Multiple Antibiotic Resistance (MAR), Plasmid Profiles, and DNA Polymorphisms among Vibrio vulnificus Isolates


Abstract
:1. Introduction
2. Results
2.1. Antibiotic Resistance
2.2. Multiple Antibiotic Resistance (MAR) Index
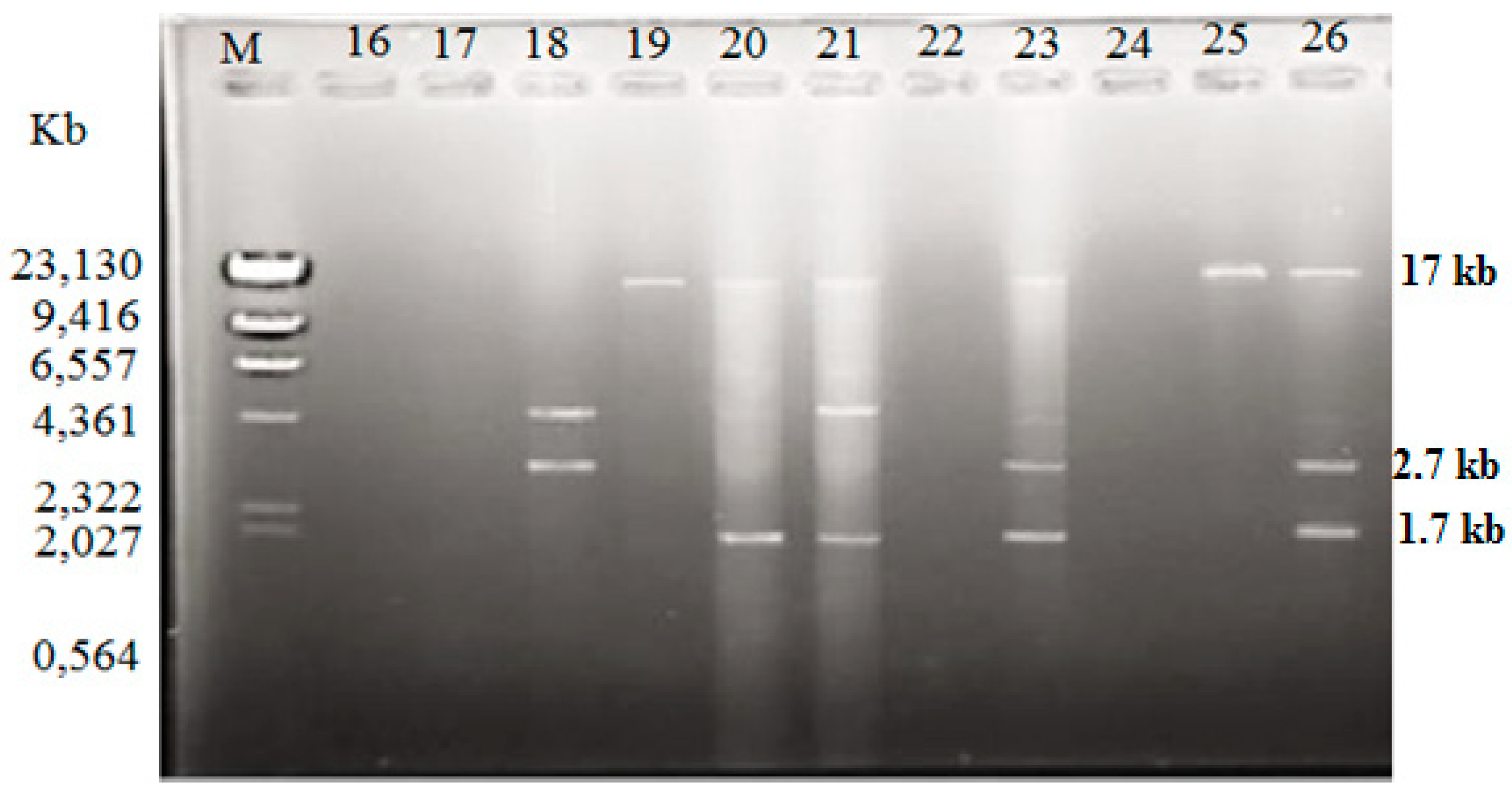
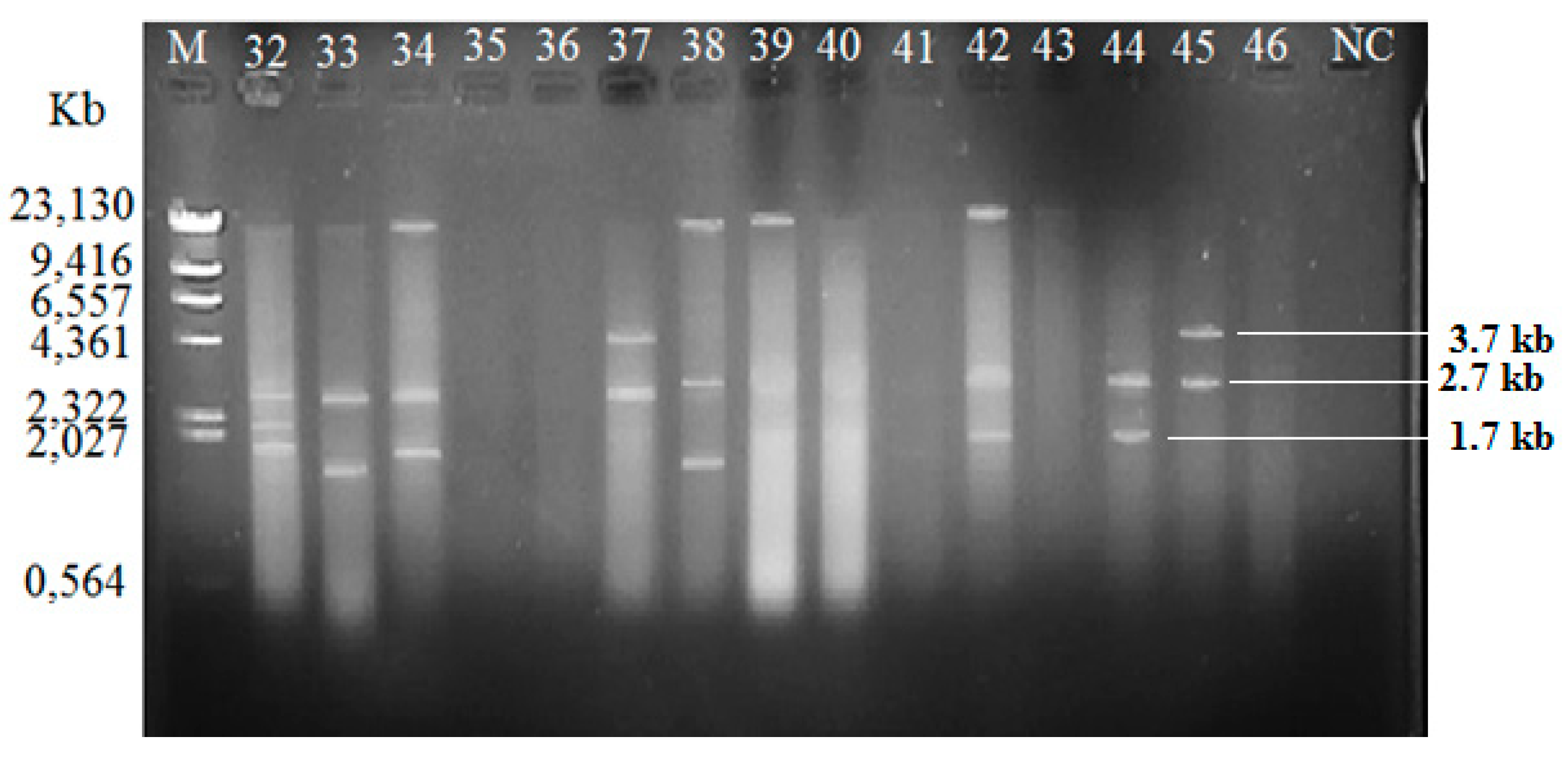
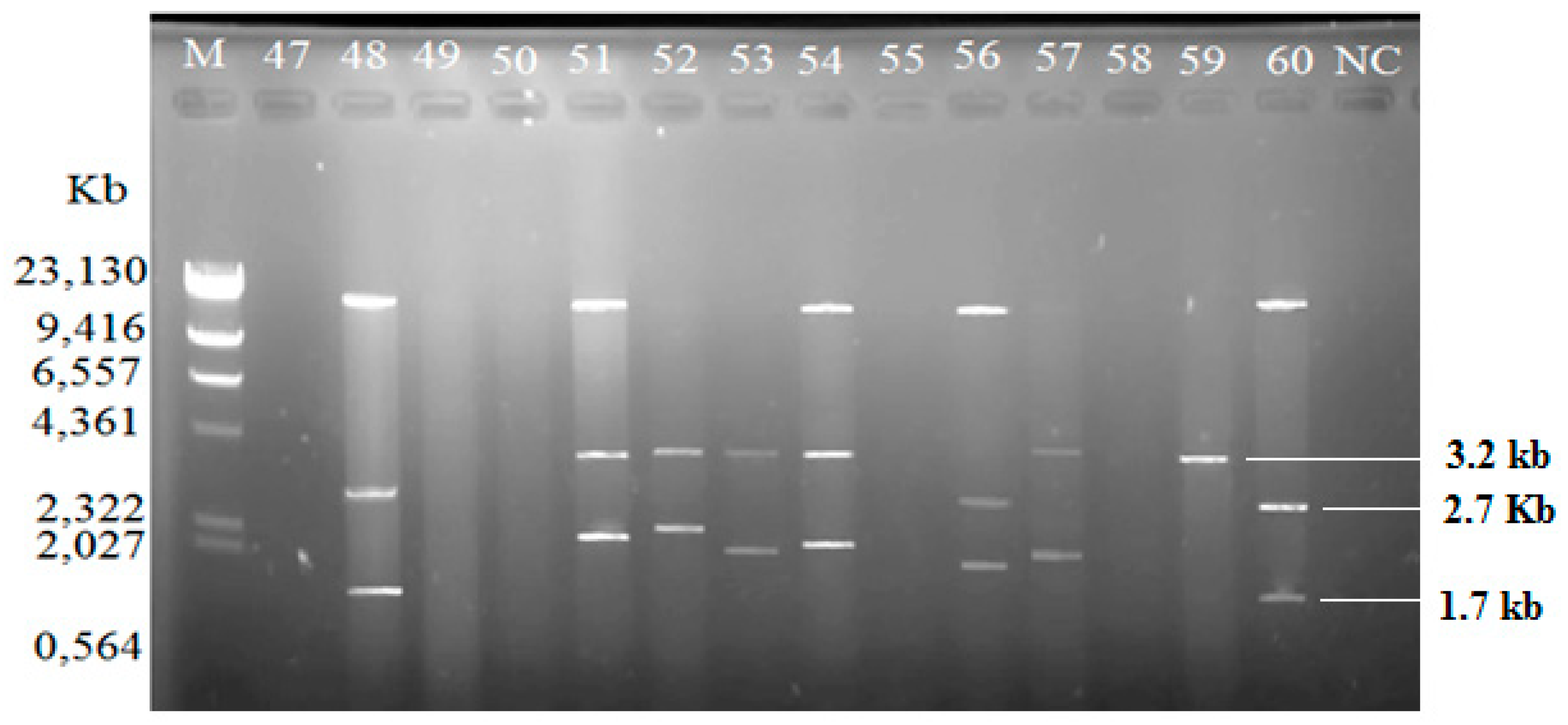
2.3. Plasmid Profiles
2.4. RAPD-PCR
3. Discussion
4. Materials and Methods
4.1. Vibrio vulnificus
4.2. Antibiotic Resistance
4.3. Multiple Antibiotic Resistance (MAR) Index
4.4. Plasmid Analysis
4.5. RAPD Fingerprinting
4.6. Phylogenetic Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miyasaka, J.; Yahiro, S.; Arahira, Y.; Tokunaga, H.; Katsuk, K.; Hara-Kudo, Y. Isolation of Vibrio parahaemolyticus and Vibrio vulnificus from wild aquatic birds in Japan. Epidemiol. Infect. 2006, 134, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.K.A.A.; Sahilah, A.M.; Ma’aruf, A.G.; Mohammed, A. A review of important virulence factors of Vibrio vulnificus. Curr. Res. J. Biol. Sci. 2014, 6, 76–88. [Google Scholar]
- Paydar, M.; Thong, K.L. Prevalence and genetic characterization of Vibrio vulnificus in raw seafood and seawater in Malaysia. J. Food Prot. 2013, 76, 1797–1800. [Google Scholar] [CrossRef] [PubMed]
