
LyeTx I-b Peptide Attenuates Tumor Burden and Metastasis in a Mouse 4T1 Breast Cancer Model

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. LyeTx I-b Peptide Shows Cytotoxicity against Human and Mouse Breast Cancer Cells
2.2. LyeTxI-b Peptide Reduces the Clonogenic Survival of 4T1 Cells
2.3. LyeTxI-b Increases Sub-Diploid DNA Content and Exposure to Phosphatidylserine
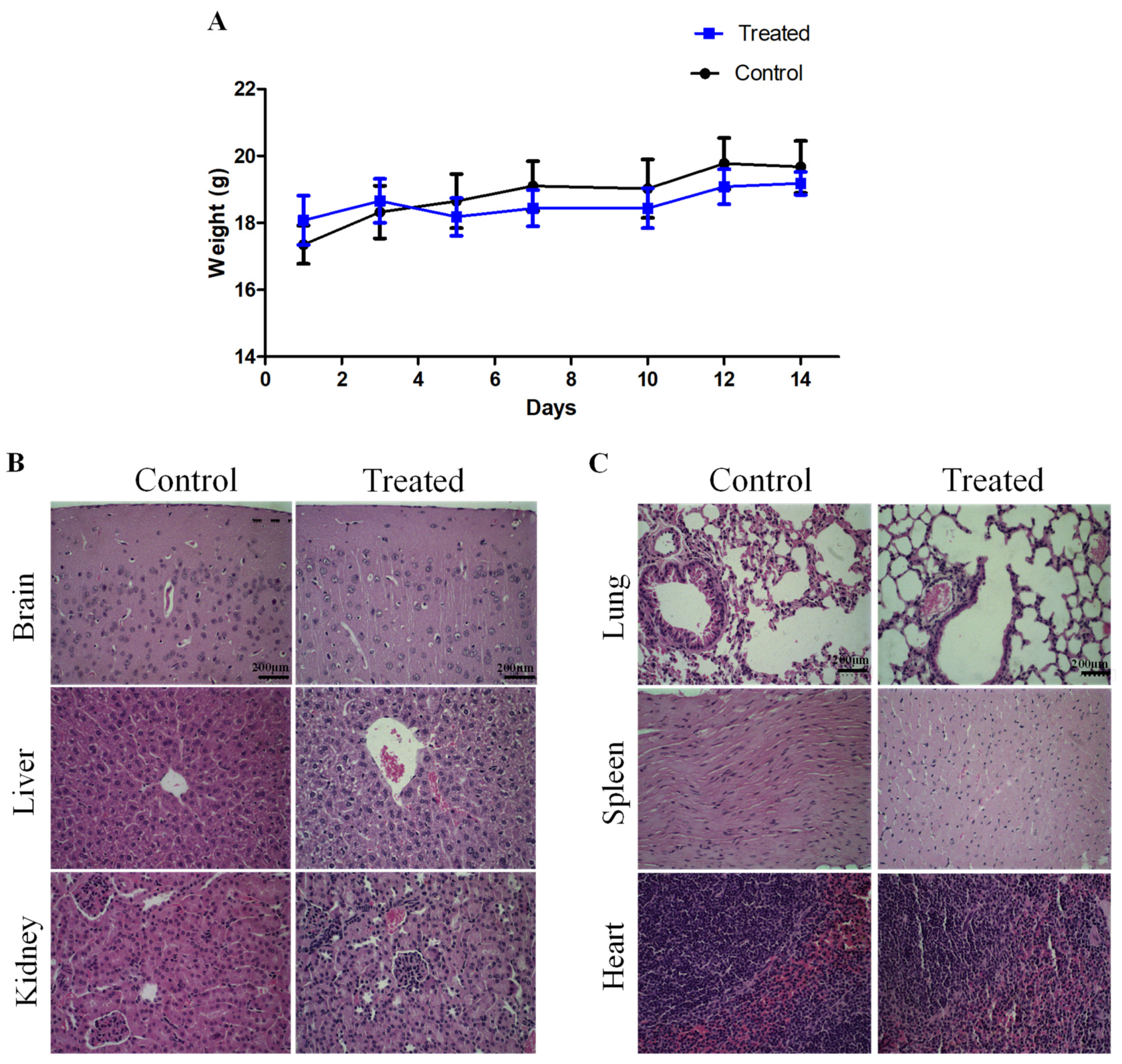
2.4. Sub-Acute Toxicity of LyeTx I-b Peptide
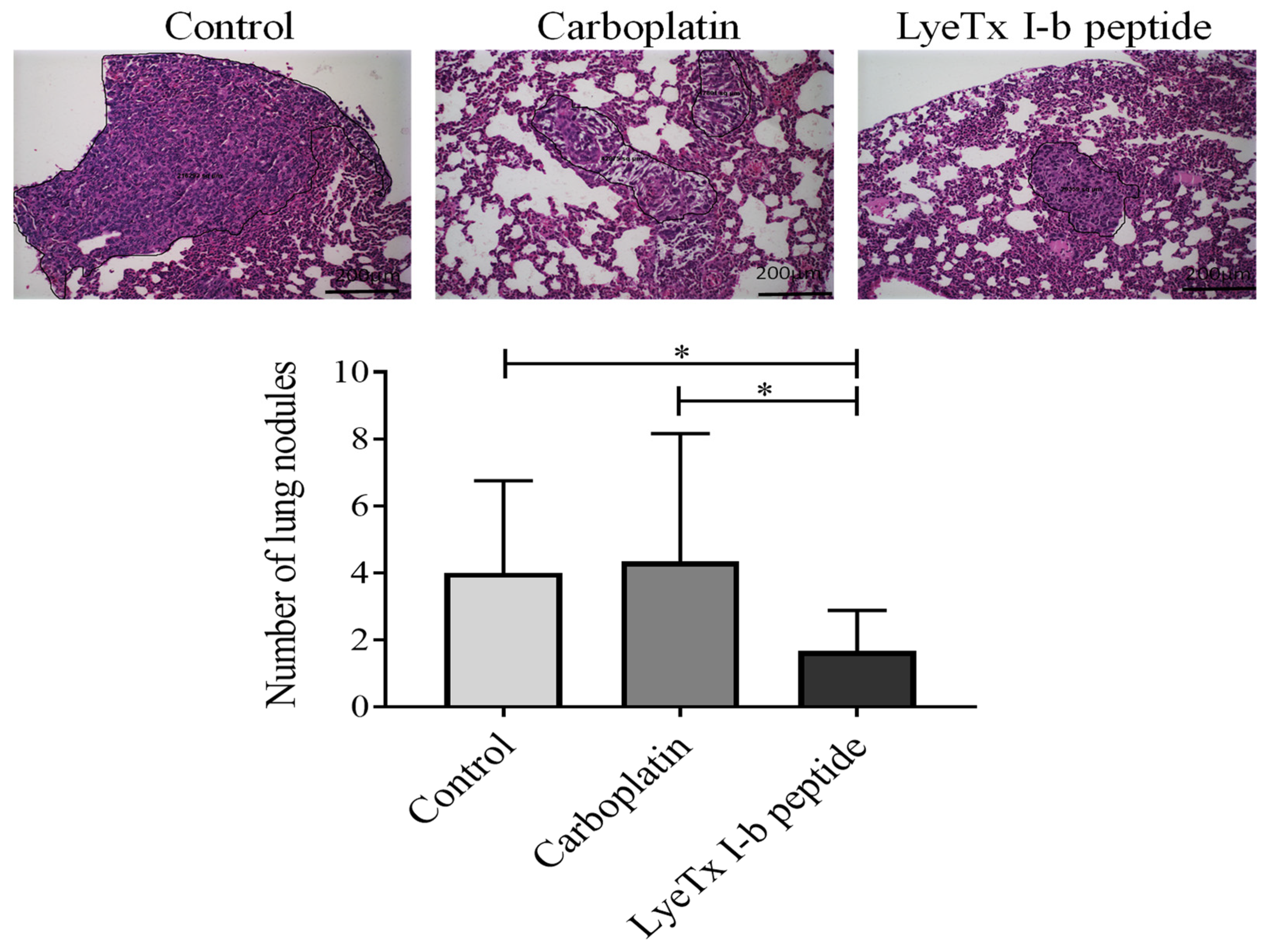
2.5. Intratumoral Administration of LyeTxI-b Decreases Tumor Growth and Lung Metastatic Nodules
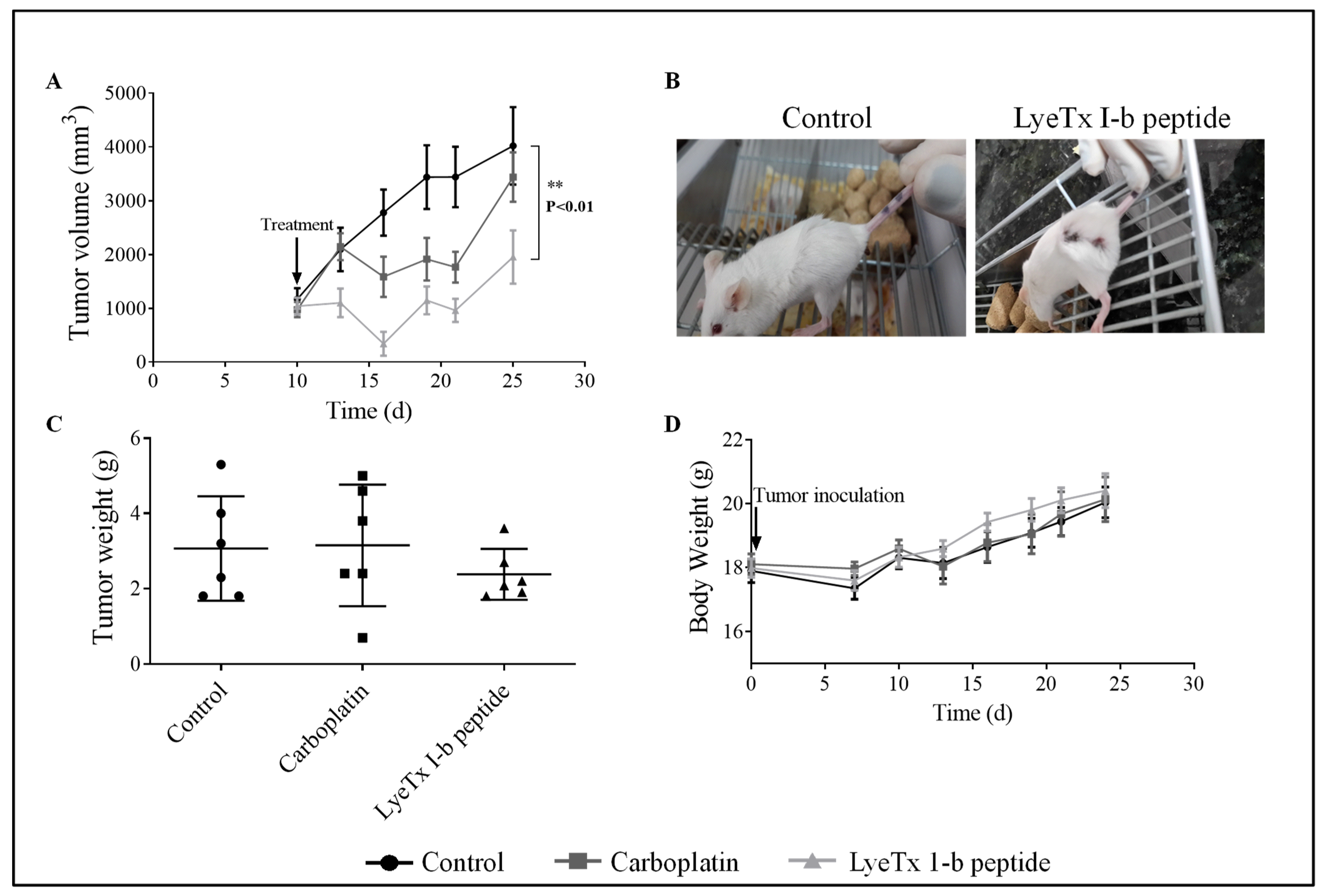
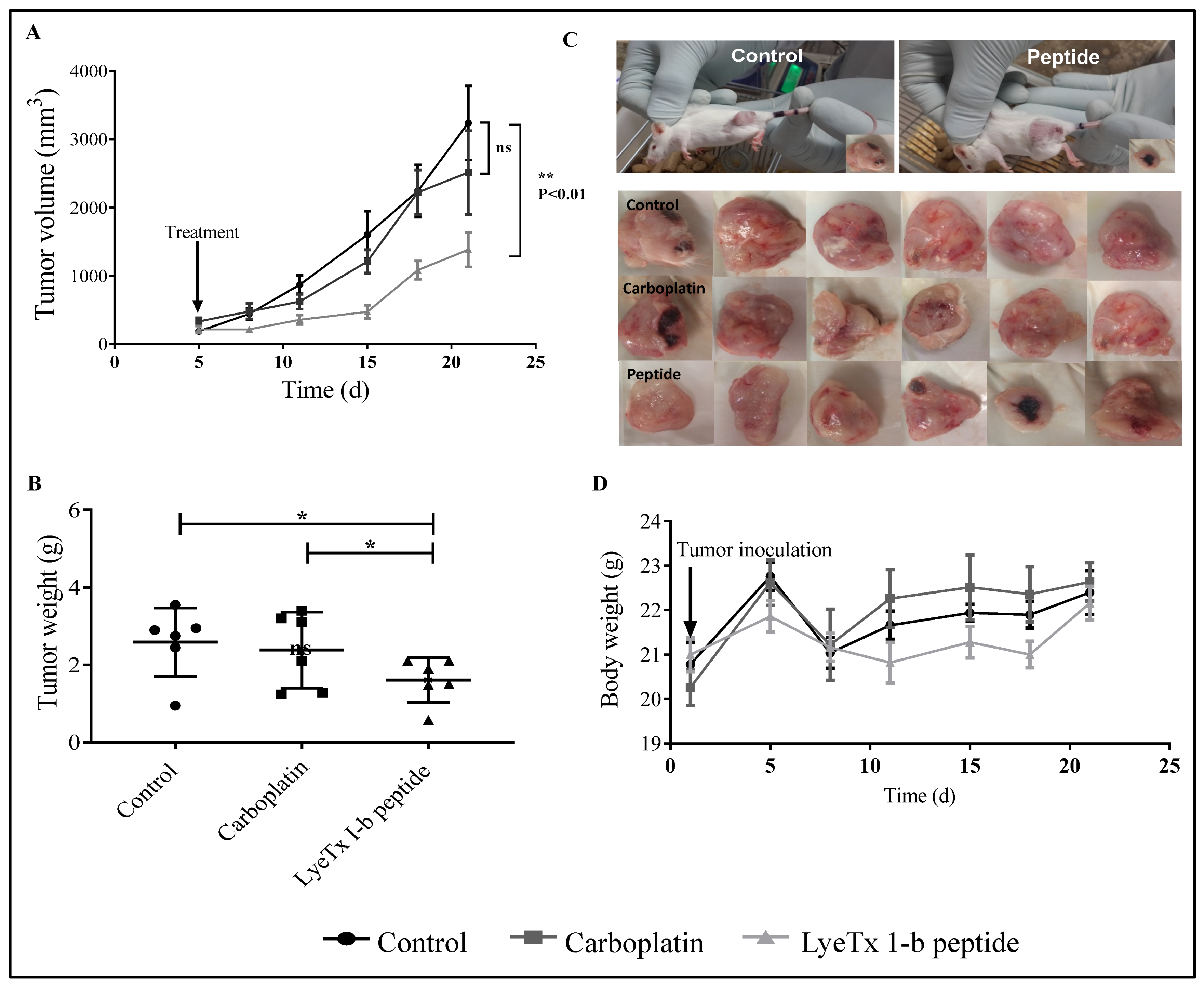
2.6. Systemic Subcutaneous Injection of LyeTx I-b Reduces Tumor Volume
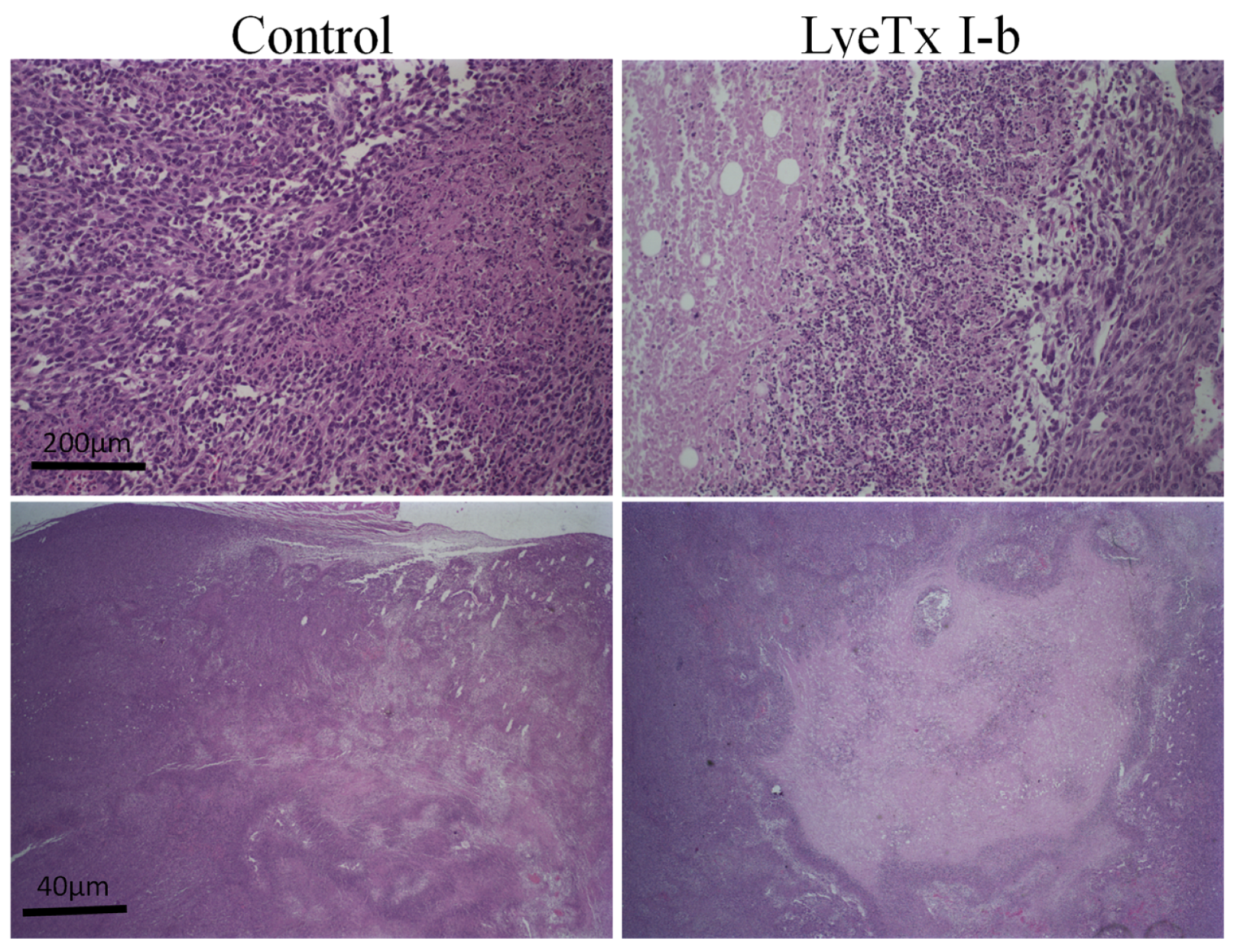
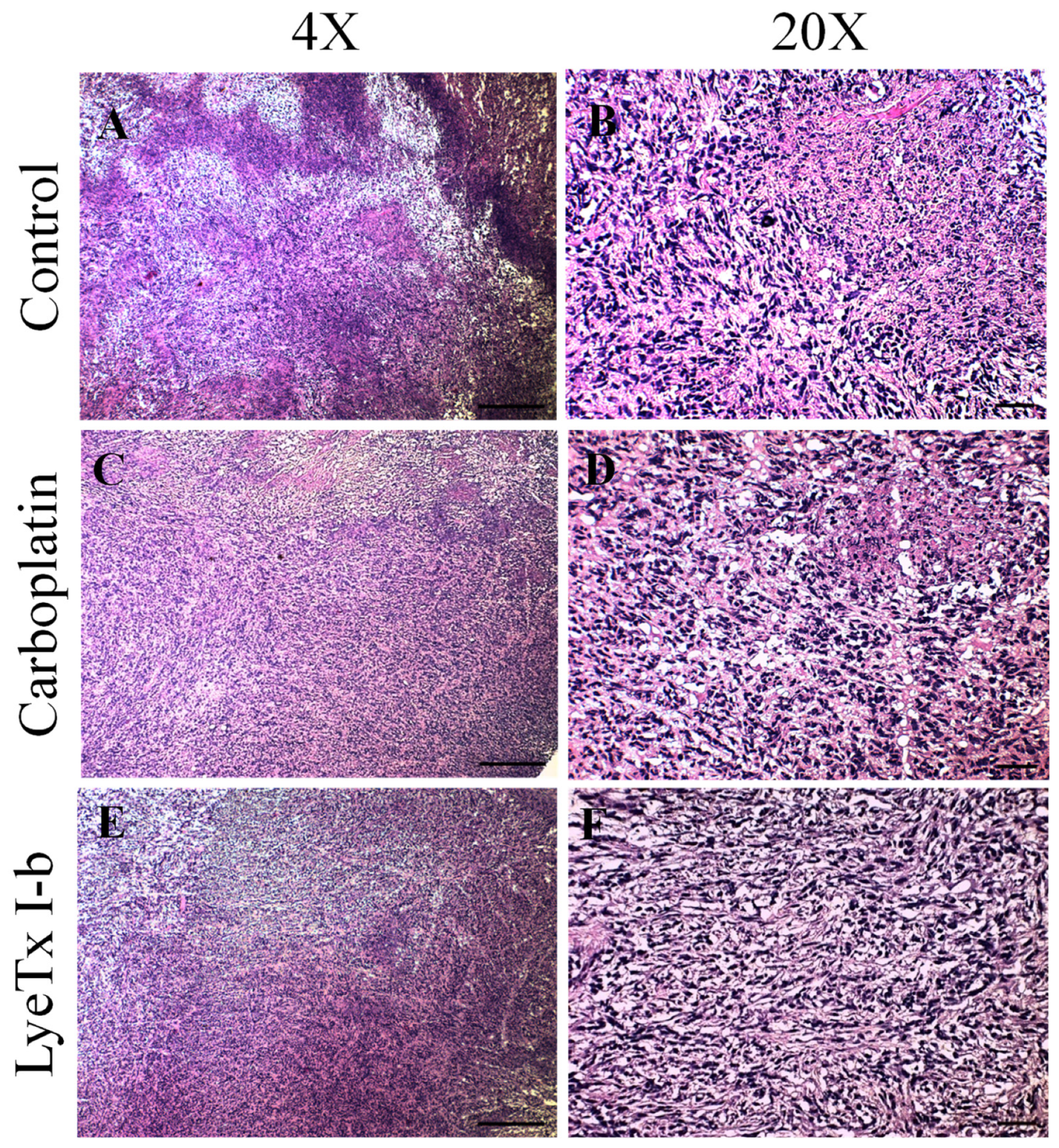
2.7. Systemic Subcutaneous Injection of LyeTx I-b Peptide Reduces Neoplastic Cells in Primary Tumor
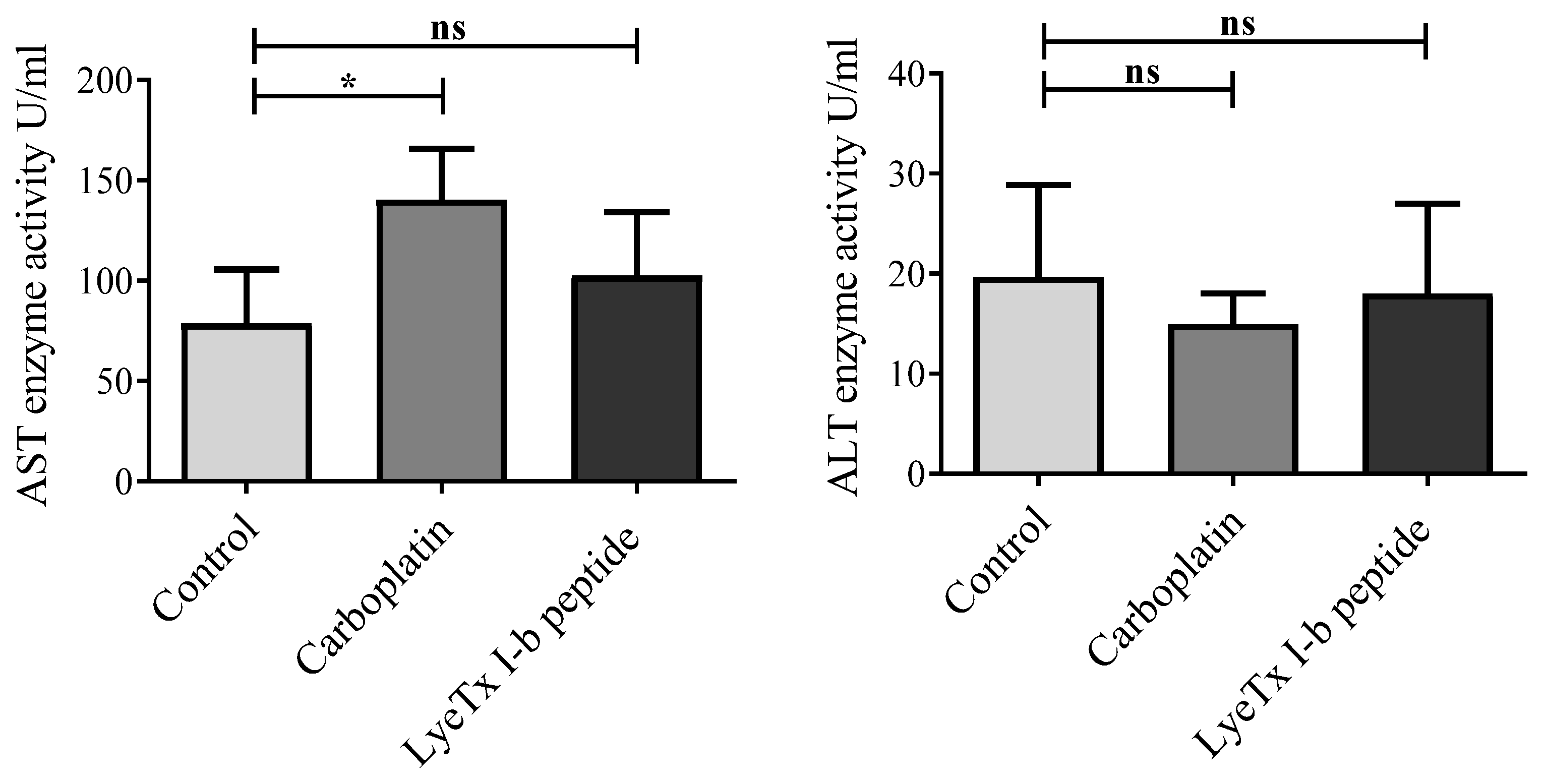
2.8. No Liver Function Alteration Was Induced by LyeTx I-b Treatment
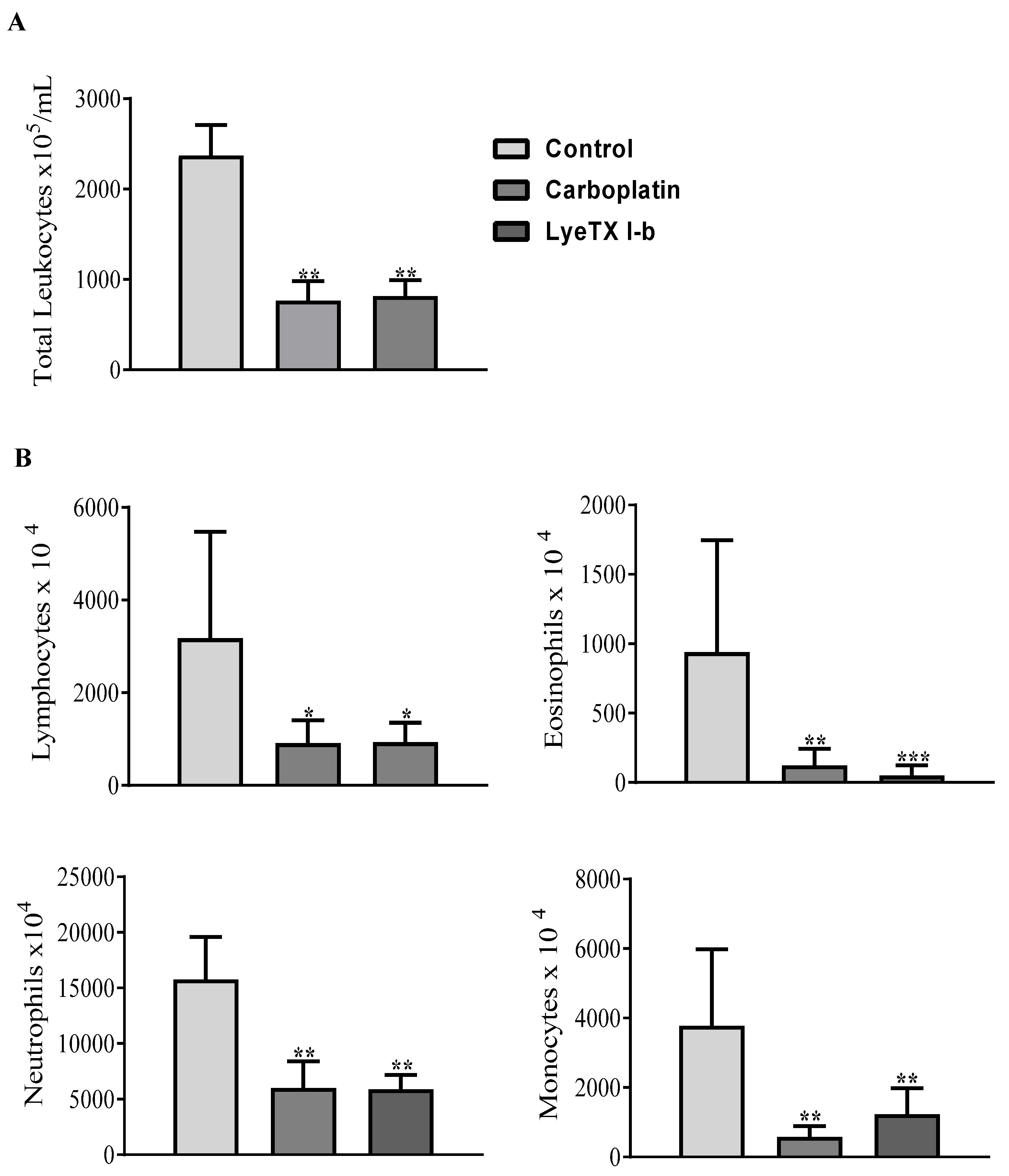
2.9. LyeTx I-b and Carboplatin Reduce the Total and Differential Leukocyte Counts
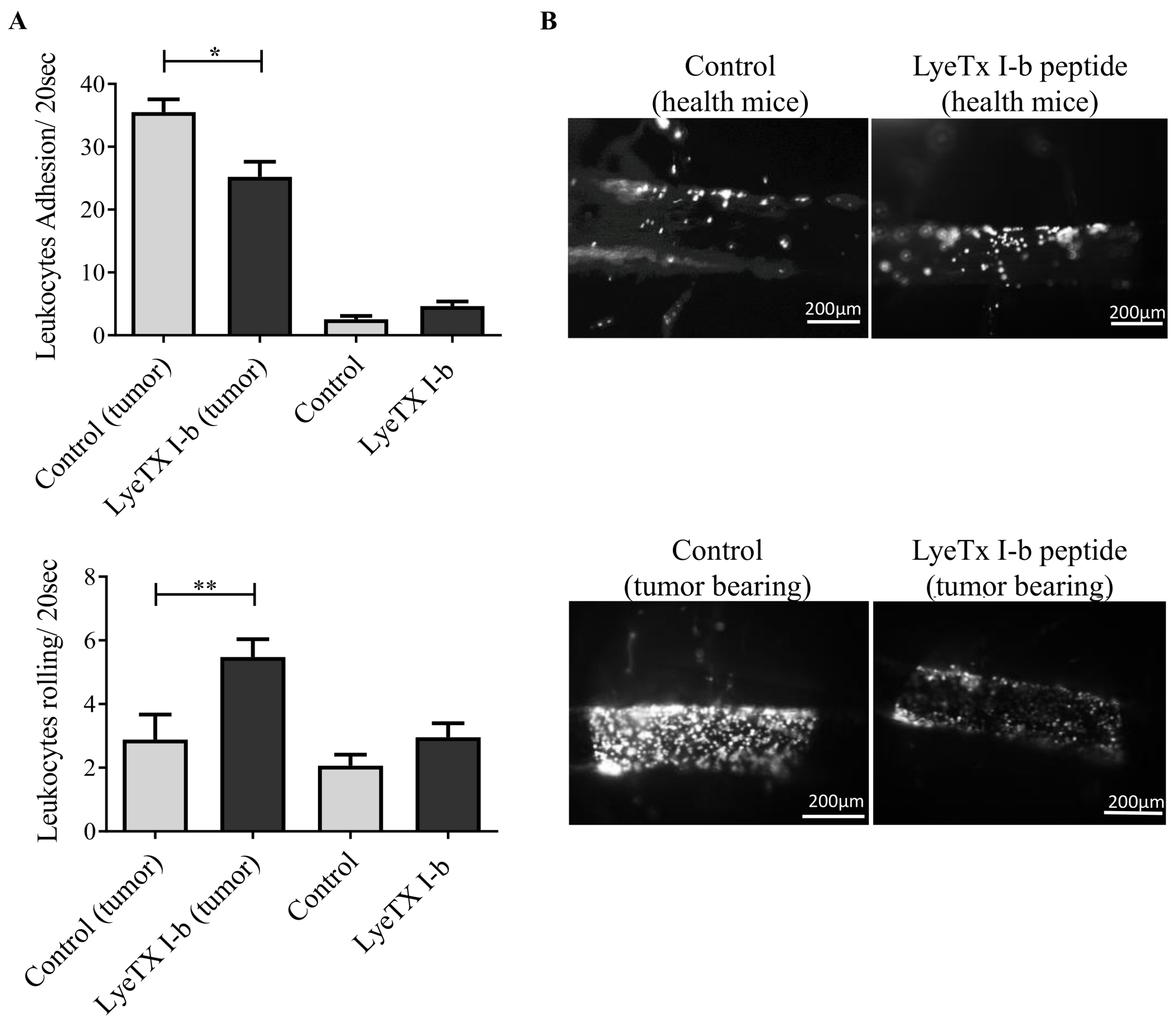
2.10. LyeTx I-b Alters the Numbers of Rolling and Adhering Leukocytes
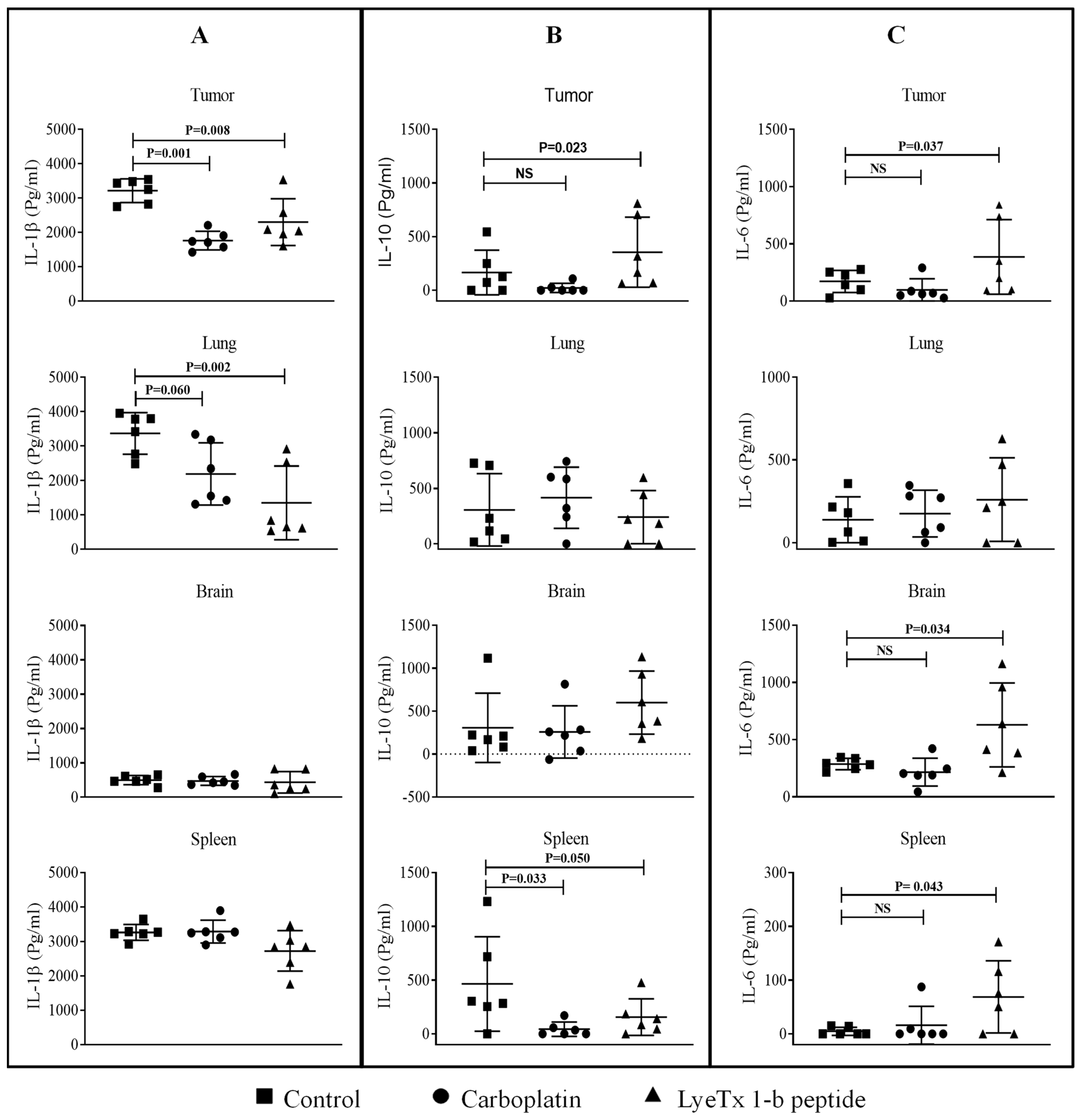
2.11. LyeTx I-b Decreases the Level of Cytokines in the Tumor Microenvironment
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Evaluation of Cell Viability Using the MTT Assay
4.3. Cell Cycle Analysis
4.4. Quantitation of Cell Death Using Annexin V and PI
4.5. Evaluation of the Clonogenic Survival of 4T1 Cells after the Treatment with LyeTx I-b
4.6. Animals and Procedures
4.7. Sub-Acute Toxicity of LyeTx I-b Peptide In Vivo
4.8. Tumor Inoculation and Drug Treatment
4.9. Intratumoral Drug Administration
4.10. Subcutaneous Cells and Drug Administration
4.11. Histological Analysis
4.12. Evaluation of Liver Integrity
4.13. Complete Blood Count
4.14. Manual Quantification of Differential Leukocytes
4.15. Detection and Quantitation of Cytokines and Growth Factors
4.16. Intravital Microscopy
4.17. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, M.; Kelly, E.D.; Abraham, J.; Kruse, M. Invasive lobular breast cancer: A review of pathogenesis, diagnosis, management, and future directions of early stage disease. Semin. Oncol. 2019, 46, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Brane, A.C.; Tollefsbol, T.O. MicroRNAs and Epigenetics Strategies to Reverse Breast Cancer. Cells 2019, 8, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Breast Cancer; World Health Organization: Geneva, Switzerland, 26 March 2021. [Google Scholar]
- Sharma, R. Breast cancer incidence, mortality and mortality-to-incidence ratio (MIR) are associated with human development, 1990–2016: Evidence from Global Burden of Disease Study 2016. Breast Cancer 2019, 26, 428–445. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Bray, F.; Ferlay, J.; Lortet-Tieulent, J.; Anderson, B.O.; Jemal, A. International Variation in Female Breast Cancer Incidence and Mortality Rates. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2015, 24, 1495–1506. [Google Scholar] [CrossRef] [Green Version]
