
Antimicrobial Resistance Profiling of Biofilm Forming Non Typhoidal Salmonella enterica Isolates from Poultry and Its Associated Food Products from Pakistan


 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Isolation of Salmonella Isolates
2.3. Molecular Detection of Salmonella using Multiplex PCR
2.4. Antimicrobial Susceptibility Assay
2.5. Molecular Detection of Antibiotic Resistance and Virulence Genes
2.6. Biofilm Characterization of Salmonella Isolates
2.7. Phenotypic Characterization of Extracellular Virulence Factors in Salmonella Isolates
2.8. Statistical Analysis
3. Results
3.1. Prevalence and Isolation of Salmonella Enterica Serovars
3.2. Antimicrobial Susceptibility Assay
3.3. Distribution of Antibiotic Resistance and Virulence Genes in Salmonella Isolates
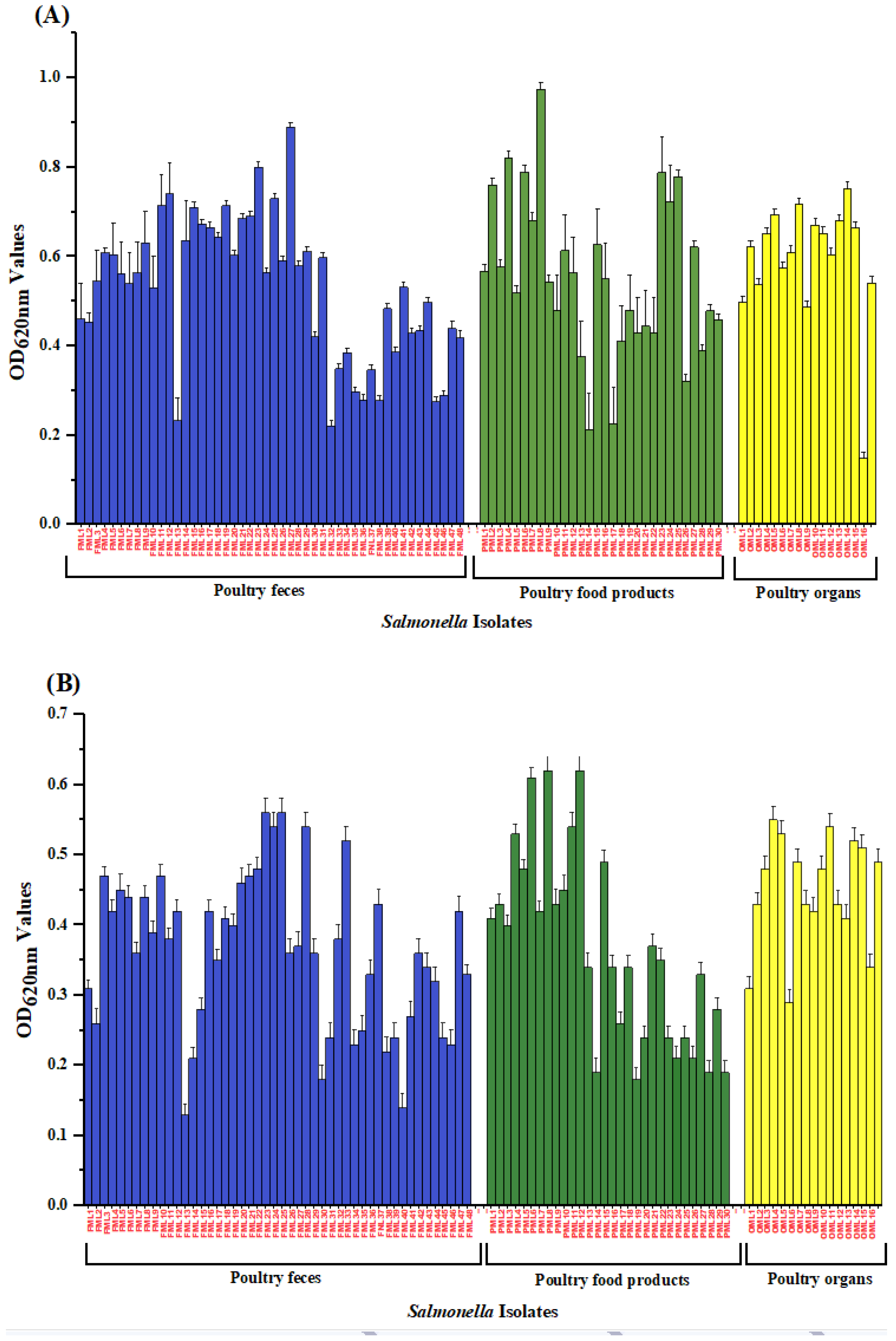
3.4. Biofilm Formation Potential of Salmonella Isolates
3.5. Correlation between the Number of Isolates Resistant to Antibiotics and the Ability to Produce Biofilms at Different Temperatures
3.6. Phenotypic Characterization of Extracellular Virulence Factors in Salmonella Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gharieb, R.; Tartor, Y.H.; Khedr, M.H.E. Non-Typhoidal Salmonella in poultry meat and diarrhoeic patients: Prevalence, antibiogram, virulotyping, molecular detection and sequencing of class I integrons in multidrug resistant strains. Gut Pathog. 2015, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Guibourdenche, M.; Roggentin, P.; Mikoleit, M.; Fields, P.I.; Bockemühl, J.; Grimont, P.; Weill, F.-X. Supplement 2003–2007 (No. 47) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2010, 161, 26–29. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Veter. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [Green Version]
- Majowicz, S.; Musto, J.; Scallan, E.; Angulo, F.; Kirk, M.; O’Brien, S.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease “Burden of Illness” Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [Green Version]
- Mouttotou, N.; Ahmad, S.; Kamran, Z.; Koutoulis, K.C. Prevalence, Risks and Antibiotic Resistance of Salmonella in Poultry Production Chain. Curr. Top. Salmonella Salmonellosis 2017, 2017, 134–215. [Google Scholar] [CrossRef]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Thai, T.H.; Hirai, T.; Lan, N.T.; Yamaguchi, R. Antibiotic resistance profiles of Salmonella serovars isolated from retail pork and chicken meat in North Vietnam. Int. J. Food Microbiol. 2012, 156, 147–151. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Venkitanarayanan, K.; Kollanoor, J.A. Antibiotic-resistant Salmonella in the food supply and the potential role of antibiotic alternatives for control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.; Wu, C.; Wu, C.; Qi, J.; Wang, Y.; Wang, H.; Liu, Y.; Shen, J. Serotype distribution and antibiotic resistance of Salmonella in food-producing animals in Shandong province of China, 2009 and 2012. Int. J. Food Microbiol. 