
Assessing the Bacterial Community Composition of Bivalve Mollusks Collected in Aquaculture Farms and Respective Susceptibility to Antibiotics

 , , and
, , and
Abstract
:1. Introduction
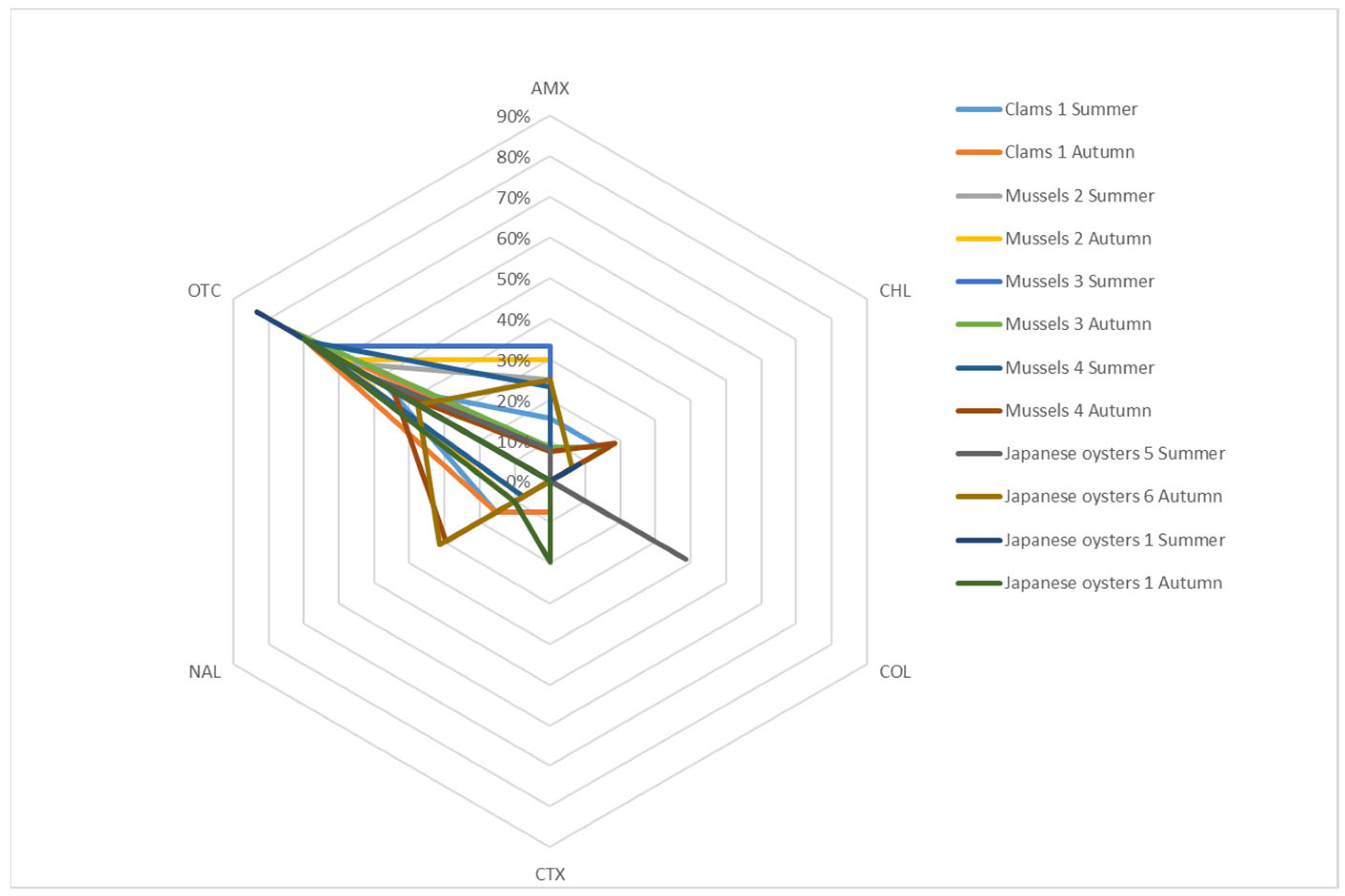
2. Results
2.1. Bacterial Diversity in Clams Samples
2.2. Bacterial Diversity in Mussels samples
2.3. Bacterial Diversity in Japanese Oysters Samples
2.4. Initial Evaluation of Decreased Susceptibilities
2.5. Antibiotic Susceptibility of β-Lactamase- and Plasmid-Mediated Quinolone Resistance (PMQR)-Producing Strains
3. Discussion
4. Materials and Methods
4.1. Sample Characterization
4.2. Bacterial Isolation and Identification
4.3. Statistical Analyses of Results
4.4. Molecular Detection of Resistance Genes
4.5. Antibiotic Susceptibility Testing of Strains with Resistance Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaspar, P.; Pires, I.; Magalhães, A.; Lda, C. Boas Práticas em Cultivo de Ostra—Algarve; APA/ARH do Algarve: Amadora, Portugal, 2017; ISBN 978-972-8577-70-4. [Google Scholar]
- Ottinger, M.; Clauss, K.; Kuenzer, C. Aquaculture: Relevance, distribution, impacts and spatial assessments—A review. Ocean Coast. Manag. 2016, 119, 244–266. [Google Scholar] [CrossRef]
- Elbashir, S.; Parveen, S.; Schwarz, J.; Rippen, T.; Jahncke, M.; DePaola, A. Seafood pathogens and information on antimicrobial resistance: A review. Food Microbiol. 2018, 70, 85–93. [Google Scholar] [CrossRef]
- Direção-Geral de Recursos Naturais Segurança e Serviços Marítimos. Plano Estratégico Para a Aquicultura Portuguesa 2014–2020; Direção-Geral de Recursos Naturais Segurança e Serviços Marítimos: Lisbon, Portugal, 2013. [Google Scholar]
- Araújo, J.; Soares, F.; Pousão-Ferreira, P. Offshore Production of Mediterranean Mussels in Southern Portugal. World Aquac. 2018, 49, 55–57. Available online: https://www.was.org/Magazine/Vol/49/2#.X5VTYIj7RPY (accessed on 25 October 2020).
- Goulletquer, P. Cultured Aquatic Species Information Programme. Mytilus edulis. Available online: http://www.fao.org/fishery/culturedspecies/Mytilus_edulis/en (accessed on 13 August 2020).
