Protective Effects of Bacteriophages against Aeromonas hydrophila Causing Motile Aeromonas Septicemia (MAS) in Striped Catfish


Abstract
:1. Introduction
2. Results
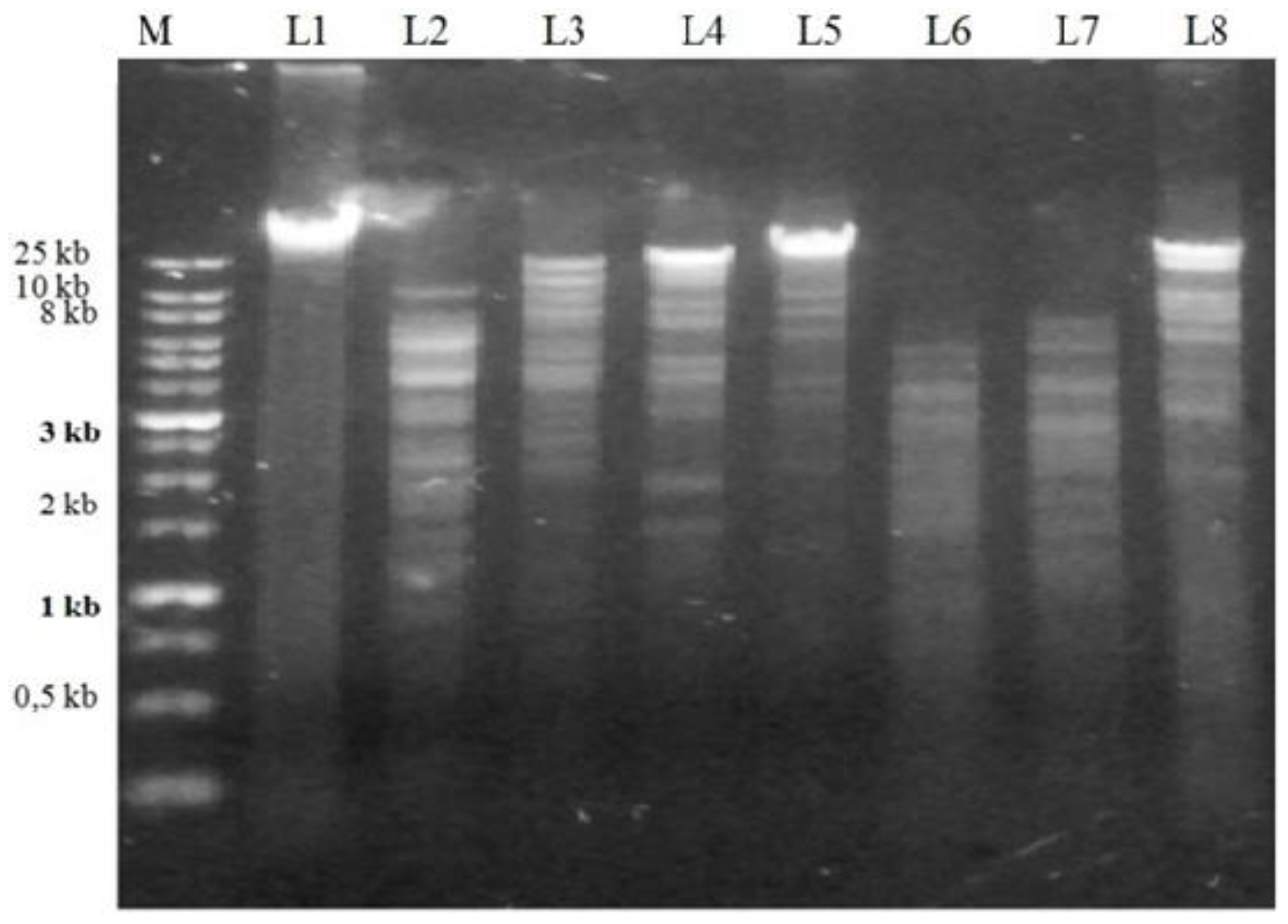
2.1. Prophage Induction
2.2. Antibiotic Susceptibility
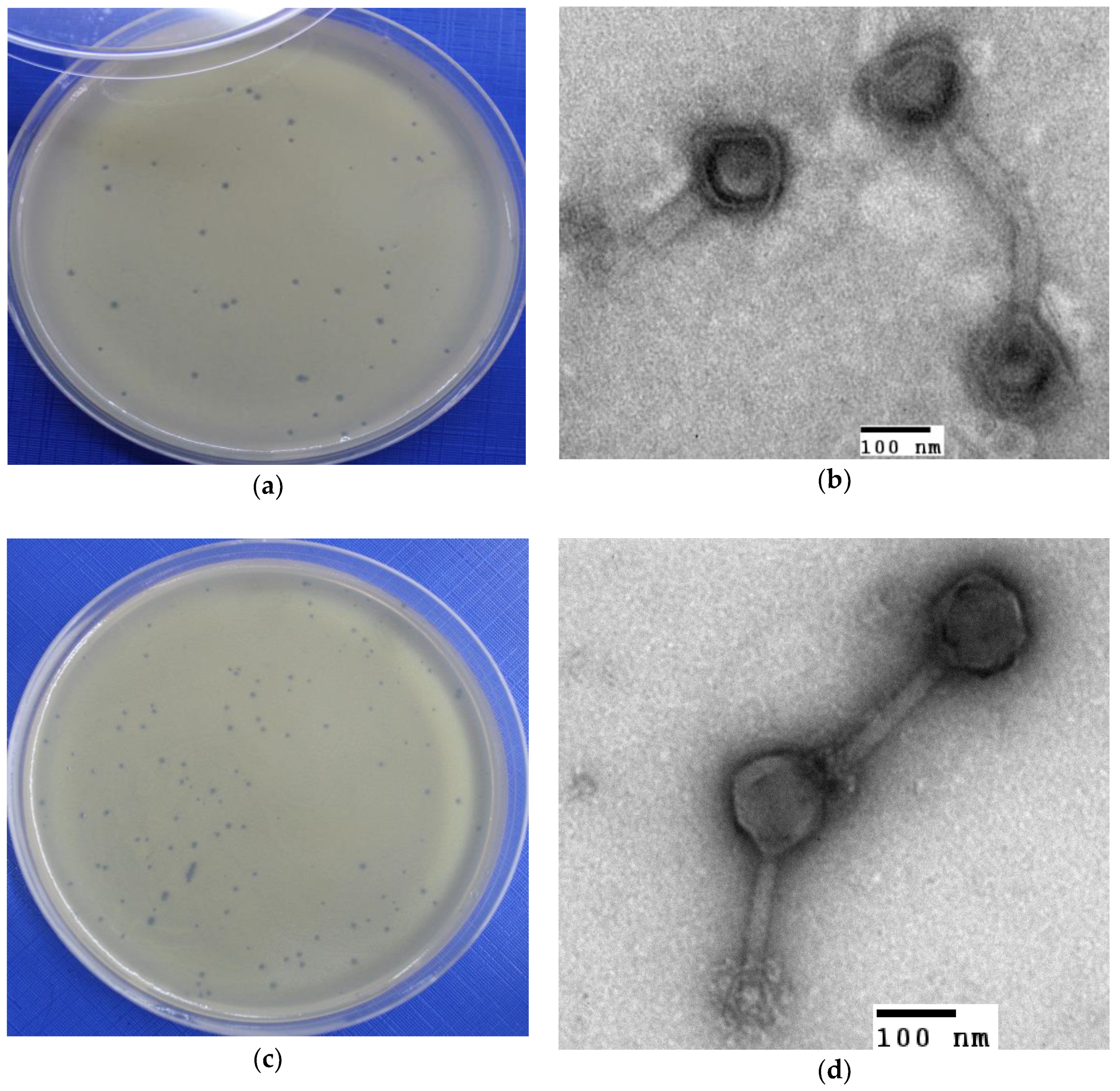