2014, 180, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Clothier, K.A.; Kim, P.; Mete, A.; Hill, A.E. Frequency, serotype distribution, and antimicrobial susceptibility patterns of Salmonella in small poultry flocks in California. J. Veter. Diagn. Investig. 2018, 30, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Reynolds, J.; Karp, B.; Tate, H.; Cray, P.; Plumblee, J.R.; Hoekstra, R.M.; Whichard, J.M.; Mahon, B.E. Ceftriaxone-Resistant Nontyphoidal Salmonella from Humans, Retail Meats, and Food Animals in the United States, 1996–2013. Foodborne Pathog. Dis. 2017, 14, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, J.; Wu, Q.; Zhang, J.; Liu, S.; Guo, W.; Cai, S.; Yu, S. Prevalence, antimicrobial resistance and genetic diversity of Salmonella isolated from retail ready-to-eat foods in China. Food Control. 2016, 60, 50–56. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuchi, A.; Inagaki, M.; Sugiyama, M.; Usui, M.; Asai, T. Association of Salmonella Serotypes with Quinolone Resistance in Broilers. Food Saf. 2018, 6, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Nde, C.W.; Fakhr, M.K.; Doetkott, C.; Logue, C.M. An evaluation of conventional culture, invA PCR, and the real-time PCR iQ-Check kit as detection tools for Salmonella in naturally contaminated premarket and retail turkey. J. Food Prot. 2008, 71, 386–391. [Google Scholar] [CrossRef]
- Gunasegaran, T.; Rathinam, X.; Kasi, M.; Sathasivam, K.; Sreenivasan, S.; Subramaniam, S. Isolation and identification of Salmonella from curry samples and its sensitivity to commercial antibiotics and aqueous extracts of Camelia sinensis (L.) and Trachyspermum ammi (L.). Asian Pac. J. Trop. Biomed. 2011, 1, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Soumet, C.; Ermel, G.; Rose, V.; Rose, N.; Drouin, P.; Salvat, G.; Colin, P. Identification by a multiplex PCR-based assay ofSalmonellaTyphimurium andSalmonellaEnteritidis strains from environmental swabs of poultry houses. Lett. Appl. Microbiol. 1999, 29, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Moe, A.Z.; Paulsen, P.; Pichpol, D.; Fries, R.; Irsigler, H.; Baumann, M.P.O.; Oo, K.N. Prevalence and Antimicrobial Resistance of Salmonella Isolates from Chicken Carcasses in Retail Markets in Yangon, Myanmar. J. Food Prot. 2017, 80, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Miranda, J.; Vazquez, B.; Cepeda, A.; Franco, C. Biofilm formation, phenotypic production of cellulose and gene expression in Salmonella enterica decrease under anaerobic conditions. Int. J. Food Microbiol. 2016, 238, 63–67. [Google Scholar] [CrossRef]
- Marangoni, F.; Corsello, G.; Cricelli, C.; Ferrara, N.; Ghiselli, A.; Lucchin, L.; Poli, A. Role of poultry meat in a balanced diet aimed at maintaining health and wellbeing: An Italian consensus document. Food Nutr. Res. 2015, 59, 27606. [Google Scholar] [CrossRef] [Green Version]