- Chiesa, L.M.; Nobile, M.; Malandra, R.; Panseri, S.; Arioli, F. Occurrence of antibiotics in mussels and clams from various FAO areas. Food Chem. 2018, 240, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueras, A. Cultured Aquatic Species Information Programme. Ruditapes decussatus. Available online: http://www.fao.org/fishery/culturedspecies/Ruditapes_decussatus/en (accessed on 13 August 2020).
- Zannella, C.; Mosca, F.; Mariani, F.; Franci, G.; Folliero, V.; Galdiero, M.; Tiscar, P.G.; Galdiero, M. Microbial Diseases of Bivalve Mollusks: Infections, Immunology and Antimicrobial Defense. Mar. Drugs 2017, 15, 182. [Google Scholar] [CrossRef]
- Fernández, N.T.; Mazón-Suástegui, J.M.; Vázquez-Juárez, R.; Ascencio-Valle, F.; Romero, J. Changes in the composition and diversity of the bacterial microbiota associated with oysters (Crassostrea corteziensis, Crassostrea gigas and Crassostrea sikamea) during commercial production. FEMS Microbiol. Ecol. 2014, 88, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Pierce, M.L.; Ward, J.E. Gut Microbiomes of the Eastern Oyster (Crassostrea virginica) and the Blue Mussel (Mytilus edulis): Temporal Variation and the Influence of Marine Aggregate-Associated Microbial Communities. mSphere 2019, 4, e00730-19. [Google Scholar] [CrossRef] [Green Version]
- Tseng, S.Y.; Liu, P.Y.; Lee, Y.H.; Wu, Z.Y.; Huang, C.C.; Cheng, C.C.; Tung, K.C. The pathogenicity of Shewanella algae and ability to tolerate a wide range of temperatures and salinities. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 6976897. [Google Scholar] [CrossRef] [Green Version]
- Grevskott, D.H.; Svanevik, C.S.; Sunde, M.; Wester, A.L.; Lunestad, B.T. Marine bivalve mollusks as possible indicators of multidrug-resistant Escherichia coli and other species of the Enterobacteriaceae family. Front. Microbiol. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopatek, M.; Wieczorek, K.; Osek, J. Prevalence and antimicrobial resistance of Vibrio parahaemolyticus isolated from raw shellfish in Poland. J. Food Prot. 2015, 78, 1029–1033. [Google Scholar] [CrossRef]
- O’Neill, J. Review on Antimicrobial Resistance. In Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations; London, UK, 2014; Available online: https://amr-review.org/Publications.html (accessed on 12 April 2021).