2.3. Isolation and Characterization of Bacteriophages
2.4. Host Range
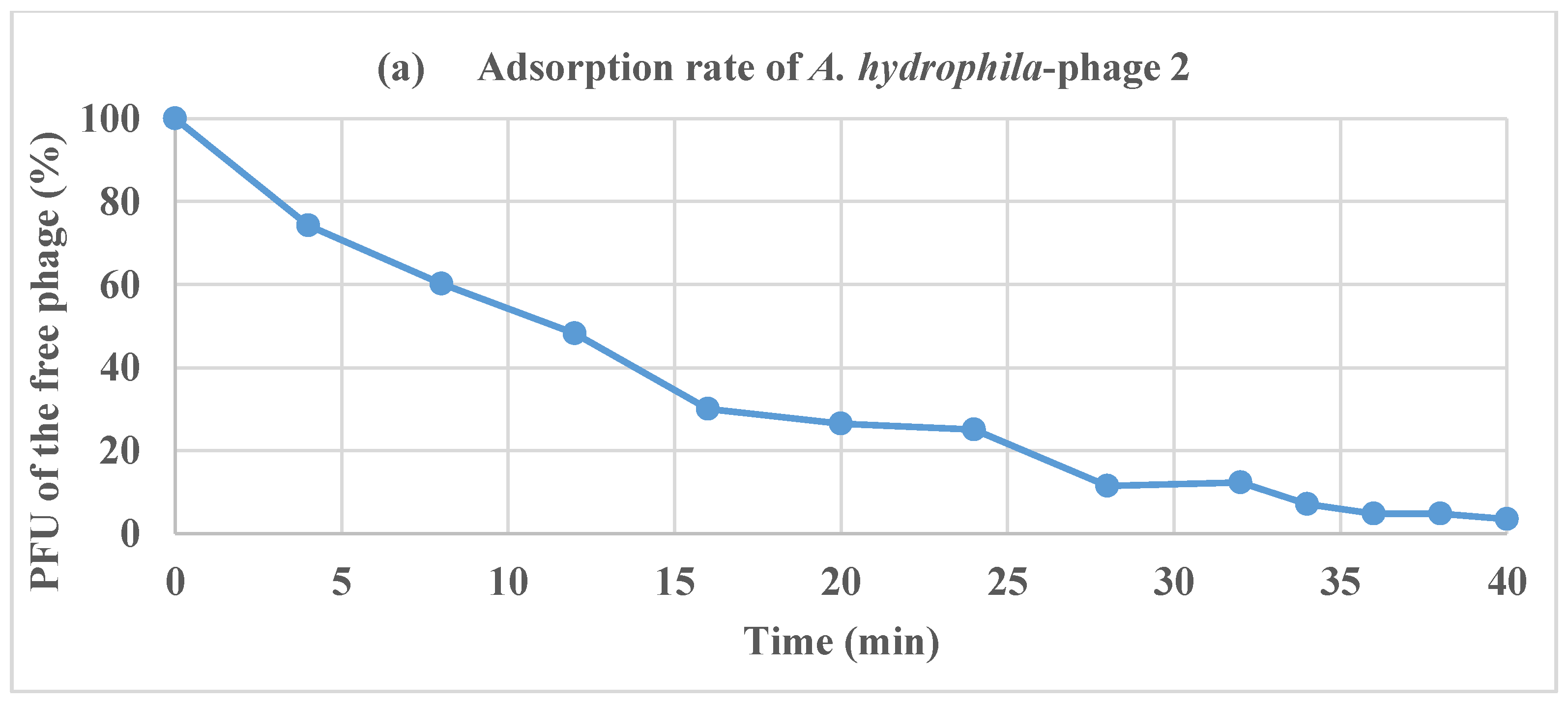
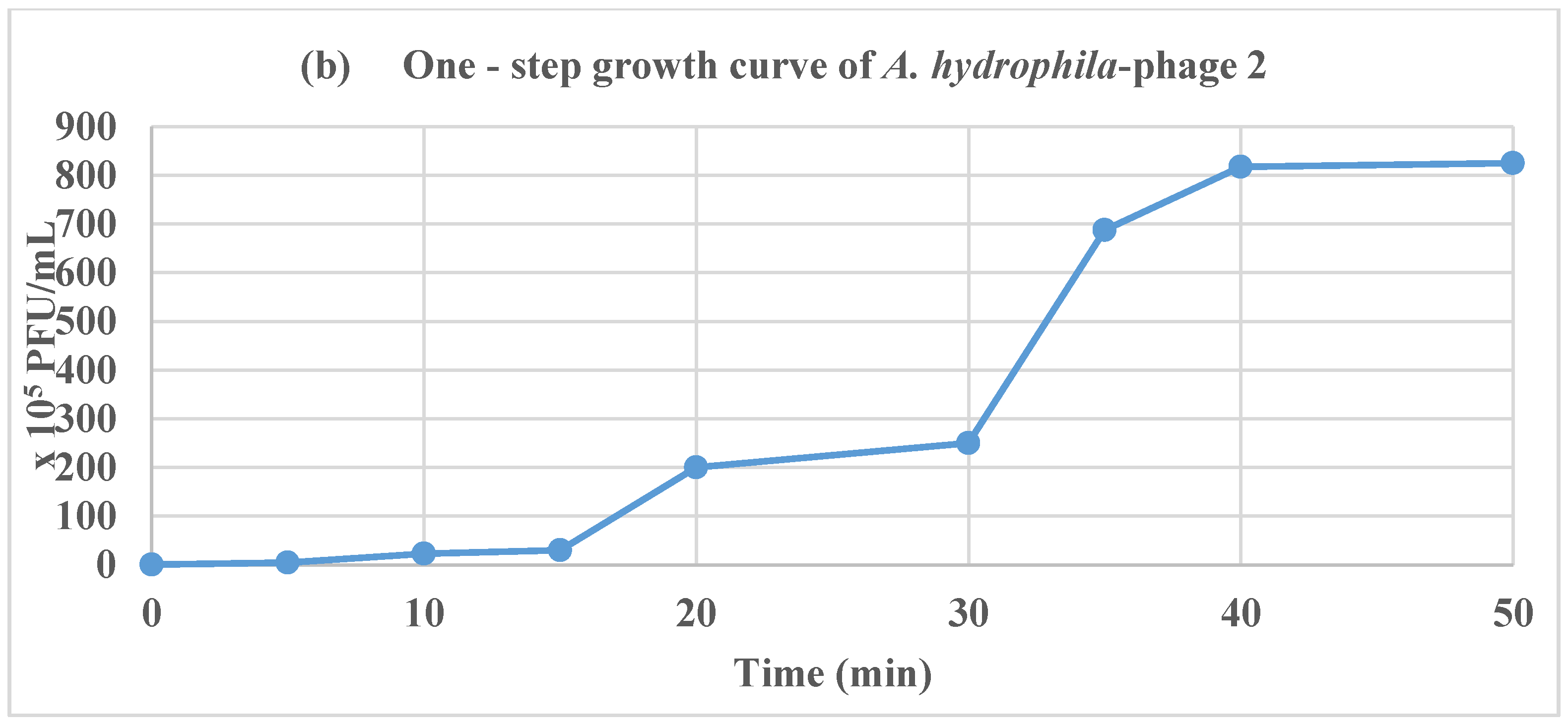
2.5. Adsorption Rate of Phages and One-Step Growth Curve
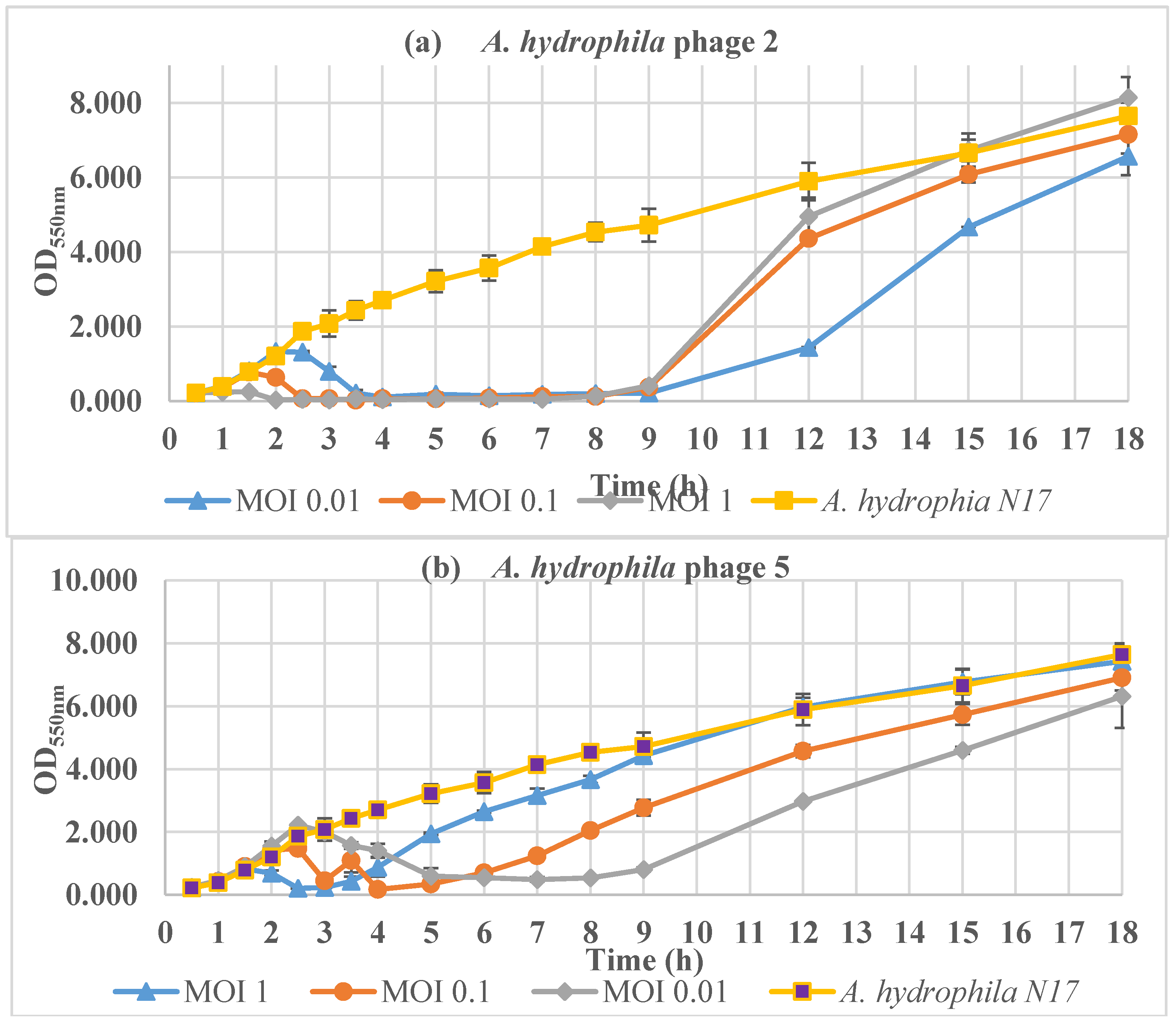
2.6. Inactivation of Aeromonas Species in Vitro
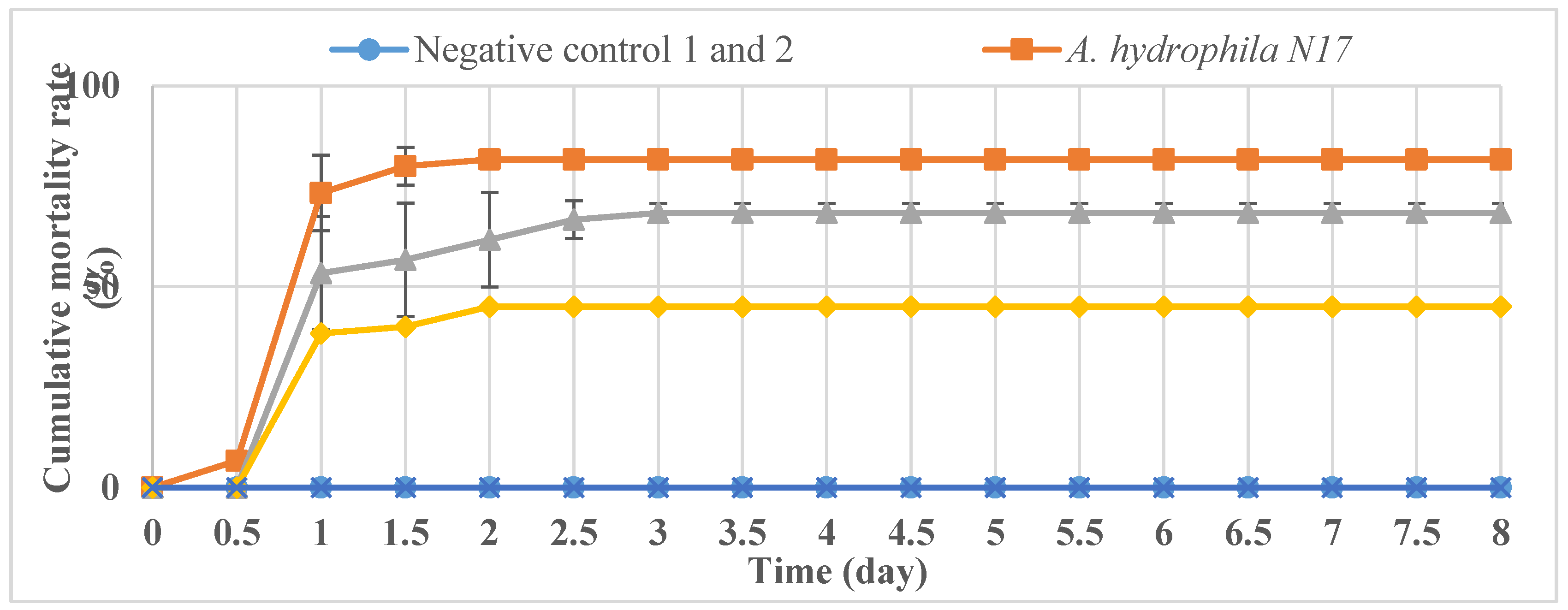