- Holm, J.; Li, J.; Darabi, H.; Eklund, M.; Eriksson, M.; Humphreys, K.; Hall, P.; Czene, K. Associations of Breast Cancer Risk Prediction Tools With Tumor Characteristics and Metastasis. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Thorat, M.A.; Balasubramanian, R. Breast cancer prevention in high-risk women. Best Pract. Res. Clin. Obstet. Gynaecol. 2020, 65, 18–31. [Google Scholar] [CrossRef]
- Hutchinson, L. Breast cancer: Challenges, controversies, breakthroughs. Nat. Rev. Clin. Oncol. 2010, 7, 669–670. [Google Scholar] [CrossRef] [Green Version]
- Hussein, E.A.; Zagho, M.M.; Nasrallah, G.K.; Elzatahry, A.A. Recent advances in functional nanostructures as cancer photothermal therapy. Int. J. Nanomed. 2018, 13, 2897–2906. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Yang, Z.; Lu, W.; Chen, Z.; Chen, L.; Han, S.; Wu, X.; Cai, T.; Cai, Y. Chemokines and chemokine receptors: A new strategy for breast cancer therapy. Cancer Med. 2020, 9, 3786–3799. [Google Scholar] [CrossRef]
- Tao, J.J.; Visvanathan, K.; Wolff, A.C. Long term side effects of adjuvant chemotherapy in patients with early breast cancer. Breast 2015, 24 (Suppl. S2), S149–S153. [Google Scholar] [CrossRef] [Green Version]
- Zurrida, S.; Veronesi, U. Milestones in breast cancer treatment. Breast J. 2015, 21, 3–12. [Google Scholar] [CrossRef]
- Fahad Ullah, M. Breast Cancer: Current Perspectives on the Disease Status. Adv. Exp. Med. Biol. 2019, 1152, 51–64. [Google Scholar] [CrossRef]
- Mariotto, A.B.; Etzioni, R.; Hurlbert, M.; Penberthy, L.; Mayer, M. Estimation of the Number of Women Living with Metastatic Breast Cancer in the United States. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2017, 26, 809–815. [Google Scholar] [CrossRef] [Green Version]
- Teshome, M. Role of Operative Management in Stage IV Breast Cancer. Surg. Clin. N. Am. 2018, 98, 859–868. [Google Scholar] [CrossRef]
- Kocatürk, B.; Versteeg, H.H. Orthotopic injection of breast cancer cells into the mammary fat pad of mice to study tumor growth. J. Vis. Exp. JoVE 2015, 96, e51967. [Google Scholar] [CrossRef] [Green Version]
- DuPre, S.A.; Hunter, K.W., Jr. Murine mammary carcinoma 4T1 induces a leukemoid reaction with splenomegaly: Association with tumor-derived growth factors. Exp. Mol. Pathol. 2007, 82, 12–24. [Google Scholar] [CrossRef]
- Jafarzadeh, M.; Mousavizadeh, K.; Joghataei, M.T.; Bahremani, M.H.; Safa, M.; Asghari, S.M. A Fibroblast Growth Factor Antagonist Peptide Inhibits Breast Cancer in BALB/c Mice. Open Life Sci. 2018, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Disis, M.L.; Stanton, S.E. Immunotherapy in breast cancer: An introduction. Breast 2018, 37, 196–199. [Google Scholar] [CrossRef] [Green Version]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino-Mathews, A.; Foote, J.B.; Emens, L.A. Immune targeting in breast cancer. Oncology 2015, 29, 375–385. [Google Scholar] [PubMed]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [Green Version]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Chu, H.L.; Yip, B.S.; Chen, K.H.; Yu, H.Y.; Chih, Y.H.; Cheng, H.T.; Chou, Y.T.; Cheng, J.W. Novel antimicrobial peptides with high anticancer activity and selectivity. PLoS ONE 2015, 10, e0126390. [Google Scholar] [CrossRef] [Green Version]