- Furushita, M.; Shiba, T.; Maeda, T.; Yahata, M.; Kaneoka, A.; Takahashi, Y.; Torii, K.; Hasegawa, T.; Ohta, M. Similarity of tetracycline resistance genes isolated from fish farm bacteria to those from clinical isolates. Appl. Environ. Microbiol. 2003, 69, 5336–5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Diversity of tetracycline resistance genes in bacteria from aquaculture sources in Australia. J. Appl. Microbiol. 2007, 103, 2016–2025. [Google Scholar] [CrossRef]
- Rivas, A.J.; Lemos, M.L.; Osorio, C.R. Photobacterium damselae subsp. Damselae, a bacterium pathogenic for marine animals and humans. Front. Microbiol. 2013, 4, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froelich, B.A.; Noble, R.T. Vibrio bacteria in raw oysters: Managing risks to human health. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, A.; Abdullah, S. Bacillus cereus food poisoning: International and Indian perspective. J. Food Sci. Technol. 2015, 52, 2500–2511. [Google Scholar] [CrossRef] [Green Version]
- Lasa, A.; Mira, A.; Camelo-Castillo, A.; Belda-Ferre, P.; Romalde, J.L. Characterization of the microbiota associated to Pecten maximus gonads using 454-pyrosequencing. Int. Microbiol. 2016, 19, 93–99. [Google Scholar] [CrossRef]
- Kacar, A. Some microbial characteristics of mussels (Mytilus galloprovincialis) in coastal city area. Environ. Sci. Pollut. Res. 2011, 18, 1384–1389. [Google Scholar] [CrossRef]
- Radó, J.; Kaszab, E.; Benedek, T.; Kriszt, B.; Szoboszlay, S. First isolation of carbapenem-resistant Acinetobacter beijerinckii from an environmental sample. Acta Microbiol. Immunol. Hung. 2019, 66, 113–130. [Google Scholar] [CrossRef]
- Richards, G.P.; Watson, M.A.; Crane, E.J.; Burt, I.G.; Bushek, D. Shewanella and Photobacterium spp. in oysters and seawater from the Delaware bay. Appl. Environ. Microbiol. 2008, 74, 3323–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgueiro, V.; Manageiro, V.; Bandarra, N.M.; Reis, L.; Caniça, M. Bacterial Diversity and Antibiotic Susceptibility of Sparus aurata from Aquaculture. Microorganisms 2020, 8, 1343. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Robles, M.F.; Álvarez-Contreras, A.K.; Juárez-García, P.; Natividad-Bonifacio, I.; Curiel-Quesada, E.; Vázquez-Salinas, C.; Quiñones-Ramírez, E.I. Virulence factors and antimicrobial resistance in environmental strains of Vibrio alginolyticus. Int. Microbiol. 2016, 19, 191–198. [Google Scholar] [CrossRef]
- Khan, A.; Aung, T.T.; Chaudhuri, D. The First Case of Native Mitral Valve Endocarditis due to Micrococcus luteus and Review of the Literature. Case Rep. Cardiol. 2019, 2019, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.W.; Hong, Y.J.; Jo, J.I.; Ha, S.D.; Kim, S.H.; Lee, H.J.; Rhee, M.S. Raw ready-to-eat seafood safety: Microbiological quality of the various seafood species available in fishery, hyper and online markets. Lett. Appl. Microbiol. 2017, 64, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bozcal, E.; Dagdeviren, M. Bacterial metagenome analysis of Mytilus galloprovincialis collected from Istanbul and Izmir coastal stations of Turkey. Environ. Monit. Assess. 2020, 192, 186. [Google Scholar] [CrossRef]
