 in CTX-M-producing Escherichia coli isolates from healthy individuals in Japan. Microb. Drug Resist. 2013, 19, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Escherichia coli Sequence Typing. Available online: https://bigsdb.pasteur.fr/ecoli/ecoli.htm (accessed on 27 August 2016).
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing—Twenty-Eighth Edition; CLSI: Annapolis Junction, MD, USA, 2018; p. M100. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 7.0; EUCAST: Växjö, Sweden, 2017. [Google Scholar]
- Zhang, R.; Wan, Y.; Liu, Z.; Li, J.; Yin, W.; Lei, L.; Wu, C.; Shen, J. Characterization of NDM-1-producing carbapenemase in Acinetobacter spp. and E. coli isolates from diseased pigs. Front. Agric. Sci. Eng. 2015, 2, 223. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, R.; Wang, L.; Sun, R.; Bai, S.; Han, L.; Fang, L.; Sun, J.; Liu, Y.; Liao, X. Molecular epidemiology of carbapenemase-producing Escherichia coli from duck farms in south-east coastal China. J. Antimicrob. Chemother. 2020, 76, 322–329. [Google Scholar] [CrossRef]
- Madden, T. The BLAST Sequence Analysis Tool. 2013. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 15 March 2013).
- Inouye, M.; Harriet Dashnow, L.A.R.; Schultz, M.B.; Pope, B.J.; Tpmita, T.; Zobel, J.; Holt, K.E. SRST2: Rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 2014, 6, 90. [Google Scholar] [CrossRef] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [Green Version]
- Tseemann. Snippy (Version 4.4.5). Available online: https://github.com/tseemann/snippy (accessed on 4 January 2020).
- Cheng, L.; Connor, T.R.; Sirén, J.; Aanensen, D.M.; Corander, J. Hierarchical and spatially explicit clustering of DNA sequences with BAPS software. Mol. Biol. Evol. 2013, 30, 1224–12248. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanaa, A.A.; Lo, C.-C.; Li, P.-E.; Davenport, K.D.; Chain, P.S.G. Chain. From raw reads to trees: Whole genome SNP phylogenetics across the tree of life. bioRxiv 2015. [Google Scholar] [CrossRef] [Green Version]
- Lei, C.-H.; Chen, Y.-A.; Kang, Z.H.; Kong, L.I.; Wang, H.-O. Characterization of a Novel SXT/R391 integrative and conjugative element carrying cfr, blaCTX-M-65, fosA3, and aac(6′)-Ib-cr in Proteus mirabilis. Antimicrob. Agents Chemother. 2018, 62, e00849-18. [Google Scholar] [CrossRef] [Green Version]
- Cyoia, C.P.S.; Koga, V.L.; Nishio, E.K.; Houle, S.; Dozois, C.M.; de Brito, K.C.T.; de Brito, B.G.; Nakazato, G.; Kobayashi, R.K.T. Distribution of ExPEC virulence factors, blaCTX-M, fosA3, and mcr-1 in Escherichia coli isolated from commercialized chicken carcasses. Front. Microbiol. 2018, 9, 3254. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Huang, X.; Deng, Y.; He, L.; Yang, T.; Zeng, Z.; Chen, Z.; Liu, J. Dissemination of the fosfomycin resistance gene fosA3 with CTX-M β-lactamase genes and rmtB carried on IncFII Plasmids among Escherichia coli isolates from pets in China. Antimicrob. Agents Chemother. 2011, 56, 2135–2138. [Google Scholar] [CrossRef] [Green Version]
- Hou, J. Detection of the plasmid-encoded fosfomycin resistance gene fosA3 in Escherichia coli of food-animal origin. J. Antimicrob. Chemother. 2012, 68, 766–770. [Google Scholar] [CrossRef]
- Ho, P.L.; Chan, J.; Lo, W.U.; Law, P.Y.; Li, Z.; Lai, E.L.; Chow, K.H. Dissemination of plasmid-mediated fosfomycin resistance fosA3 among multidrug-resistant Escherichia coli from livestock and other animals. J. Appl. Microbiol. 2013, 114, 695–702. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Fang, L.; Ge, Q.; Wang, D.; He, B.; Lu, J.; Zhong, Z.; Wang, X.; Yu, Y.; Lian, X.; et al. Emergence of fosA3 and bla CTX-M-14 in multidrug-resistant citrobacter freundii isolates from flowers and the retail environment in China. Front. Microbiol. 2021, 12, 586504. [Google Scholar] [CrossRef]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Chew, K.L.; Octavia, S.; Ng, O.T.; Marimuthu, K.; Venkatachalam, I.; Cheng, B.; Lin, R.T.P.; Teo, J.W.P. Challenge of drug resistance in Pseudomonas aeruginosa: Clonal spread of NDM-1-positive ST-308 within a tertiary hospital. J. Antimicrob. Chemother. 2019, 74, 2220–2224. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, L.; Lu, X.-Q.; Liu, X.-W.; Liao, M.-N.; Sun, R.-Y.; Xie, Y.; Liao, X.-P.; Liu, Y.-H.; Sun, J.; Zhang, R.-M. Molecular Epidemiology of Fosfomycin Resistant E. coli from a Pigeon Farm in China. Antibiotics 2021, 10, 777. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070777
Han L, Lu X-Q, Liu X-W, Liao M-N, Sun R-Y, Xie Y, Liao X-P, Liu Y-H, Sun J, Zhang R-M. Molecular Epidemiology of Fosfomycin Resistant E. coli from a Pigeon Farm in China. Antibiotics. 2021; 10(7):777. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070777
Chicago/Turabian StyleHan, Lu, Xiao-Qing Lu, Xu-Wei Liu, Mei-Na Liao, Ruan-Yang Sun, Yao Xie, Xiao-Ping Liao, Ya-Hong Liu, Jian Sun, and Rong-Min Zhang. 2021. "Molecular Epidemiology of Fosfomycin Resistant E. coli from a Pigeon Farm in China" Antibiotics 10, no. 7: 777. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070777