
Phylogenetic Groups and Antimicrobial Resistance Genes in Escherichia coli from Different Meat Species

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Phenotypical Resistance
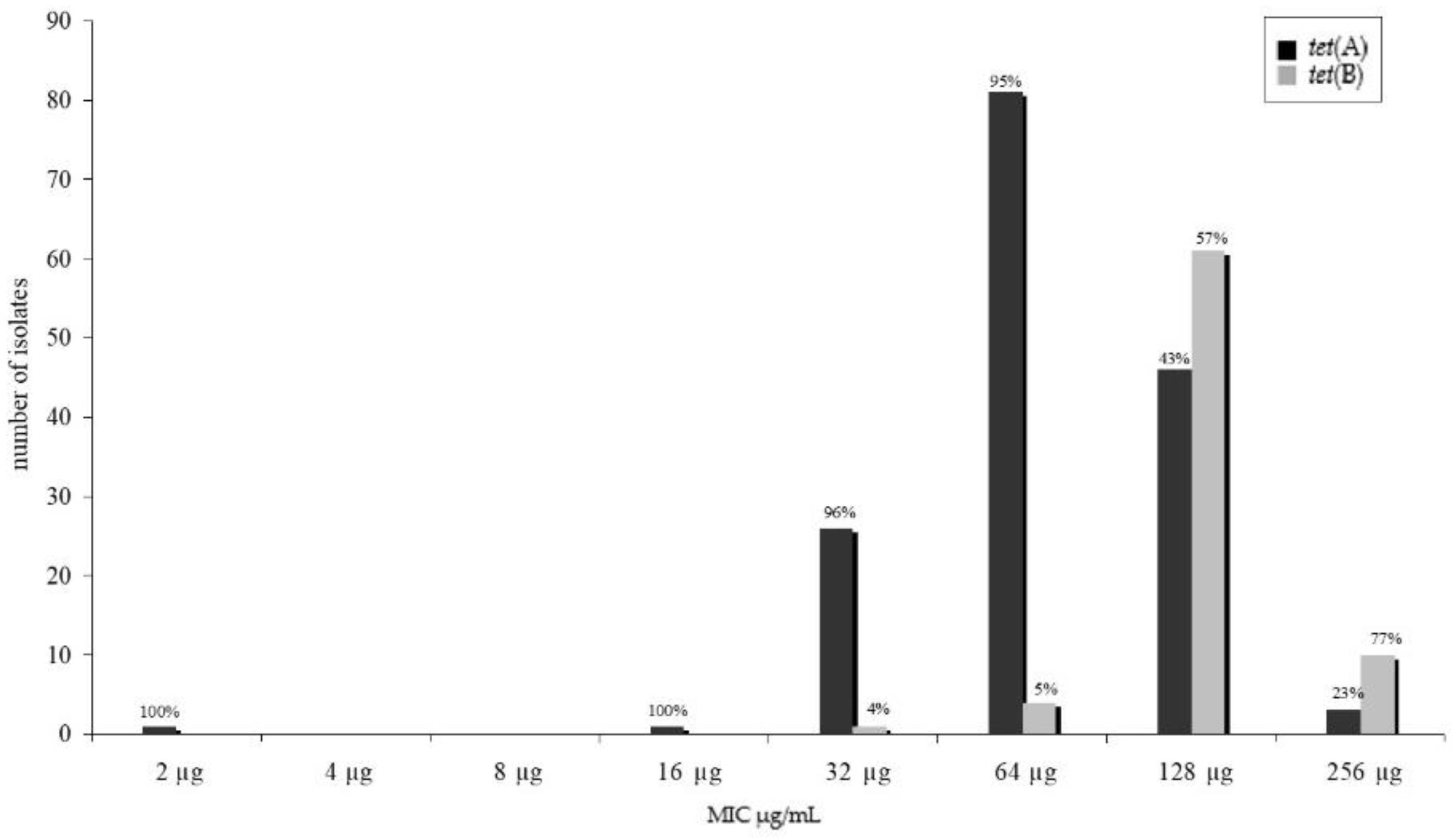
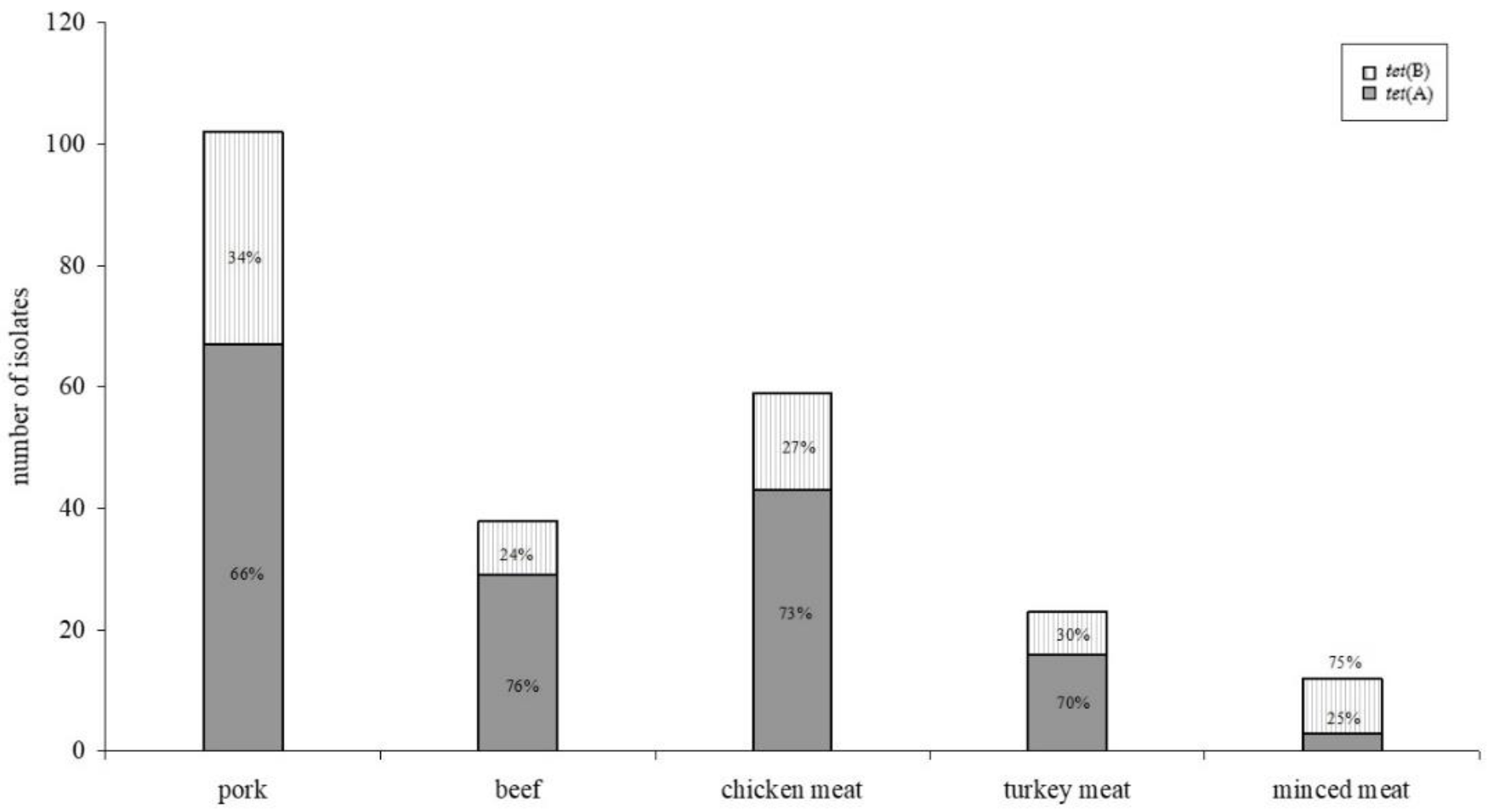
2.2. Tetracycline Resistance Genes
2.3. Sulphonamide Resistance Genes
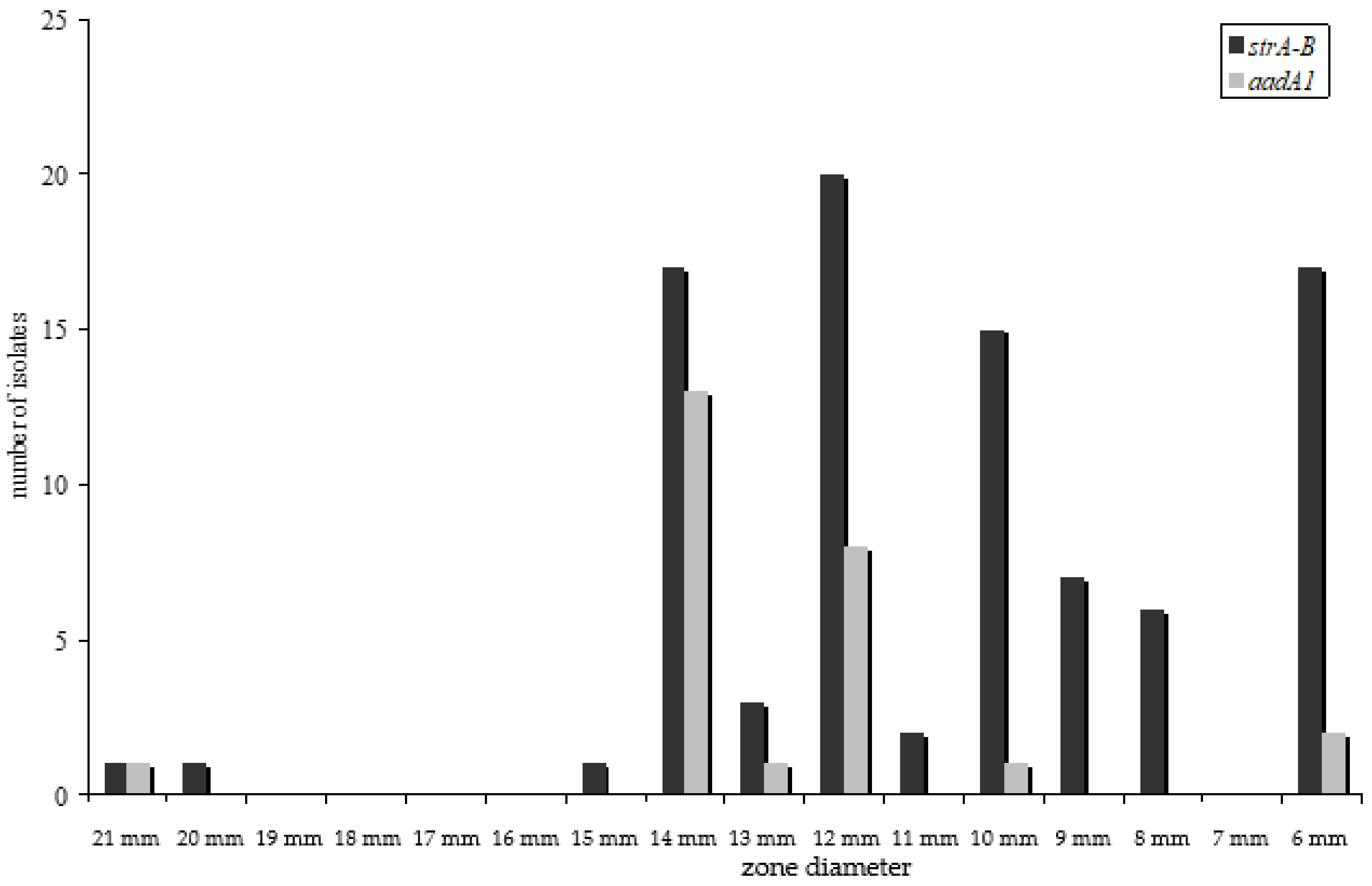
2.4. Streptomycin Resistance Genes
2.5. Ampicillin Resistance Genes
2.6. Trimethoprim Resistance Genes
2.7. Kanamycin Resistanc Genes
2.8. Multi-Resistant Isolates and Gene Association
2.9. Phylogenetic Groups
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Scientific Opinion on the whole genome sequencing and metagenomics for outbreak investigation, source attribution and risk assessment of food-borne microorganisms. EFSA J. 2019, 17, e05898. [Google Scholar] [CrossRef] [Green Version]
- Llarena, A.-K.; Ribeiro-Gonçalves, B.F.; Nuno Silva, D.; Halkilahti, J.; Machado, M.P.; Da Silva, M.S.; Jaakkonen, A.; Isidro, J.; Hämäläinen, C.; Joenperä, J.; et al. INNUENDO: A cross-sectoral platform for the integration of genomics in the surveillance of food-borne pathogens. EFSA Support. Publ. 2018, 15, 142p. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartof, S.Y.; Solberg, O.D.; Riley, L.W. Genotypic analyses of uropathogenic Escherichia coli based on fimH single nucleotide polymorphisms (SNPs). J. Med Microbiol. 2007, 56, 1363–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, S.J.; Hansen, N.I.; Zaterka-Baxter, K.; Higgins, R.D.; Stoll, B.J. Emergence of Antibiotic Resistance-Associated Clones AmongEscherichia coliRecovered From Newborns with Early-Onset Sepsis and Meningitis in the United States, 2008–2009. J. Pediatr. Infect. Dis. Soc. 2016, 5, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratshilingano, M.T.; du Plessis, E.M.; Duvenage, S.; Korsten, L. Characterisation of multidrug resistant Escherichia coli isolated from two commercial lettuce and spinach supply chains. J. Food Prot. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Olesen, B.; Scheutz, F.; Menard, M.; Skov, M.N.; Kolmos, H.J.; Kuskowski, M.A.; Johnson, J.R. Three-Decade Epidemiological Analysis of Escherichia coli O15:K52:H1. J. Clin. Microbiol. 2009, 47, 1857–1862. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Shibata, N.; Yamane, K.; Wachino, J.-I.; Ito, K.; Arakawa, Y. Change in the prevalence of extended-spectrum- -lactamase-producing Escherichia coli in Japan by clonal spread. J. Antimicrob. Chemother. 2009, 63, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Ciccozzi, M.; Giufre, M.; Accogli, M.; Presti, A.L.; Graziani, C.; Cella, E.; Cerquetti, M. Phylogenetic analysis of multidrug-resistant Escherichia coli clones isolated from humans and poultry. New Microbiol. 2013, 36, 385–394. [Google Scholar]
- Guerra, S.T.; Dalanezi, F.M.; De Paula, C.L.; Hernandes, R.T.; Pantoja, J.C.D.F.; Listoni, F.J.P.; Langoni, H.; Ribeiro, M.G. Putative virulence factors of extra-intestinalEscherichia coliisolated from bovine mastitis with different clinical scores. Lett. Appl. Microbiol. 2019, 68, 403–408. [Google Scholar] [CrossRef]
- Khan, S.B.; Zou, G.; Cheng, Y.-T.; Xiao, R.; Li, L.; Wu, B.; Zhou, R. Phylogenetic grouping and distribution of virulence genes in Escherichia coli along the production and supply chain of pork around Hubei, China. J. Microbiol. Immunol. Infect. 2017, 50, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheinberg, J.; Dudley, E.G.; Campbell, J.; Roberts, B.; DiMarzio, M.; Debroy, C.; Cutter, C.N. Prevalence and Phylogenetic Characterization of Escherichia coli and Hygiene Indicator Bacteria Isolated from Leafy Green Produce, Beef, and Pork Obtained from Farmers’ Markets in Pennsylvania. J. Food Prot. 2017, 80, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal Pathogenic Escherichia coli Strains of Avian and Human Origin: Link between Phylogenetic Relationships and Common Virulence Patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Mayrhofer, S.; Paulsen, P.; Smulders, F.J.; Hilbert, F. Antimicrobial Resistance in Commensal Escherichia coli Isolated from Muscle Foods as Related to the Veterinary Use of Antimicrobial Agents in Food-Producing Animals in Austria. Microb. Drug Resist. 2006, 12, 278–283. [Google Scholar] [CrossRef]
- Ryu, S.-H.; Lee, J.-H.; Park, S.-H.; Song, M.-O.; Park, S.-H.; Jung, H.-W.; Park, G.-Y.; Choi, S.-M.; Kim, M.-S.; Chae, Y.-Z.; et al. Antimicrobial resistance profiles among Escherichia coli strains isolated from commercial and cooked foods. Int. J. Food Microbiol. 2012, 159, 263–266. [Google Scholar] [CrossRef] [PubMed]
- CLSI 2019. Performance Standards for Antimicrobial Susceptibility Testing M100, 29th ed.; CLSI: Annapolis Junction, MD, USA, 2019. [Google Scholar]
- Furlan, J.P.R.; Stehling, E.G. Multiple sequence types, virulence determinants and antimicrobial resistance genes in multidrug- and colistin-resistant Escherichia coli from agricultural and non-agricultural soils. Environ. Pollut. 2021, 288, 117804. [Google Scholar] [CrossRef]
- Sheikh, A.A.; Checkley, S.; Avery, B.; Chalmers, G.; Bohaychuk, V.; Boerlin, P.; Reid-Smith, R.; Aslam, M. Antimicrobial Resistance and Resistance Genes in Escherichia coli Isolated from Retail Meat Purchased in Alberta, Canada. Foodborne Pathog. Dis. 2012, 9, 625–631. [Google Scholar] [CrossRef]
- Pietsch, M.; Irrgang, A.; Roschanski, N.; Brenner, M.G.; Hamprecht, A.; Rieber, H.; Käsbohrer, A.; Schwarz, S.; Rösler, U.; Kreienbrock, L.; et al. Whole genome analyses of CMY-2-producing Escherichia coli isolates from humans, animals and food in Germany. BMC Genom. 2018, 19, 601. [Google Scholar] [CrossRef]