2.7. Phage Treatment of Infected Fish
3. Discussion
4. Materials and Methods
4.1. Aeromonas Species
4.2. Prophage Induction
4.3. Antibiotic Susceptibility
4.4. Isolation and Characterization of Bacteriophages
4.5. Host Range
4.6. Adsorption Rate of Phages
4.7. One-Step Growth Curve
4.8. Inactivation of Aeromonas hydrophila N17 in Vitro
4.9. Phage Treatment of Infected Fish
4.10. Statistical analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nguyen, N. Improving Sustainability of Striped Catfish (Pangasianodon hypophthalmus) Farming in the Mekong Delta, Vietnam through Recirculation Technology. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2016. [Google Scholar]
- Dung, T.; Ngoc, N.; Thinh, N.; Thy, D.; Tuan, N.; Shinn, A.; Crumlish, M. Common diseases of pangasius catfish farmed in Viet Nam. GAA 2008, 11, 77–78. [Google Scholar]
- Hien, N.T.; Lan, M.T.; Anh, P.V.N.; Phuong, V.H.; Loc, N.H.; Trong, C.Q.; Trung, C.T.; Phuoc, L.H. Report “Genetics of Aeromonas hydrophila on Catfish”. Research Institute for Aquaculture No. 2. 2014. Available online: http://www.sinhhoctomvang.vn/ban-tin/chi-tiet/Phat-hien-gen-gay-doc-cua-vi-khuan-Aeromonas-hydrophila-gay-benh-xuat-huyet-tren-ca-tra-106/ (accessed on 7 August 2017). (in Vietnamese).