- Galloway, W.R.; Hodgkinson, J.; Bowden, S.; Welch, M.; Spring, D.R. Applications of small molecule activators and inhibitors of quorum sensing in Gram-negative bacteria. Trends Microbiol. 2012, 20, 449–458. [Google Scholar] [CrossRef]
- Odoch, T.; Sekse, C.; L’Abee-Lund, T.M.; Hansen, H.C.H.; Kankya, C.; Wasteson, Y. Diversity and Antimicrobial Resistance Genotypes in Non-Typhoidal Salmonella Isolates from Poultry Farms in Uganda. Int. J. Environ. Res. Public Health 2018, 15, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babapour, E.; Haddadi, A.; Mirnejad, R.; Angaji, S.A.; Amirmozafari, N. Biofilm formation in clinical isolates of nosocomial Acinetobacter baumannii and its relationship with multidrug resistance. Asian Pac. J. Trop. Biomed. 2016, 6, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vukovic, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Eguale, T.; Gebreyes, W.A.; Asrat, D.; Alemayehu, H.; Gunn, J.S.; Engidawork, E. Non-typhoidal Salmonella serotypes, antimicrobial resistance and co-infection with parasites among patients with diarrhea and other gastrointestinal complaints in Addis Ababa, Ethiopia. BMC Infect. Dis. 2015, 15, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Dahiya, S.P.; Ratwan, P. Backyard poultry farming in India: A tool for nutritional security and women empowerment. Biol. Rhythm. Res. 2019. [Google Scholar] [CrossRef]
- Michael, G.; Schwarz, S. Antimicrobial resistance in zoonotic nontyphoidal Salmonella: An alarming trend? Clin. Microbiol. Infect. 2016, 22, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Duc, V.M.; Nakamoto, Y.; Fujiwara, A.; Toyofuku, H.; Obi, T.; Chuma, T. Prevalence of Salmonella in broiler chickens in Kagoshima, Japan in 2009 to 2012 and the relationship between serovars changing and antimicrobial resistance. BMC Veter. Res. 2019, 15, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wajid, M.; Awan, A.B.; Saleemi, M.K.; Weinreich, J.; Schierack, P.; Sarwar, Y.; Ali, A. Multiple Drug Resistance and Virulence Profiling ofSalmonella entericaSerovars Typhimurium and Enteritidis from Poultry Farms of Faisalabad, Pakistan. Microb. Drug Resist. 2019, 25, 133–142. [Google Scholar] [CrossRef]
- Li, B.; Liu, C.; Liu, L.; Li, S.; Fan, N.; Hou, H.; Jin, J.; Xing, Y. [Prevalence and etiologic agent of Salmonella in livestock and poultry meats in Huai’an City during 2015–2016]. Wei Sheng Yan Jiu J. Hyg. Res. 2018, 47, 260–300. [Google Scholar]
- Stock, I.; Wiedemann, B. Natural antibiotic susceptibility of Salmonella enterica strains. Int. J. Antimicrob. Agents 2000, 16, 211–217. [Google Scholar] [CrossRef]