- Orafaie, A.; Bahrami, A.R.; Matin, M.M. Use of anticancer peptides as an alternative approach for targeted therapy in breast cancer: A review. Nanomedicine 2021, 16, 415–433. [Google Scholar] [CrossRef]
- Mulder, K.C.; Lima, L.A.; Miranda, V.J.; Dias, S.C.; Franco, O.L. Current scenario of peptide-based drugs: The key roles of cationic antitumor and antiviral peptides. Front. Microbiol. 2013, 4, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms: My Perspective. Adv. Exp. Med. Biol. 2019, 1117, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Melo-Braga, M.N.; De Marco Almeida, F.; Dos Santos, D.M.; de Avelar Júnior, J.T.; Dos Reis, P.V.M.; de Lima, M.E. Antimicrobial Peptides From Lycosidae (Sundevall, 1833) Spiders. Curr. Protein Pept. Sci. 2020, 21, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Roudi, R.; Syn, N.L.; Roudbary, M. Antimicrobial Peptides As Biologic and Immunotherapeutic Agents against Cancer: A Comprehensive Overview. Front. Immunol. 2017, 8, 1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalzini, A.; Bergamini, C.; Biondi, B.; De Zotti, M.; Panighel, G.; Fato, R.; Peggion, C.; Bortolus, M.; Maniero, A.L. The rational search for selective anticancer derivatives of the peptide Trichogin GA IV: A multi-technique biophysical approach. Sci. Rep. 2016, 6, 24000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget 2017, 8, 46635–46651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Joncour, V.; Laakkonen, P. Seek & Destroy, use of targeting peptides for cancer detection and drug delivery. Bioorg. Med. Chem. 2018, 26, 2797–2806. [Google Scholar] [CrossRef]
- Liscano, Y.; Medina, L.; Oñate-Garzón, J.; Gúzman, F.; Pickholz, M.; Delgado, J.P. In Silico Selection and Evaluation of Pugnins with Antibacterial and Anticancer Activity Using Skin Transcriptome of Treefrog (Boana pugnax). Pharmaceutics 2021, 13, 578. [Google Scholar] [CrossRef]
- Yacoub, T.; Rima, M.; Karam, M.; Fajloun, J. Antimicrobials from Venomous Animals: An Overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- Santos, D.M.; Verly, R.M.; Piló-Veloso, D.; de Maria, M.; de Carvalho, M.A.; Cisalpino, P.S.; Soares, B.M.; Diniz, C.G.; Farias, L.M.; Moreira, D.F.; et al. LyeTx I, a potent antimicrobial peptide from the venom of the spider Lycosa erythrognatha. Amino Acids 2010, 39, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.V.M.; Boff, D.; Verly, R.M.; Melo-Braga, M.N.; Cortés, M.E.; Santos, D.M.; Pimenta, A.M.C.; Amaral, F.A.; Resende, J.M.; de Lima, M.E. LyeTxI-b, a Synthetic Peptide Derived From Lycosa erythrognatha Spider Venom, Shows Potent Antibiotic Activity in Vitro and in Vivo. Front. Microbiol. 2018, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.V.M.; Lima, V.M.; Souza, K.R.; Cardoso, G.A.; Melo-Braga, M.N.; Santos, D.M.; Verly, R.M.; Pimenta, A.M.C.; Dos Santos, V.L.; de Lima, M.E. Synthetic Peptides Derived From Lycosa Erythrognatha Venom: Interaction With Phospholipid Membranes and Activity Against Resistant Bacteria. Front. Mol. Biosci. 2021, 8, 680940. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.R.D.; Paiva, M.R.B.; Dourado, L.F.N.; Silva, R.O.; Silva, C.N.D.; Costa, B.L.D.; Toledo, C.R.; de Lima, M.E.; Silva-Cunha, A.D. Intravitreal injection of the synthetic peptide LyeTx I b, derived from a spider toxin, into the rabbit eye is safe and prevents neovascularization in a chorio-allantoic membrane model. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 31. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.N.D.; Silva, F.R.D.; Dourado, L.F.N.; Reis, P.; Silva, R.O.; Costa, B.L.D.; Nunes, P.S.; Amaral, F.A.; Santos, V.L.D.; de Lima, M.E.; et al. A New Topical Eye Drop Containing LyeTxI-b, A Synthetic Peptide Designed from A Lycosa erithrognata Venom Toxin, Was Effective to Treat Resistant Bacterial Keratitis. Toxins 2019, 11, 203. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Salam, M.A.L.; Carvalho-Tavares, J.; Gomes, K.S.; Teixeira-Carvalho, A.; Kitten, G.T.; Nyffeler, J.; Dias, F.F.; Dos Reis, P.V.M.; Pimenta, A.M.C.; Leist, M.; et al. The synthetic peptide LyeTxI-b derived from Lycosa erythrognatha spider venom is cytotoxic to U-87 MG glioblastoma cells. Amino Acids 2019, 51, 433–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
- Di, L. Strategic approaches to optimizing peptide ADME properties. AAPS J. 2015, 17, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Fang, M.; Alroy, J.; Sahagian, G.G. Imagable 4T1 model for the study of late stage breast cancer. BMC Cancer 2008, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, W., 3rd; Sosnoski, D.M.; Ohanessian, S.E.; Chandler, P.; Mobley, A.; Meisel, K.D.; Mastro, A.M. Role of Megakaryocytes in Breast Cancer Metastasis to Bone. Cancer Res. 2017, 77, 1942–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, T.; Doughty-Shenton, D.; Cassetta, L.; Fragkogianni, S.; Brownlie, D.; Kato, Y.; Carragher, N.; Pollard, J.W. Monocytes Differentiate to Immune Suppressive Precursors of Metastasis-Associated Macrophages in Mouse Models of Metastatic Breast Cancer. Front. Immunol. 2017, 8, 2004. [Google Scholar] [CrossRef] [Green Version]
- Walker Ii, W.H.; Borniger, J.C.; Surbhi; Zalenski, A.A.; Muscarella, S.L.; Fitzgerald, J.A.; Zhang, N.; Gaudier-Diaz, M.M.; DeVries, A.C. Mammary Tumors Induce Central Pro-inflammatory Cytokine Expression, but Not Behavioral Deficits in Balb/C Mice. Sci. Rep. 2017, 7, 8152. [Google Scholar] [CrossRef] [Green Version]
- Łakomska, I.; Hoffmann, K.; Wojtczak, A.; Sitkowski, J.; Maj, E.; Wietrzyk, J. Cytotoxic malonate platinum(II) complexes with 1,2,4-triazolo[1,5-a]pyrimidine derivatives: Structural characterization and mechanism of the suppression of tumor cell growth. J. Inorg. Biochem. 2014, 141, 188–197. [Google Scholar] [CrossRef]
- Rafehi, H.; Orlowski, C.; Georgiadis, G.T.; Ververis, K.; El-Osta, A.; Karagiannis, T.C. Clonogenic assay: Adherent cells. J. Vis. Exp. JoVE 2011, e2573. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Nuryadi, E.; Komatsu, S.; Hirota, Y.; Shibata, A.; Oike, T.; Nakano, T. Robustness of Clonogenic Assays as a Biomarker for Cancer Cell Radiosensitivity. Int. J. Mol. Sci. 2019, 20, 4148. [Google Scholar] [CrossRef] [Green Version]
- Douglas, S.; Hoskin, D.W.; Hilchie, A.L. Assessment of antimicrobial (host defense) peptides as anti-cancer agents. Methods Mol. Biol. 2014, 1088, 159–170. [Google Scholar] [CrossRef]
- Hao, W.; Hu, C.; Huang, Y.; Chen, Y. Coadministration of kla peptide with HPRP-A1 to enhance anticancer activity. PLoS ONE 2019, 14, e0223738. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tian, L.L.; Li, S.; Li, H.B.; Zhou, Y.; Wang, H.; Yang, Q.Z.; Ma, L.J.; Shang, D.J. Rapid cytotoxicity of antimicrobial peptide tempoprin-1CEa in breast cancer cells through membrane destruction and intracellular calcium mechanism. PLoS ONE 2013, 8, e60462. [Google Scholar] [CrossRef] [Green Version]