- Vivekanandhan, G.; Savithamani, K.; Hatha, A.; Lakshmanaperumalsamy, P. Antibiotic resistance of aeromonas hydrophila isolated from marketed fish and prawn of south India. Int. J. Food Microbiol. 2002, 76, 165–168. [Google Scholar] [CrossRef]
- Guz, L.; Kozinska, A. Antibiotic susceptibility of Aeromonas hydrophila and A. sobria isolated from farmed carp (cyprinus carpio l.). Bull. Vet. Inst. Pulawy 2004, 48, 391–395. [Google Scholar]
- Thi, Q.V.C.; Dung, T.T.; Hiep, D.P.H. The current status antimicrobial resistance in Edwardsiella ictaluri and Aeromonas hydrophila cause disease on the striped catfish farmed in the mekong delta. Cantho Univ. J. Sci. 2014, 2, 7–14. (In vietnamese) [Google Scholar]
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. CAB Rev. 2012, 7, 1–16. [Google Scholar] [CrossRef]
- Hsu, C.-H.; Lo, C.-Y.; Liu, J.-K.; Lin, C.-S. Control of the eel (anguilla japonica) pathogens, Aeromonas hydrophila and Edwardsiella tarda, by bacteriophages. J. Fish. Soc. Taiwan 2000, 27, 21–31. [Google Scholar]
- El-Araby, D.; El-Didamony, G.; Megahed, M. New approach to use phage therapy against Aeromonas hydrophila induced Motile Aeromonas Septicemia in nile tilapia. J. Mar. Sci. Res. Dev. 2016, 6. [Google Scholar] [CrossRef]
- Ackermann, H.-W. 5500 phages examined in the electron microscope. Arch. Virol. 2007, 152, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Haq, I.U.; Chaudhry, W.N.; Andleeb, S.; Qadri, I. Isolation and partial characterization of a virulent bacteriophage ihq1 specific for Aeromonas punctata from stream water. Microb. Ecol. 2012, 63, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.W.; Kim, J.H.; Shin, S.P.; Han, J.E.; Chai, J.Y.; Park, S.C. Protective effects of the Aeromonas phages pah1-c and pah6-c against mass mortality of the cyprinid loach (misgurnus anguillicaudatus) caused by Aeromonas hydrophila. Aquaculture 2013, 416, 289–295. [Google Scholar] [CrossRef]
- Kim, J.; Son, J.; Choi, Y.; Choresca, C.; Shin, S.; Han, J.; Jun, J.; Kang, D.; Oh, C.; Heo, S. Isolation and characterization of a lytic Myoviridae bacteriophage pas-1 with broad infectivity in Aeromonas salmonicida. Curr. Microbiol. 2012, 64, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Anand, T.; Vaid, R.K.; Bera, B.C.; Singh, J.; Barua, S.; Virmani, N.; Yadav, N.K.; Nagar, D.; Singh, R.K.; Tripathi, B. Isolation of a lytic bacteriophage against virulent Aeromonas hydrophila from an organized equine farm. J. Basic Microb. 2016, 56, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Chow, M.S.; Rouf, M. Isolation and partial characterization of two Aeromonas hydrophila bacteriophages. Appl. Environ. Microbiol. 1983, 45, 1670–1676. [Google Scholar] [PubMed]
- Wu, J.-L.; Lin, H.-M.; Jan, L.; Hsu, Y.-L.; Chang, L.-H. Biological control of fish bacterial pathogen, Aeromonas hydrophila, by bacteriophage ah 1. Fish Pathol. 1981, 15, 271–276. [Google Scholar] [CrossRef]
- Fortier, L.-C.; Moineau, S. Morphological and genetic diversity of temperate phages in clostridium difficile. Appl. Environ. Microbiol. 2007, 73, 7358–7366. [Google Scholar] [CrossRef] [PubMed]
- Walakira, J.; Carrias, A.; Hossain, M.; Jones, E.; Terhune, J.; Liles, M. Identification and characterization of bacteriophages specific to the catfish pathogen, Edwardsiella ictaluri. J. Appl. Microbiol. 2008, 105, 2133–2142. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance standards for antimicrobial susceptibility testing 27th ed. CLSI supplement m100. Clinical and Laboratory Standards Institute. 2017. Available online: http://www.facm.ucl.ac.be/intranet/CLSI/CLSI-2017-M100-S27.pdf (accessed on 10 November 2017).