- Touchon, M.; Perrin, A.; de Sousa, J.A.M.; Vangchhia, B.; Burn, S.; O’Brien, C.L.; Denamur, E.; Gordon, D.; Rocha, E.P. Phylogenetic background and habitat drive the genetic diversification of Escherichia coli. PLoS Genet. 2020, 16, e1008866. [Google Scholar] [CrossRef]
- Wadepohl, K.; Müller, A.; Seinige, D.; Rohn, K.; Blaha, T.; Meemken, D.; Kehrenberg, C. Association of intestinal colonization of ESBL-producing Enterobacteriaceae in poultry slaughterhouse workers with occupational exposure—A German pilot study. PLoS ONE 2020, 15, e0232326. [Google Scholar] [CrossRef] [PubMed]
- Boerlin, P.; Travis, R.; Gyles, C.L.; Reid-Smith, R.; Lim, N.J.H.; Nicholson, V.; McEwen, S.A.; Friendship, R.; Archambault, M. Antimicrobial Resistance and Virulence Genes of Escherichia coli Isolates from Swine in Ontario. Appl. Environ. Microbiol. 2005, 71, 6753–6761. [Google Scholar] [CrossRef] [Green Version]
- Sunde, M.; Norström, M. The genetic background for streptomycin resistance in Escherichia coli influences the distribution of MICs. J. Antimicrob. Chemother. 2005, 56, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Jaffé, A.; A Chabbert, Y.; Derlot, E. Selection and characterization of beta-lactam-resistant Escherichia coli K-12 mutants. Antimicrob. Agents Chemother. 1983, 23, 622–625. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.F.; Fritsch, E.F.; Maniatis, T. (Eds.) Molecular Cloning: A Laboratory Manual; Cold Spring Habor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Ng, L.-K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, S.; White, D.G.; Schroeder, C.M.; Lu, R.; Yang, H.; McDermott, P.F.; Ayers, S.; Meng, J. Characterization of Multiple-Antimicrobial-Resistant Salmonella Serovars Isolated from Retail Meats. Appl. Environ. Microbiol. 