- Abd-Elghany, S.; Sallam, K.; Abd-Elkhalek, A.; Tamura, T. Occurrence, genetic characterization and antimicrobial resistance of Salmonella isolated from chicken meat and giblets. Epidem. Infect. 2015, 143, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tayeb, M.A.; Ibrahim, A.S.; Al-Salamah, A.A.; Almaary, K.S.; Elbadawi, Y.B. Prevalence, serotyping and antimicrobials resistance mechanism of Salmonella enterica isolated from clinical and environmental samples in Saudi Arabia. Braz. J. Microbiol. 2017, 48, 499–508. [Google Scholar] [CrossRef]
- Gad, A.H.; Abo-Shama, U.H.; Harclerode, K.K.; Fakhr, M.K. Prevalence, Serotyping, Molecular Typing, and Antimicrobial Resistance of Salmonella Isolated From Conventional and Organic Retail Ground Poultry. Front. Microbiol. 2018, 9, 2653. [Google Scholar] [CrossRef] [PubMed]
- Temkin, E.; Adler, A.; Lerner, A.; Carmeli, Y. Carbapenem-resistant Enterobacteriaceae: Biology, epidemiology, and management. Ann. N. Y. Acad. Sci. 2014, 1323, 22–42. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Guerra, B.; Rodicio, M.R. Resistance to Carbapenems in Non-Typhoidal Salmonella enterica Serovars from Humans, Animals and Food. Veter. Sci. 2018, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, E.J.; Shakoor, S.; Page, A.J.; Qamar, F.N.; Judge, K.; Saeed, D.K.; Wong, V.K.; Dallman, T.J.; Nair, S.; Baker, S.; et al. Emergence of an Extensively Drug-Resistant Salmonella enterica Serovar Typhi Clone Harboring a Promiscuous Plasmid Encoding Resistance to Fluoroquinolones and Third-Generation Cephalosporins. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Soomro, A.H.; Khaskheli, M.; Bhutto, M.B.; Shah, G.; Memon, A.; Dewani, P. Prevalence and antimicrobial resistance of Salmonella serovars isolated from poultry meat in Hyderabad, Pakistan. Turk. J. Veter. Anim. Sci. 2011, 34, 455–460. [Google Scholar]
- Borges, K.A.; Furian, T.Q.; Souza SNd Salle, C.T.P.; Moraes, H.L.d.S.; Nascimento, V.P.D. Antimicrobial resistance and molecular characterization of Salmonella enterica serotypes isolated from poultry sources in Brazil. Braz. J. Poult. Sci. 2019, 21. [Google Scholar] [CrossRef]
- Budiati, T.; Rusul, G.; Wan-Abdullah, W.N.; Arip, Y.M.; Ahmad, R.; Thong, K.L.J.A. Prevalence, antibiotic resistance and plasmid profiling of Salmonella in catfish (Clarias gariepinus) and tilapia (Tilapia mossambica) obtained from wet markets and ponds in Malaysia. Aquaculture 2013, 372, 127–132. [Google Scholar] [CrossRef]
- Beshiru, A.; Igbinosa, I.H.; Igbinosa, E.O. Prevalence of antimicrobial resistance and virulence gene elements of Salmonella serovars from ready-to-eat (RTE) shrimps. Front. Microbiol. 2019, 10, 1613. [Google Scholar] [CrossRef] [Green Version]
- Deekshit, V.K.; Kumar, B.K.; Rai, P.; Rohit, A.; Karunasagar, I. Simultaneous detection of Salmonella pathogenicity island 2 and its antibiotic resistance genes from seafood. J. Microbiol. Methods 2013, 93, 233–238. [Google Scholar] [CrossRef]