- Darzynkiewicz, Z. Critical aspects in analysis of cellular DNA content. Curr. Protoc. Cytom. 2010, 52, 7.2.1–7.2.8. [Google Scholar] [CrossRef]
- Marqus, S.; Pirogova, E.; Piva, T.J. Evaluation of the use of therapeutic peptides for cancer treatment. J. Biomed. Sci. 2017, 24, 21. [Google Scholar] [CrossRef] [Green Version]
- Mader, J.S.; Mookherjee, N.; Hancock, R.E.; Bleackley, R.C. The human host defense peptide LL-37 induces apoptosis in a calpain- and apoptosis-inducing factor-dependent manner involving Bax activity. Mol. Cancer Res. MCR 2009, 7, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087288. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, T.F.; Shi, Y.; Zhou, H.W.; Chen, Q.; Wei, B.Y.; Wang, X.; Yang, T.X.; Chinn, Y.E.; Kang, J.; et al. PFR peptide, one of the antimicrobial peptides identified from the derivatives of lactoferrin, induces necrosis in leukemia cells. Sci. Rep. 2016, 6, 20823. [Google Scholar] [CrossRef]
- Wang, C.; Dong, S.; Zhang, L.; Zhao, Y.; Huang, L.; Gong, X.; Wang, H.; Shang, D. Cell surface binding, uptaking and anticancer activity of L-K6, a lysine/leucine-rich peptide, on human breast cancer MCF-7 cells. Sci. Rep. 2017, 7, 8293. [Google Scholar] [CrossRef]
- Richter, W.F.; Bhansali, S.G.; Morris, M.E. Mechanistic determinants of biotherapeutics absorption following SC administration. AAPS J. 2012, 14, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Kagan, L. Pharmacokinetic modeling of the subcutaneous absorption of therapeutic proteins. Drug Metab. Dispos. Biol. Fate Chem. 2014, 42, 1890–1905. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.; Kerkis, A.; Hayashi, M.A.; Pereira, A.S.; Silva, F.S.; Oliveira, E.B.; Prieto da Silva, A.R.; Yamane, T.; Rádis-Baptista, G.; Kerkis, I. Crotamine toxicity and efficacy in mouse models of melanoma. Expert Opin. Investig. Drugs 2011, 20, 1189–1200. [Google Scholar] [CrossRef]
- Iadocicco, K.; Monteiro, L.H.; Chaui-Berlinck, J.G. A theoretical model for estimating the margination constant of leukocytes. BMC Physiol. 2002, 2, 3. [Google Scholar] [CrossRef]
- Strell, C.; Entschladen, F. Extravasation of leukocytes in comparison to tumor cells. Cell Commun. Signal. CCS 2008, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Borsig, L. Selectins in cancer immunity. Glycobiology 2018, 28, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturi, G.M.; Tu, L.; Kadono, T.; Khan, A.I.; Fujimoto, Y.; Oshel, P.; Bock, C.B.; Miller, A.S.; Albrecht, R.M.; Kubes, P.; et al. Leukocyte migration is regulated by L-selectin endoproteolytic release. Immunity 2003, 19, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Janssen, L.M.E.; Ramsay, E.E.; Logsdon, C.D.; Overwijk, W.W. The immune system in cancer metastasis: Friend or foe? J. Immunother. Cancer 2017, 5, 79. [Google Scholar] [CrossRef]
- Shand, F.H.; Ueha, S.; Otsuji, M.; Koid, S.S.; Shichino, S.; Tsukui, T.; Kosugi-Kanaya, M.; Abe, J.; Tomura, M.; Ziogas, J.; et al. Tracking of intertissue migration reveals the origins of tumor-infiltrating monocytes. Proc. Natl. Acad. Sci. USA 2014, 111, 7771–7776. [Google Scholar] [CrossRef] [Green Version]