- Fernández-Delgado, M.; Contreras, M.; García-Amado, M.A.; Gueneau, P.; Suárez, P. Occurrence of Proteus mirabilis associated with two species of Venezuelan oysters. Rev. Inst. Med. Trop. Sao Paulo 2007, 49, 355–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacramento, A.G.; Fernandes, M.R.; Sellera, F.P.; Dolabella, S.S.; Zanella, R.C.; Cerdeira, L.; Lincopan, N. VanA-type vancomycin-resistant Enterococcus faecium ST1336 isolated from mussels in an anthropogenically impacted ecosystem. Mar. Pollut. Bull. 2019, 142, 533–536. [Google Scholar] [CrossRef]
- Roslev, P.; Iversen, L.; Sønderbo, H.L.; Iversen, N.; Bastholm, S. Uptake and persistence of human associated Enterococcus in the mussel Mytilus edulis: Relevance for faecal pollution source tracking. J. Appl. Microbiol. 2009, 107, 944–953. [Google Scholar] [CrossRef]
- Chapman, C. Investigation into the Microbiological Causes of Epizootics of Pacific Oyster Larvae (Crassostrea gigas) in Commercial Production. Ph.D. Thesis, University of Tasmania, Hobart, Australia, 2012. [Google Scholar]
- Drzewiecka, D. Significance and Roles of Proteus spp. Bacteria in Natural Environments. Microb. Ecol. 2016, 72, 741–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa, O.V.; Dos Fernandes Vieira, R.H.S.; De Menezes, F.G.R.; Dos Reis, C.M.F.; Hofer, E. Detection of Vibrio parahaemolyticus and Vibrio cholerae in oyster, Crassostrea rhizophorae, collected from a natural nursery in the Cocó river estuary, Fortaleza, Ceará, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2004, 46, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Baldi, F.; Gallo, M.; Marchetto, D.; Faleri, C.; Maida, I.; Fani, R. Manila clams from Hg polluted sediments of Marano and Grado lagoons (Italy) harbor detoxifying Hg resistant bacteria in soft tissues. Environ. Res. 2013, 125, 188–196. [Google Scholar] [CrossRef]
- Santoiemma, P.P.; Kalainov, D.M.; Mehta, M.P.; Bolon, M.K. An unusual case of Staphylococcus pasteuri osteomyelitis. Infect. Dis. Rep. 2020, 12, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, K.P.; Pai, P.G. A rare infective endocarditis caused by Vagococcus fluvialis. J. Cardiol. Cases 2019, 20, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Mori, N.; Kawai, F.; Sakurai, A.; Toyoda, M.; Mikami, Y.; Uehara, Y.; Furukawa, K. Vagococcus fluvialis as a causative pathogen of bloodstream and decubitus ulcer infection: Case report and systematic review of the literature. J. Infect. Chemother. 2020, 6. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, J.; Hu, Q.; Rao, X. Morganella morganii, a non-negligent opportunistic pathogen. Int. J. Infect. Dis. 2016, 50, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Sharma, P.; Soni, P. First case report of Providencia Rettgeri neonatal sepsis. BMC Res. Notes 2017, 10, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Kanuparthy, A.; Challa, T.; Meegada, S.; Siddamreddy, S.; Muppidi, V. Staphylococcus warneri: Skin Commensal and a Rare Cause of Urinary Tract Infection. Cureus 2020, 12, e8337. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Xu, C.G.; Yang, Y.B.; Xing, X.X.; Liu, X.; Qu, Q.W.; Ding, W.Y.; Bello-Onaghise, G.; Li, Y.H. Histidine metabolism and IGPD play a key role in cefquinome inhibiting biofilm formation of Staphylococcus xylosus. Front. Microbiol. 2018, 9, 665. [Google Scholar] [CrossRef] [Green Version]