- Biffi, C.P.; Stefani, L.M.; Miletti, L.C.; Matiello, C.A.; Backes, R.G.; Almeida, J.M.; Neves, G.B. Phenotypic and genotypic resistance profile of Salmonella Typhimurium to antimicrobials commonly used in poultry. Braz. J. Poult. Sci. 2014, 16, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yang, J.; Zhang, B.; Sun, S.; Chang, W. Characterization of Integrons and Resistance Genes in Salmonella Isolates from Farm Animals in Shandong Province, China. Front. Microbiol. 2017, 8, 1300. [Google Scholar] [CrossRef]
- Borges, K.A.; Furian, T.Q.; Borsoi, A.; Moraes, H.L.; Salle, C.T.P.; Nascimento, V.P.D. Detection of virulence-associated genes in Salmonella Enteritidis isolates from chicken in South of Brazil. Pesquisa Veterinária Brasileira 2013, 33, 1416–1422. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Tahoun, A.; El-Gohary, A.E.; El-Abasy, M.; El-Khayat, F.; Gillespie, T.; Kitade, Y.; Hafez, H.M.; Neubauer, H.; El-Adawy, H. Epidemiological, molecular characterization and antibiotic resistance of Salmonella enterica serovars isolated from chicken farms in Egypt. Gut Pathogens. 2017, 9, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, K.L.; Threlfall, E.J. Frequency and polymorphism of sopE in isolates of Salmonella enterica belonging to the ten most prevalent serotypes in England and Wales. J. Med. Microbiol. 2004, 53, 539–543. [Google Scholar] [CrossRef]
- Han, J.; Gokulan, K.; Barnette, D.; Khare, S.; Rooney, A.W.; Deck, J.; Nayak, R.; Stefanova, R.; Hart, M.E.; Foley, S.L. Evaluation of Virulence and Antimicrobial Resistance inSalmonella entericaSerovar Enteritidis Isolates from Humans and Chicken- and Egg-Associated Sources. Foodborne Pathog. Dis. 2013, 10, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.M.; Gamazo, C.; Penadés, J.R.; Lasa, I. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol. Microbiol. 2005, 58, 1322–1339. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.L.C.; de Araújo Soares, R.; Piccoli, R.H. A Weibull model to describe antimicrobial kinetics of oregano and lemongrass essential oils against Salmonella Enteritidis in ground beef during refrigerated storage. Meat Sci. 2013, 93, 645–651. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New Perspectives on the Use of Phytochemicals as an Emergent Strategy to Control Bacterial Infections Including Biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef]
- Jensen, M.B.F.; Schjørring, S.; Björkman, J.T.; Torpdahl, M.; Litrup, E.; Nielsen, E.M.; Niskanen, T. External quality assessment for molecular typing of Salmonella 2013–2015: Performance of the European national public health reference laboratories. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1923–1932. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Petri, N.; Daron, C.; Pereira, R.; Mendoza, M.; Hassan, H.M.; Koci, M.D. Poultry Body Temperature Contributes to Invasion Control through Reduced Expression of Salmonella Pathogenicity Island 1 Genes in Salmonella enterica Serovars Typhimurium and Enteritidis. Appl. Environ. Microbiol. 2015, 81, 8192–8201. [Google Scholar] [CrossRef] [Green Version]