- Sahilah, A.M.; Laila, R.A.; Sallehuddin, H.M.; Osman, H.; Aminah, A.; Ahmad Azuhairi, A. Antibiotic resistance and molecular typing among cockle (Anadara granosa) strains of Vibrio parahaemolyticus by polymerase chain reaction (PCR)-based analysis. World J. Microbiol. Biotechnol. 2014, 30, 649–659. [Google Scholar] [CrossRef]
- Antibiotic Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 25 January 2019).
- Srinivasan, P.; Ramasamy, P. Occurrence, distribution and antibiotic resistance patterns of Vibrio species associated with viral diseased shrimp of south Indian Aquaculture environment. Int. J. Agric. Sci. 2009, 1, 1–10. [Google Scholar]
- Lechumanan, V.; Yin, W.F.; Lee, L.H.; Chan, K.G. Prevelence and antimicrobial susceptibility of Vibrio parahaemolyticus o isolated from retail shrimps in Malaysia. Front. Microbiol. 2015, 6, 33. [Google Scholar]
- Lechumanan, V.; Pusparajah, P.; Tan, L.T.H.; Yin, W.F.; Lee, L.H.; Chan, K.G. Occurence and antibiotic resistance of Vibrio parahaemolyticus from shellfish in Selangor, Malaysia. Front. Microbiol. 2015, 6, 1417. [Google Scholar]
- Zulkifli, Y.; Alitheen, N.B.; Son, R.; Raha, A.R.; Samuel, L.; Yeap, S.K.; Nishibuchi, M. Random amplified polymorphic DNA-PCR and ERIC PCR analysis on Vibrio parahaemolyticus isolated from cockles in Padang, Indonesia. Int. Food Res. J. 2009, 16, 141–150. [Google Scholar]