- Walczak, N.; Puk, K.; Guz, L. Bacterial flora associated with diseased freshwater ornamental fish. J. Vet. Res. 2017, 61, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Al-Kharousi, Z.S.; Guizani, N.; Al-Sadi, A.M.; Al-Bulushi, I.M.; Shaharoona, B. Hiding in Fresh Fruits and Vegetables: Opportunistic Pathogens May Cross Geographical Barriers. Int. J. Microbiol. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Herridge, W.P.; Shibu, P.; O’Shea, J.; Brook, T.C.; Hoyles, L. Bacteriophages of Klebsiella spp., their diversity and potential therapeutic uses. J. Med. Microbiol. 2020, 69, 176–194. [Google Scholar] [CrossRef]
- Pérez-Vazquez, M.; Oteo-Iglesias, J.; Sola-Campoy, P.J.; Carrizo-Manzoni, H.; Bautista, V.; Lara, N.; Aracil, B.; Alhambra, A.; Martínez-Martínez, L.; Campos, J.; et al. Characterization of Carbapenemase-Producing Klebsiella oxytoca in Spain, 2016–2017. Antimicrob. Agents Chemother. 2019, 63, e02529-18. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Li, H.; Feng, J.; Li, Y.; Chen, X.; Guo, X.; Chen, W.; Wang, L.; Lin, L.; Yang, H.; et al. NDM-1 encoded by a pNDM-BJ01-like plasmid p3SP-NDM in clinical Enterobacter aerogenes. Front. Microbiol. 2015, 6, 294. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Ben Omar, N.; Molinos, A.C.; López, R.L.; Grande, M.J.; Martínez-viedma, P.; Ortega, E.; Cañamero, M.M.; Galvez, A. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol. 2008, 123, 38–49. [Google Scholar] [CrossRef]
- Kubota, H.; Mitani, A.; Niwano, Y.; Takeuchi, K.; Tanaka, A.; Yamaguchi, N.; Kawamura, Y.; Hitomi, J. Moraxella species are primarily responsible for generating malodor in laundry. Appl. Environ. Microbiol. 2012, 78, 3317–3324. [Google Scholar] [CrossRef] [Green Version]
- Alkhatib, N.J.; Younis, M.H.; Alobaidi, A.S.; Shaath, N.M. An unusual osteomyelitis caused by Moraxella osloensis: A case report. Int. J. Surg. Case Rep. 2017, 41, 146–149. [Google Scholar] [CrossRef]
- Kaeuffer, C.; Schramm, F.; Meyer, A.; Hansmann, Y.; Guffroy, A.; Argemi, X. First case of Comamonas aquatica bacteremia complicated by septic shock. Med. Mal. Infect. 2018, 48, 540–542. [Google Scholar] [CrossRef]
- Parthasarathy, A.; Gan, H.M.; Wong, N.H.; Savka, M.A.; Steiner, K.K.; Henry, K.R.; Hudson, A.O. Isolation and genomic characterization of six endophytic bacteria isolated from Saccharum sp (sugarcane): Insights into antibiotic, secondary metabolite and quorum sensing metabolism. J. Genom. 2018, 6, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Gaastra, W.; Kusters, J.G.; van Duijkeren, E.; Lipman, L.J.A. Escherichia fergusonii . Vet. Microbiol. 2014, 172, 7–12. [Google Scholar] [CrossRef]
- Hajjar, R.; Ambaraghassi, G.; Sebajang, H.; Schwenter, F.; Su, S.H. Raoultella ornithinolytica: Emergence and resistance. Infect. Drug Resist. 2020, 13, 1091–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rösch, R.M.; Buschmann, K.; Brendel, L.; Schwanz, T.; Vahl, C.-F. Lactococcus garvieae Endocarditis in a Prosthetic Aortic Valve: A Case Report and Literature Review. J. Investig. Med. High. Impact Case Rep. 2019, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Nagaya, Y.; Pradel, E.; Ooka, T.; Ogura, Y.; Katsura, K.; Kurokawa, K.; Oshima, K.; Hattori, M.; Parkhill, J.; et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol. Evol. 2014, 6, 2096–2110. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.D.; Chun, C.; Hong, K.S. Hemolytic uremic syndrome caused by Escherichia fergusonii infection. Kidney Res. Clin. Pract. 2019, 38, 253–255. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.X.; Wang, A.; Wang, J.Y. Antibiotic resistance monitoring in heterotrophic bacteria from anthropogenic-polluted seawater and the intestines of oyster Crassostrea hongkongensis. Ecotoxicol. Environ. Saf. 2014, 109, 27–31. [Google Scholar] [CrossRef]