- Naito, T.; Baba, T.; Takeda, K.; Sasaki, S.; Nakamoto, Y.; Mukaida, N. High-dose cyclophosphamide induces specific tumor immunity with concomitant recruitment of LAMP1/CD107a-expressing CD4-positive T cells into tumor sites. Cancer Lett. 2015, 366, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Galluzzi, L.; Buqué, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunological Effects of Conventional Chemotherapy and Targeted Anticancer Agents. Cancer Cell 2015, 28, 690–714. [Google Scholar] [CrossRef] [Green Version]
- Shrihari, T.G. Dual role of inflammatory mediators in cancer. Ecancermedicalscience 2017, 11, 721. [Google Scholar] [CrossRef]
- Buenrostro, D.; Kwakwa, K.A.; Putnam, N.E.; Merkel, A.R.; Johnson, J.R.; Cassat, J.E.; Sterling, J.A. Early TGF-β inhibition in mice reduces the incidence of breast cancer induced bone disease in a myeloid dependent manner. Bone 2018, 113, 77–88. [Google Scholar] [CrossRef]
- Liu, J.; Liao, S.; Diop-Frimpong, B.; Chen, W.; Goel, S.; Naxerova, K.; Ancukiewicz, M.; Boucher, Y.; Jain, R.K.; Xu, L. TGF-β blockade improves the distribution and efficacy of therapeutics in breast carcinoma by normalizing the tumor stroma. Proc. Natl. Acad. Sci. USA 2012, 109, 16618–16623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEarchern, J.A.; Kobie, J.J.; Mack, V.; Wu, R.S.; Meade-Tollin, L.; Arteaga, C.L.; Dumont, N.; Besselsen, D.; Seftor, E.; Hendrix, M.J.; et al. Invasion and metastasis of a mammary tumor involves TGF-beta signaling. Int. J. Cancer 2001, 91, 76–82. [Google Scholar] [CrossRef]
- Sheng, Y.; Li, F.; Qin, Z. TNF Receptor 2 Makes Tumor Necrosis Factor a Friend of Tumors. Front. Immunol. 2018, 9, 1170. [Google Scholar] [CrossRef] [Green Version]
- Sasi, S.P.; Bae, S.; Song, J.; Perepletchikov, A.; Schneider, D.; Carrozza, J.; Yan, X.; Kishore, R.; Enderling, H.; Goukassian, D.A. Therapeutic non-toxic doses of TNF induce significant regression in TNFR2-p75 knockdown Lewis lung carcinoma tumor implants. PLoS ONE 2014, 9, e92373. [Google Scholar] [CrossRef] [Green Version]
- Eftekhari, R.; Esmaeili, R.; Mirzaei, R.; Bidad, K.; de Lima, S.; Ajami, M.; Shirzad, H.; Hadjati, J.; Majidzadeh, A.K. Study of the tumor microenvironment during breast cancer progression. Cancer Cell Int. 2017, 17, 123. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.M.; Varghese, S.; Xu, H.; Alexander, H.R. Interleukin-1 and cancer progression: The emerging role of interleukin-1 receptor antagonist as a novel therapeutic agent in cancer treatment. J. Transl. Med. 2006, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Holen, I.; Lefley, D.V.; Francis, S.E.; Rennicks, S.; Bradbury, S.; Coleman, R.E.; Ottewell, P. IL-1 drives breast cancer growth and bone metastasis in vivo. Oncotarget 2016, 7, 75571–75584. [Google Scholar] [CrossRef] [Green Version]