- Roig, F.J.; Amaro, C. Plasmid diversity in Vibrio vulnificus biotypes. Microbiology 2009, 155, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Elhadi, N. Antibiotic Resistance and Plasmid Profiling of Clinically Significant Vibrio vulnificus Isolated from Coastal Water in Eastern Province of Saudi Arabia. Br. J. Pharm. Toxicol. 2012, 3, 93–97. [Google Scholar]
- Radu, S.; Elhadi, N.H.Z.; Rusul, G.; Lihan, S.; Fifadara, N.; Yuherman; Purwati, E. Characterization of Vibrio vulnificus isolated from cockles (Anadara granosa): Antimicrobial resistance, plasmid profiles and random amplification of polymorphic DNA analysis. FEMS Microbiol. Lett. 1998, 165, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Radu, S.; Vincent, M.; Apun, K.; Rahim, R.A.; Benjamin, P.G.; Yuherman; Rusul, G. Molecular characterization of Vibrio cholerae O1 outbreak strains in Miri, Sarawak (Malaysia). Acta Trop. 2002, 83, 168–176. [Google Scholar] [CrossRef]
- Okoh, A.I.; Igbinosa, E.O. Antibiotic susceptibility profiles of some Vibrio strains isolated from wastewater final effluents in a rural community of Eastern Cape Province of South Africa. BMC Microbiol. 2010, 10, 143. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental occurrence and clinical impact of Vibrio vulnificus and Vibrio parahaemolyticus: A European perspective. Environ. Microbiol. Rep. 2010, 2, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Tunung, R.; Jeyaletchumi, P.; Noorlis, A.; Tang, Y.H.; Sandra, A.; Ghazali, F.M.; Noranizan, M.A.; Lesley, M.B.; Haresh, K.K.; Nakaguchi, Y.; et al. Biosafety of Vibrio parahaemolyticus from vegetables based on antimicrobial sensitivity and RAPD profiling. Int. Food Res. J. 2012, 19, 467–474. [Google Scholar]
- Lee, S.W.; Wendy, W. Antibiogram and Heavy Metal Resistance Pattern of Salmonella spp. Isolated from Wild Asian Sea Bass (Lates calcarifer) from Tok Bali, Kelantan, Malaysia. Jordan J. Biol. Sci. 2011, 4, 125–128. [Google Scholar]
- Dale, J.W.; Park, S. Molecular Genetics of Bacteria, 1st ed.; John Wiley & Sons Inc.: Chichester, UK, 2010; pp. 147–148. [Google Scholar]
- Guglielgmetti, E.; Korhonen, J.M.; Haikkinen, J.; Morelli, L.; Von Wright, A. Transfer and plasmid-mediated resistance to tetracycline in pathogenic bacteria from fish and aquaculture environments. FEMS Microbiol. Lett. 2009, 293, 28–34. [Google Scholar] [CrossRef]