- Catry, B.; Cavaleri, M.; Baptiste, K.; Grave, K.; Grein, K.; Holm, A.; Jukes, H.; Liebana, E.; Navas, A.L.; Mackay, D.; et al. Use of colistin-containing products within the European Union and European Economic Area (EU/EEA): Development of resistance in animals and possible impact on human and animal health. Int. J. Antimicrob. Agents 2015, 46, 297–306. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, R.; Schwarz, S.; Wu, C.; Shen, J.; Walsh, T.R.; Wang, Y. Farm animals and aquaculture: Significant reservoirs of mobile colistin resistance genes. Environ. Microbiol. 2020, 22, 2469–2484. [Google Scholar] [CrossRef] [Green Version]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Dahanayake, P.S.; Hossain, S.; Wickramanayake, M.V.K.S.; Wimalasena, S.H.M.P.; Heo, G.-J. Manila clam (Ruditapes philippinarum) marketed in Korea as a source of vibrios harbouring virulence and β-lactam resistance genes. Lett. Appl. Microbiol. 2020, 71, 46–53. [Google Scholar] [CrossRef]
- Jones-dias, D.; Manageiro, V.; Francisco, A.P.; Martins, A.P.; Domingues, G.; Louro, D.; Ferreira, E.; Caniça, M. Assessing the molecular basis of transferable quinolone resistance in Escherichia coli and Salmonella spp. from food-producing animals and food products. Vet. Microbiol. 2013, 167, 523–531. [Google Scholar] [CrossRef]
- Su, H.; Hu, X.; Wang, L.; Xu, W.; Xu, Y.; Wen, G.; Li, Z.; Cao, Y. Contamination of antibiotic resistance genes (ARGs) in a typical marine aquaculture farm: Source tracking of ARGs in reared aquatic organisms. J. Environ. Sci. Heal. Part. B. 2020, 55, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Campos, J.; Mourão, J.; Pereira, J.; Novais, C.; Peixe, L. Inflow water is a major source of trout farming contamination with Salmonella and multidrug resistant bacteria. Sci. Total Environ. 2018, 642, 1163–1171. [Google Scholar] [CrossRef]
- De Silva, B.C.J.; Hossain, S.; Dahanayake, P.S.; Heo, G.-J. Aeromonas spp. from marketed Yesso scallop (Patinopecten yessoensis): Molecular characterization, phylogenetic analysis, virulence properties and antimicrobial susceptibility. J. Appl. Microbiol. 2019, 126, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Giedraitiene, A.; Vitkauskiene, A.; Naginiene, R.; Pavilonis, A. Antibiotic resistance mechanisms of clinically important bacteria. Medicina (B. Aires) 2011, 47, 137–146. [Google Scholar] [CrossRef]
- Cabello, F.C.; Godfrey, H.P.; Buschmann, A.H.; Dölz, H.J. Aquaculture as yet another environmental gateway to the development and globalisation of antimicrobial resistance. Lancet Infect. Dis. 2016, 16, e127–e133. [Google Scholar] [CrossRef]
- Jones-Dias, D.; Manageiro, V.; Caniça, M. Influence of agricultural practice on mobile bla genes: IncI1-bearing CTX-M, SHV, CMY and TEM in Escherichia coli from intensive farming soils. Environ. Microbiol. 2016, 18, 260–272. [Google Scholar] [CrossRef]
- Dean, A.G.; Sullivan, K.M.; Soe, M.M. OpenEpi: Open Source Epidemiologic Statistics for Public Health. Available online: www.OpenEpi.com (accessed on 29 July 2020).