- Mumm, J.B.; Emmerich, J.; Zhang, X.; Chan, I.; Wu, L.; Mauze, S.; Blaisdell, S.; Basham, B.; Dai, J.; Grein, J.; et al. IL-10 elicits IFNγ-dependent tumor immune surveillance. Cancer Cell 2011, 20, 781–796. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Ammar, A.; Storr, S.J.; Green, A.R.; Rakha, E.; Ellis, I.O.; Martin, S.G. IL-6 and IL-10 are associated with good prognosis in early stage invasive breast cancer patients. Cancer Immunol. Immunother. CII 2018, 67, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Stojanovska, V.; McQuade, R.M.; Fraser, S.; Prakash, M.; Gondalia, S.; Stavely, R.; Palombo, E.; Apostolopoulos, V.; Sakkal, S.; Nurgali, K. Oxaliplatin-induced changes in microbiota, TLR4+ cells and enhanced HMGB1 expression in the murine colon. PLoS ONE 2018, 13, e0198359. [Google Scholar] [CrossRef]
- Elsea, C.R.; Kneiss, J.A.; Wood, L.J. Induction of IL-6 by Cytotoxic Chemotherapy Is Associated With Loss of Lean Body and Fat Mass in Tumor-free Female Mice. Biol. Res. Nurs. 2015, 17, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Volk-Draper, L.; Hall, K.; Griggs, C.; Rajput, S.; Kohio, P.; DeNardo, D.; Ran, S. Paclitaxel therapy promotes breast cancer metastasis in a TLR4-dependent manner. Cancer Res. 2014, 74, 5421–5434. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Diana, J. The Dual Role of Antimicrobial Peptides in Autoimmunity. Front. Immunol. 2020, 11, 2077. [Google Scholar] [CrossRef]
- Percoco, G.; Merle, C.; Jaouen, T.; Ramdani, Y.; Bénard, M.; Hillion, M.; Mijouin, L.; Lati, E.; Feuilloley, M.; Lefeuvre, L.; et al. Antimicrobial peptides and pro-inflammatory cytokines are differentially regulated across epidermal layers following bacterial stimuli. Exp. Dermatol. 2013, 22, 800–806. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A. Antimicrobial/anticancer peptides: Bioactive molecules and therapeutic agents. Immunotherapy 2021, 13, 669–684. [Google Scholar] [CrossRef]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paull, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigro-Wolff, A.; et al. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor cell lines. J. Natl. Cancer Inst. 1991, 83, 757–766. [Google Scholar] [CrossRef]
- Perdigão, G.M.C.; Lopes, M.S.; Marques, L.B.; Prazeres, P.; Gomes, K.S.; de Oliveira, R.B.; Pinto, M.C.X.; de Souza-Fagundes, E.M. Novel nitroaromatic compound activates autophagy and apoptosis pathways in HL60 cells. Chem.-Biol. Interact. 2018, 283, 107–115. [Google Scholar] [CrossRef]
- Nicoletti, I.; Migliorati, G.; Pagliacci, M.C.; Grignani, F.; Riccardi, C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Methods 1991, 139, 271–279. [Google Scholar] [CrossRef]
- Paschall, A.V.; Liu, K. An Orthotopic Mouse Model of Spontaneous Breast Cancer Metastasis. J. Vis. Exp. JoVE 2016. [Google Scholar] [CrossRef]
- Szczepanski, C.; Tenstad, O.; Baumann, A.; Martinez, A.; Myklebust, R.; Bjerkvig, R.; Prestegarden, L. Identification of a novel lytic peptide for the treatment of solid tumours. Genes Cancer 2014, 5, 186–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulzele, S.V.; Chatterjee, A.; Shaik, M.S.; Jackson, T.; Singh, M. Inhalation delivery and anti-tumor activity of celecoxib in human orthotopic non-small cell lung cancer xenograft model. Pharm. Res. 2006, 23, 2094–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitson, T.D.; Wigg, B.; Becker, G.J. Tissue preparation for histochemistry: Fixation, embedding, and antigen retrieval for light microscopy. Methods Mol. Biol. 2010, 611, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Dutra, A.P.; Azevedo Júnior, G.M.; Schmitt, F.C.; Cassali, G.D. Assessment of cell proliferation and prognostic factors in canine mammary gland tumors. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2008, 60, 1403–1412. [Google Scholar] [CrossRef]