2004, 70, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic Resistance in the ECOR Collection: Integrons and Identification of a Novel aad Gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Kerrn, M.B.; Klemmensen, T.; Frimodt-Møller, N.; Espersen, F. Susceptibility of Danish Escherichia coli strains isolated from urinary tract infections and bacteraemia, and distribution of sul genes conferring sulphonamide resistance. J. Antimicrob. Chemother. 2002, 50, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Maidhof, H.; Guerra, B.; Abbas, S.; Elsheikha, H.M.; Whittam, T.S.; Beutin, L. A Multiresistant Clone of Shiga Toxin-Producing Escherichia coli O118:[H16] Is Spread in Cattle and Humans over Different European Countries. Appl. Environ. Microbiol. 2002, 68, 5834–5842. [Google Scholar] [CrossRef] [Green Version]
- Henriques, I.; Moura, A.; Alves, A.; Saavedra, M.J.; Correia, A. Analysing diversity among Î2-lactamase encoding genes in aquatic environments. FEMS Microbiol. Ecol. 2006, 56, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navia, M.M.; Ruiz, J.; Sanchez-Cespedes, J.; Vila, J. Detection of dihydrofolate reductase genes by PCR and RFLP. Diagn. Microbiol. Infect. Dis. 2003, 46, 295–298. [Google Scholar] [CrossRef]
- Grape, M.; Motakefi, A.; Pavuluri, S.; Kahlmeter, G. Standard and real-time multiplex PCR methods for detection of trimethoprim resistance dfr genes in large collections of bacteria. Clin. Microbiol. Infect. 2007, 13, 1112–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandvang, D.; Aarestrup, F.M. Characterization of Aminoglycoside Resistance Genes and Class 1 Integrons in Porcine and Bovine Gentamicin-ResistantEscherichia coli. Microb. Drug Resist. 2000, 6, 19–27. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| tet(A) n = 158 | tet(B) n = 76 | sulI n = 33 | sulII n = 114 | blaTEM n = 93 | dfrA n = 71 | dfr7&17 n = 3 | aphA1 n = 28 | strA–B n = 89 | addA1 n = 26 | |