- McAuliffe, G.N.; Hennessy, J.; Baird, R.W. Relative frequency, characteristics, and antimicrobial susceptibility patterns of Vibrio spp., Aeromonas spp., Chromobacterium violaceum, and Shewanella spp. in the Northern Territory of Australia, 2000–2013. Am. J. Trop. Med. Hyg. 2015, 92, 605–610. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Family | Bacterial Species | Bivalve Species |
|---|---|---|
| Aeromonadaceae | Aeromonas punctata | R. decussatus |
| Aeromonas sp. | Mytilus spp. | |
| Bacillaceae | Bacillus sp. | R. decussatus |
| Bacillus cereus group | C. gigas, Mytilus spp. and R. decussatus | |
| Comamonadaceae | Comamonas aquatica | R. decussatus |
| Enterobacteriaceae | Citrobacter werkmanii | |
| Pseudocitrobacter faecalis | ||
| Enterobacter cancerogenus | Mytilus spp. | |
| Escherichia fergusonii | C. gigas | |
| Raoultella ornithinolytica | Mytilus spp. and R. decussatus | |
| Citrobacter braakii | C. gigas and R. decussatus | |
| Klebsiella aerogenes | ||
| Enterobacter spp. (E. hormaechei, E. kobei) | C. gigas and Mytilus spp. | |
| Klebsiella spp. (K. pneumoniae, K. oxytoca) | ||
| Citrobacter freundii | C. gigas, Mytilus spp. and R. decussatus | |
| Enterobacter cloacae | ||
| Escherichia coli | ||
| Enterococcaceae | Enterococcus spp. (E. faecalis, E. hirae) | |
| Enterococcus faecium | C. gigas | |
| Vagococcus fluvialis | Mytilus spp. | |
| Micrococcaceae | Micrococcus luteus | |
| Moraxellaceae | Acinetobacter spp. (A. beijerinckii, A. junii, A. pittii, A. ursingii) | C. gigas |
| Moraxella osloensis | ||
| Morganellaceae | Morganella morganii | C. gigas and R. decussatus |
| Proteus hauseri | Mytilus spp. | |
| Proteus vulgaris | C. gigas, Mytilus spp. and R. decussatus | |
| Providencia spp. (P. rettgeri, P. stuartii) | ||
| Pseudomonadaceae | Pseudomonas mendocina | Mytilus spp. and R. decussatus |
| Pseudomonas putida | C. gigas and Mytilus spp. | |
| Shewanellaceae | Shewanella algae | |
| Staphylococcaceae | Staphylococcus pasteuri | Mytilus spp. and R. decussatus |
| Staphylococcus warneri | Mytilus spp. | |
| Staphylococcus xylosus | R. decussatus | |
| Streptococcaceae | Lactococcus garvieae | |
| Vibrionaceae | Photobacterium damselae | C. gigas and Mytilus spp. |
| Vibrio alginolyticus | C. gigas, Mytilus spp. and R. decussatus | |
| Vibrio fluvialis | Mytilus spp. | |
| Vibrio spp. (V. furnissii, V. vulnificus) | R. decussatus | |
| Yersiniaceae | Serratia marcescens | C. gigas |
| Bivalve’s Common Name | Bivalve Species | Bacterial Family | Season | Collection Site | Antibiotic | OR 1 | 95% CI | p Value |
|---|---|---|---|---|---|---|---|---|
| Clams | R. decussatus/All | Morganellaceae | Summer | All | NA | 6.933 | 1.02–54.16 | 0.02 |
| Mussels | Mytilus spp./All | Enterobacteriaceae | Summer | All | NA | 0.1908 (P) | 0.05897–0.597 | ≤0.01 |
| Mytilus spp./All | Enterobacteriaceae | Autumn | All | NA | 5.242 | 1.675–16.96 | ≤0.01 | |
| Mytilus spp. | Enterobacteriaceae | Summer | All | NA | 0.1689 (P) | 0.0584–0.4595 | ≤0.01 | |
| Mytilus spp. | Enterobacteriaceae | Autumn | All | NA | 5.92 | 2.176–17.12 | ≤0.01 | |