- Lamy, B.; Laurent, F.; Kodjo, A.; Roger, F.; Jumas-Bilak, E.; Marchandin, H.; Group, C.S. Which antibiotics and breakpoints should be used for Aeromonas susceptibility testing? Considerations from a comparison of agar dilution and disk diffusion methods using Enterobacteriaceae breakpoints. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2369–2377. [Google Scholar] [CrossRef] [PubMed]
- Daka, D.; Yihdego, D. Antibiotic-resistance Staphylococcus aureus isolated from cow’s milk in the hawassa area, south Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Cerveny, K.E.; DePaola, A.; Duckworth, D.H.; Gulig, P.A. Phage therapy of local and systemic disease caused by Vibrio vulnificus in iron-dextran-treated mice. Infect. Immun. 2002, 70, 6251–6262. [Google Scholar] [CrossRef] [PubMed]
- Le, T.S.; Southgate, P.C.; O’Connor, W.; Poole, S.; Kurtböke, D.I. Bacteriophages as biological control agents of enteric bacteria contaminating edible oysters. Curr. Microbiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Paterson, W.; Douglas, R.; Grinyer, I.; McDermott, L. Isolation and preliminary characterization of some Aeromonas salmonicida bacteriophages. J. Fish. Board Canada 1969, 26, 629–632. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; Dauguet, C.; Paterson, W.; Popoff, M.; Rouf, M.; Vieu, J.-F. Aeromonas bacteriophages: Reexamination and classification. Ann. Inst. Pasteur Virol. 1985, 136, 175–199. [Google Scholar] [CrossRef]
- Goodridge, L.; Gallaccio, A.; Griffiths, M.W. Morphological, host range, and genetic characterization of two coliphages. Appl. Environ. Microbiol. 2003, 69, 5364–5371. [Google Scholar] [CrossRef] [PubMed]
- Phumkhachorn, P.; Rattanachaikunsopon, P. Isolation and partial characterization of a bacteriophage infecting the shrimp pathogen Vibrio harveyi. Afr. J. Microbiol. Res. 2010, 4, 1794–1800. [Google Scholar]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. Methods Mol. Boil. 2009, 501, 175–202. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | Number of Resistant Isolates (n = 6) |
|---|---|
| Tetracycline | 2 |
| Oxytetracycline | 6 |
| Gentamycin | 6 |
| Kanamycin | 5 |
| Bactrim (SMX/TMP) | 6 |
| Doxycycline | 2 |
| Enrofloxacin | 6 |
| Amoxicillin/clavulanic acid | 6 |
| Ampicillin | 6 |
| Ciprofloxacin | 2 |
| Φ | Concentration PFU/mL | Head (nm) | Neck (nm) | Tail Sheath (nm) | Genus | |||
|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | |||
| 2 | 109 | 129 | 10 | 15 | 173 | 15 | Spounalikevirus | |
| 5 | 1010 | 120 | 15 | 15 | 198 | 15 | Spounalikevirus | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, T.S.; Nguyen, T.H.; Vo, H.P.; Doan, V.C.; Nguyen, H.L.; Tran, M.T.; Tran, T.T.; Southgate, P.C.; Kurtböke, D.İ. Protective Effects of Bacteriophages against Aeromonas hydrophila Causing Motile Aeromonas Septicemia (MAS) in Striped Catfish. Antibiotics 2018, 7, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics7010016
Le TS, Nguyen TH, Vo HP, Doan VC, Nguyen HL, Tran MT, Tran TT, Southgate PC, Kurtböke Dİ. Protective Effects of Bacteriophages against Aeromonas hydrophila Causing Motile Aeromonas Septicemia (MAS) in Striped Catfish. Antibiotics. 2018; 7(1):16. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics7010016
Chicago/Turabian StyleLe, Tuan Son, Thi Hien Nguyen, Hong Phuong Vo, Van Cuong Doan, Hong Loc Nguyen, Minh Trung Tran, Trong Tuan Tran, Paul C. Southgate, and D. İpek Kurtböke. 2018. "Protective Effects of Bacteriophages against Aeromonas hydrophila Causing Motile Aeromonas Septicemia (MAS) in Striped Catfish" Antibiotics 7, no. 1: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics7010016