- Runkel, S.; Wells, H.C.; Rowley, G. Living with stress: A lesson from the enteric pathogen Salmonella enterica. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 83, pp. 87–144. [Google Scholar]
- Simm, R.; Ahmad, I.; Rhen, M.; Le Guyon, S.; Römling, U. Regulation of biofilm formation in Salmonella enterica serovar Typhimurium. Future Microbiol. 2014, 9, 1261–1282. [Google Scholar] [CrossRef] [PubMed]
- Baugh, S.; Ekanayaka, A.S.; Piddock, L.J.V.; Webber, M.A. Loss of or inhibition of all multidrug resistance efflux pumps of Salmonella enterica serovar Typhimurium results in impaired ability to form a biofilm. J. Antimicrob. Chemother. 2012, 67, 2409–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frees, D.; Brøndsted, L.; Ingmer, H. Bacterial proteases and virulence. In Regulated Proteolysis in Microorganisms; Springer: Berlin/Heidelberg, Germany, 2013; pp. 161–192. [Google Scholar]
- Rosenau, F.; Isenhardt, S.; Gdynia, A.; Tielker, D.; Schmidt, E.; Tielen, P.; Schobert, M.; Jahn, D.; Wilhelm, S.; Jaeger, K.-E. Lipase LipC affects motility, biofilm formation and rhamnolipid production in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2010, 309, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, N.K.; Wattal, C.; Chandy, S.J.; Arora, S.K.; Gupta, U.; Kotwani, A. Situation Analysis Antibiotic Use and Resistance in India; Public Health Foundation of India, and Center for Disease Dynamics, Economics and Policy: New Delhi, India, 2011. [Google Scholar]
- Pruden, A.; Larsson, D.J.; Amézquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management Options for Reducing the Release of Antibiotics and Antibiotic Resistance Genes to the Environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target Sequence | Target Strain | Primer Sets | Length | Primer Sequence 5′----3′ | Amplification Region | Reference |
|---|---|---|---|---|---|---|
| Random Sequence | Salmonella spp. | ST11 ST15 | 24 24 | GCCAACCATTGCTAAATTGGCGCA GGTAGAAATTCCCAGCGGGTACTGG | 429 | [17] |
| fliC gene | Salmonella Typhimurium | Fli15 Tym | 22 22 | CGGTGTTGCCCAGGTTGGTAAT ACTCTTGCTGGCGGTGCGACTT | 559 | [17] |
| sefA gene | Salmonella Enteritidis | Sef 167 Sef 478 | 20 20 | AGGTTCAGGCAGCGGTTACT GGGACATTTAGCGTTTCTTG | 312 | [17] |
| Genes | Sequences (5′-3′) | Annealing Temp. (°C) | Amplicon Size | References |
|---|---|---|---|---|
| bla PSE-1 | CGCTTCCCGTTAACAAGTAC CTGGTTCATTTCAGATAGCG | 50 | 430 | [18] |
| bla CMY-2 | TGGCCAGAACTGACAGGCAAA TTTCTCCTGAACGTGGCTGGC | 57 | 870 | |
| bla TEM-1 | CAGCGGTAAGATCCT TGAGA ACTCGCCGTCGTGTAGATAA | 55 | 643 | [19] |
| bla OxA-1 | ATGAAAAACACAATACATATC AATTTAGTGTGTTTAGAATGG | 50 | 830 | |
| bla NDM-1 | GGG CAG TCG CTT CCA ACG GT GTA GTG CTC AGT GTC GGC AT | 58 | 475 | [20] |