| Mytilus spp. | Morganellaceae | All | All | NA | 0.437 (P) | 0.1842–0.983 | 0.02 | |
| Mytilus spp. | Shewanellaceae | Summer | All | NA | 10.76 | 1.202–503 | ≤0.01 | |
| Mytilus spp. | Vibrionaceae | Summer | All | NA | 3.54 | 1.058–12.75 | 0.02 | |
| Japanese oysters | C. gigas/All | Enterobacteriaceae | Summer | All | NA | 9.429 | 2.263–45.46 | ≤0.01 |
| C. gigas | Morganellaceae | Summer | All | NA | 0.0692 (P) | 0.001588–0.5202 | ≤0.01 | |
| C. gigas | Morganellaceae | Autumn | All | NA | 14.45 | 1.922–629.7 | ≤0.01 | |
| C. gigas | All | All | All | Oxytetracycline | 0.4167 (P) | 0.2293–0.7527 | ≤0.01 | |
| C. gigas | All | Summer | All | Oxytetracycline | 2.786 | 1.061–7.35 | 0.02 |
| Bacterial Species | Farm (No. of Strains) | Bivalve Mollusk Species | Season | Decreased Susceptibility Profile | AR Genes |
|---|---|---|---|---|---|
| C. braakii | 1 (n = 1) | R. decussatus | A | AMX, AMC, FOX, CHL, FLO, OTC | qnrB-type1 |
| C. freundii | 1 (n = 1) | C. gigas | A | AMC, CAZ, FOX, FLO, OTC | qnrB44 |
| E. coli | 4 (n = 8) | Mytilus spp. | A | (AMX), AMC, (CAZ), (CIP), CHL, FLO, (FMQ), OTC, SXT | qnrB19,blaTEM-1 |
| R. ornithinolytica | 4 (n = 3) | Mytilus spp. | A | (CIP), FMQ, NAL, OTC | oqxAB |
| S. algae | 2 (n = 3) | Mytilus spp. | S | (AMX), (FMQ), OTC | qnrA3 |
| 2 (n = 1) | Mytilus spp. | S | CIP, FMQ, OTC | qnrA11 | |
| 5 (n = 1) | C. gigas | S | COL | qnrA12 | |
| 3 (n = 1) | Mytilus spp. | A | FMQ, OTC | qnrA2 |
| Bacterial Family | Method | Antibiotics Tested (Concentration) | Breakpoints |
|---|---|---|---|
| Enterobacteriaceae | Disk diffusion | AMC (20 + 10 µg), AZT (30 µg), FEP (30 µg), CTX (5 µg), FOX (30 µg), CAZ (10 µg), ERT (10 µg), IMP (10 µg), MEM (10 µg), PTZ (36 µg), CIP (5 µg), SXT (25 µg), GEN (10 µg) | EUCAST 1 |
| MIC | CHL, FLO, OTC FMQ CIP | CLSI VET08 2 CASFM VET 2019 3 EUCAST | |
| Shewanellaceae | Disk diffusion | AZT (30 µg), FEP (30 µg), CAZ (10 µg), IMP (10 µg), MEM (10 µg), PTZ (36 µg), CIP (5 µg), LEV (5 µg), AN (30 µg), GEN (10 µg), NET (10 µg), TMN (10 µg) | EUCAST 4 |
| MIC | CHL, FLO, OTC, CIP, FMQ | CLSI M100 4,5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgueiro, V.; Reis, L.; Ferreira, E.; Botelho, M.J.; Manageiro, V.; Caniça, M. Assessing the Bacterial Community Composition of Bivalve Mollusks Collected in Aquaculture Farms and Respective Susceptibility to Antibiotics. Antibiotics 2021, 10, 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091135
Salgueiro V, Reis L, Ferreira E, Botelho MJ, Manageiro V, Caniça M. Assessing the Bacterial Community Composition of Bivalve Mollusks Collected in Aquaculture Farms and Respective Susceptibility to Antibiotics. Antibiotics. 2021; 10(9):1135. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091135
Chicago/Turabian StyleSalgueiro, Vanessa, Lígia Reis, Eugénia Ferreira, Maria João Botelho, Vera Manageiro, and Manuela Caniça. 2021. "Assessing the Bacterial Community Composition of Bivalve Mollusks Collected in Aquaculture Farms and Respective Susceptibility to Antibiotics" Antibiotics 10, no. 9: 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091135