- Oliveira-Lima, O.C.; Bernardes, D.; Xavier Pinto, M.C.; Esteves Arantes, R.M.; Carvalho-Tavares, J. Mice lacking inducible nitric oxide synthase develop exacerbated hepatic inflammatory responses induced by Plasmodium berghei NK65 infection. Microbes Infect. 2013, 15, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Lima, O.C.; Pinto, M.C.; Duchene, J.; Qadri, F.; Souza, L.L.; Alenina, N.; Bader, M.; Santos, R.A.; Carvalho-Tavares, J. Mas receptor deficiency exacerbates lipopolysaccharide-induced cerebral and systemic inflammation in mice. Immunobiology 2015, 220, 1311–1321. [Google Scholar] [CrossRef]
- Van Fraga, I.T.; Limborço-Filho, M.; Lima, O.C.; Lacerda-Queiroz, N.; Guidine, P.A.; Moraes, M.F.; Nascimento Araújo, R.; Moraes-Santos, T.; Massensini, A.R.; Arantes, R.M.; et al. Effects of tityustoxin on cerebral inflammatory response in young rats. Neurosci. Lett. 2015, 588, 24–28. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | LyeTx I-b IC50 (µM) | Carboplatin IC50 (µM) |
|---|---|---|
| 4T1 | 6.5 ± 5.30 | ≥100 |
| MCF-7 | 7.34 ± 3.09 | ≥100 |
| MDA-MB-231 | 5.77 ± 0.83 | 69.20 ± 1.22 |
| The IC50 values were calculated based on the linear regression of the dose-log response curves after 48 h exposure to the peptide or carboplatin, determined by the MTT and resazurin assays. The values represent the mean ± S.D. of at least 3 independent experiments. Representative concentration-response curves are displayed below. | ||
 | ||
| Hematological Parameter | Control | LyeTx I_b Peptide |
|---|---|---|
| RBC (×106/mm3) | 6.32 ± 0.667 | 7.46 ± 0.965 |
| Hb (g/dL) | 10.8 ± 2.66 | 12.9 ± 0.680 |
| Hematocrit | 38.9 ± 8.66 | 39.4 ± 2.56 |
| MCV (fL) | 61 ± 7.57 | 48 ± 4.50 |
| MCH (Pg) | 16.7 ± 0.458 | 16.0 ± 0.11 |
| MCHC % | 28.5 ± 3.06 | 31.6 ± 1.34 |
| WBC (×103/mm3) | 2.3 ± 2.26 | 4.1 ± 0.70 |
| Lymphocytes | 91.7 ± 5.13 | 89.2 ± 4.03 |
| Monocytes | 4.3 ± 1.556 | 4.1 ± 1.553 |
| Neutrophils | 4 ± 3.59 | 6.7 ± 2.66 |
| Platelets (×103/mm3) | 443 ± 73.6 | 278 ± 80.0 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Salam, M.A.L.; Pinto, B.; Cassali, G.; Bueno, L.; Pêgas, G.; Oliveira, F.; Silva, I.; Klein, A.; Souza-Fagundes, E.M.d.; de Lima, M.E.; et al. LyeTx I-b Peptide Attenuates Tumor Burden and Metastasis in a Mouse 4T1 Breast Cancer Model. Antibiotics 2021, 10, 1136. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091136
Abdel-Salam MAL, Pinto B, Cassali G, Bueno L, Pêgas G, Oliveira F, Silva I, Klein A, Souza-Fagundes EMd, de Lima ME, et al. LyeTx I-b Peptide Attenuates Tumor Burden and Metastasis in a Mouse 4T1 Breast Cancer Model. Antibiotics. 2021; 10(9):1136. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091136
Chicago/Turabian StyleAbdel-Salam, Mostafa A. L., Bárbara Pinto, Geovanni Cassali, Lilian Bueno, Gabriela Pêgas, Fabrício Oliveira, Irismara Silva, André Klein, Elaine Maria de Souza-Fagundes, Maria Elena de Lima, and et al. 2021. "LyeTx I-b Peptide Attenuates Tumor Burden and Metastasis in a Mouse 4T1 Breast Cancer Model" Antibiotics 10, no. 9: 1136. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10091136