| ampC | AACACACTGATTGCGTCTGAC CTGGGCCTCATCGTCAGTTA | 60 | 1226 | [9] |
| SopE | ACACACTTTCCACGAGGAAGCG GGATGCCTTCTGATGTTGACTGG | 50 | 398 | [21] |
| Sr. No. | Sample Source | No. of Samples (n) | Positive Samples (%) | Serovar (s) Isolated | No. of Serovar out of Positive Sample (%) |
|---|---|---|---|---|---|
| 1 | Poultry feces | 180 | 48 (26.7) | S.Typhimurium | 9 (18.75) |
| S. Enteritidis | 8 16.6) | ||||
| Other Salmonella | 31 (64.6) | ||||
| 2 | Poultry organs | 70 | 17 (24.3) | S.Typhimurium | 1 (5.8) |
| S. Enteritidis | 2 (11.7) | ||||
| Other Salmonella | 14 (82.3) | ||||
| 3 | Poultry Meat | 60 | 21 (35) | S.Typhimurium | 2 (9.5) |
| S. Enteritidis | 1 (4.7) | ||||
| Other Salmonella | 18 (85.7) | ||||
| 4 | Poultry eggs | 60 | 9 (15) | S.Typhimurium | 2 (22.2) |
| S.Enteritidis | 1 (11.1) | ||||
| Other Salmonella | 6 (66.6) |
| Salmonella spp. | Temperature (°C) | Weak Biofilm | Moderate Biofilm | Strong Biofilm | No Biofilm |
|---|---|---|---|---|---|
| S. Typhimurium (n = 14) | 30 | 3 (21.4) | 4 (28.5) | 7 (50) | 0 (0) |
| 37 | 1 (8) | 2 (14.2) | 11 (78.5) | 0 (0) | |
| S. Enteritidis (n = 12) | 30 | 2 (16.6) | 7 (58.3) | 3 (25) | 0 (0) |
| 37 | 2 (16.6) | 5 (41.7) | 5 (41.7) | 0 (0) | |
| Other Salmonella spp. (n = 69) | 30 | 18 (26.1) | 30 (43.4) | 21 (30.4) | 0 (0) |
| 37 | 11 (15.9) | 20 (28.9) | 38 (55.1) | 0 (0) |
| Salmonella spp. | DNA Degrading Activity | Hemolytic Activity | Lipase Activity | Protease Activity | Swimming Motility | Swarming Motility |
|---|---|---|---|---|---|---|
| S. Typhimurium (n = 14) | 14 (100) | 13 (92.8) | 11 (78.6) | 10 (83) | 14 (100) | 14 (100) |
| S. Enteritidis (n = 12) | 12 (100) | 12 (100) | 11 (91.6) | 10 (83) | 12 (100) | 12 (100) |
| Other Salmonella spp. (n = 69) | 67 (97.1) | 68 (98.5) | 65 (94.2) | 68 (98.5) | 69 (100) | 69 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddique, A.; Azim, S.; Ali, A.; Andleeb, S.; Ahsan, A.; Imran, M.; Rahman, A. Antimicrobial Resistance Profiling of Biofilm Forming Non Typhoidal Salmonella enterica Isolates from Poultry and Its Associated Food Products from Pakistan. Antibiotics 2021, 10, 785. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070785
Siddique A, Azim S, Ali A, Andleeb S, Ahsan A, Imran M, Rahman A. Antimicrobial Resistance Profiling of Biofilm Forming Non Typhoidal Salmonella enterica Isolates from Poultry and Its Associated Food Products from Pakistan. Antibiotics. 2021; 10(7):785. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070785
Chicago/Turabian StyleSiddique, Abubakar, Sara Azim, Amjad Ali, Saadia Andleeb, Aitezaz Ahsan, Muhammad Imran, and Abdur Rahman. 2021. "Antimicrobial Resistance Profiling of Biofilm Forming Non Typhoidal Salmonella enterica Isolates from Poultry and Its Associated Food Products from Pakistan" Antibiotics 10, no. 7: 785. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10070785