|---|---|---|---|---|---|---|---|---|---|---|
| tet(A) | - | 2 | 17 | 55 | 47 | 43 | 2 | 13 | 54 | 10 |
| tet(B) | - | 10 | 37 | 21 | 16 | 1 | 13 | 19 | 8 | |
| sulI | - | 3 | 12 | 10 | 0 | 5 | 4 | 3 | ||
| sulII | - | 53 | 58 | 3 | 19 | 65 | 4 | |||
| blaTEM | - | 42 | 3 | 17 | 41 | 7 | ||||
| dfrA | - | 0 | 9 | 42 | 3 | |||||
| dfr7&17 | - | 1 | 1 | 0 | ||||||
| aphA1 | - | 17 | 2 | |||||||
| strA–B | - | 3 | ||||||||
| aadA1 | - |
| Meat Species | Group A | Group B1 | Group B2 | Group D | Number |
|---|---|---|---|---|---|
| Pork | 86 | 29 | 4 | 3 | 122 |
| Chicken meat | 56 | 28 | 6 | 9 | 99 |
| Beef | 35 | 15 | 1 | 2 | 53 |
| Turkey meat | 16 | 4 | 0 | 4 | 24 |
| Minced meat | 11 | 4 | 0 | 0 | 15 |
| Primer Name | Sequence | Target Gene | Amplicon Size | Annealing Temp (°C) | Reference |
|---|---|---|---|---|---|
| tet(A)-f tet(A)-r | 5′-gctacatcctgcttgtgccttc-3′ 5′-catagatcgccgtgaagagg-3′ | tet(A) | 210 | 57 | [26] |
| tet(B)-f tet(B)-r | 5′-ttggttcggggcaagttttg-3′ 5′-gtaatgggccaataacaccg-3′ | tet(B) | 659 | 57 | [26] |
| tet(C)-f tet(C)-r | 5′-cttgagagccttcaacccag-3′ 5′-atggtcgtcatctacctgcc-3′ | tet(C) | 418 | 55 | [26] |
| tet(G)-f tet(G)-r | 5′-agcaggtcgctggacactat-3′ 5′-cgcggtgttccactgaaaac-3′ | tet(G) | 623 | 55 | [27] |
| sulI-f sulI-r | 5′-tggtgacggtgttcggcattc3′ 5′-gcgaaggtttccgagaaggtg-3′ | sulI | 790 | 63 | [28] |
| sulII-f sulII-r | 5′-gcgctcaaggcagatggcatt-3′ 5′-gcgtttgataccggcacccgt-3′ | sulII | 293 | 60 | [29] |
| strA–B-f strA_B-r | 5′-ccaatcgcagatagaaggcaag-3′ 5′-atcaactggcaggaggaacagg-3′ | strA | 580 | 65 | [30] |
| aadA1-f aadA1-r | 5′-aacgaccttttggaaacttcgg -3′ 5′-ttcgctcatcgccagcccag-3′ | aadA1 | 352 | 60 | [30] |
| blaCMY-f blaCMY-r | 5′-tggccgttgccgttatctac-3′ 5′-cccgttttatgcacccatga-3′ | blaCMY | 870 | 55 | [27] |
| blaIMP-f blaIMP-r | 5′-gaatagagtggattaattctc-3′ 5′-ggtttaayaaaacaaccacc-3′ | blaIMP | 232 | 55 | [31] |
| blaOXA-2-f blaOXA-2-r | 5′-caagccaaaggcacgatagttg-3′ 5′-ctcaacccatcctacccacc-3′ | blaOXA | 561 | 56 | [31] |
| blaPSE-f blaPSE-r | 5′-tgcttcgcaactatgactac-3′ 5′-agcctgtgtttgagctagat-3′ | blaPSE | 438 | 55 | [27] |
| blaTEM-1-f blaTEM-1-r | 5′-cagcggtaagatccttgaga-3′ 5′-actccccgtcgtgtagataa-3′ | blaTEM | 643 | 55 | [27] |
| dfr1-f dfr1-r | 5′-gtgaaactatcactaatgg-3′ 5′-ttaacccttttgccagattt-3′ | dfrA1, dfrA5, dfrA15, dfrA15b, dfrA16, dfrA16b | 474 | 55 | [32] |
| dfr2-f dfr2-r | 5′-gatcgcgtgcgcaagaaatc-3′ 5′-aagcgcagccacaggataaat-3′ | dfrB1, dfrB2, dfrB3 | 141 | 60 | [32] |
| dfr7&17-f dfr7&17-r | 5′-acatttgactctatgggtgttcttc-3′ 5′-aaaactgttcaaaaaccaaattgaa-3′ | dfr7&17 | 280 | 55 | [33] |
| aphA-1a-f aphA-1a-r | 5′-aacgtcttgctcgaggccgcg-3′ 5′-ggcaagatcctggtatcggtctgc-3′ | aphA-1a | 670 | 65 | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacher-Pirklbauer, A.; Klein-Jöbstl, D.; Sofka, D.; Blanc-Potard, A.-B.; Hilbert, F. Phylogenetic Groups and Antimicrobial Resistance Genes in Escherichia coli from Different Meat Species. Antibiotics 2021, 10, 1543. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121543
Sacher-Pirklbauer A, Klein-Jöbstl D, Sofka D, Blanc-Potard A-B, Hilbert F. Phylogenetic Groups and Antimicrobial Resistance Genes in Escherichia coli from Different Meat Species. Antibiotics. 2021; 10(12):1543. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121543
Chicago/Turabian StyleSacher-Pirklbauer, Angelika, Daniela Klein-Jöbstl, Dmitrij Sofka, Anne-Béatrice Blanc-Potard, and Friederike Hilbert. 2021. "Phylogenetic Groups and Antimicrobial Resistance Genes in Escherichia coli from Different Meat Species" Antibiotics 10, no. 12: 1543. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10121543