- Aoki, T.; Kitao, T.; Watanabe, S.; Takeshita, S. Drug resistance and R plasmids in Vibrio anguillarum isolated in cultured ayu (Plecoglossus altivelis). Microbiol. Immunol. 1984, 28, 1–9. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Gu, J.D. Identification of environmental plasmid-bearing Vibrio species isolated from polluted and pristine marine reserves of Hong Kong, and resistance to antibiotics and mercury. Antonic van Leeuwenhoek 2006, 89, 307–315. [Google Scholar] [CrossRef]
- Zhang, R.; Pan, L.; Zhao, Z. High incidence of plasmids in marine Vibrio species isolated from Mai Po Nature Reserve of Hong Kong. Ecotoxicology 2012, 21, 1661–1668. [Google Scholar] [CrossRef] [Green Version]
- Sahilah, A.M.; Audrey, L.Y.Y.; Ong, S.L.; Wan Sakeenah, W.N.; Safiyyah, S.; Norrakiah, A.S.; Aminah, A.; Ahmad Azuhairi, A. DNA profiling among egg and beaf meat isolates of Escherichia coli by enterobacterial repetitive intergenic consensus-PCR (ERIC-PCR) and random amplified polymorphic DNA-PCR (RAPD-PCR). Int. Food Res. J. 2010, 17, 853–866. [Google Scholar]
- Nisreen, J.K.; Sahilah, A.M. Genetic diversity of Bacillus cereus isolated from fried rice. Int. J. ChemTech Res. 2018, 11, 52–59. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turl, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Sahilah, A.M.; Son, R.; Rusul, G.; Samuel, L.; Hassan, Z.; Lum, K.Y.; Ahmad, M.A. Molecular typing of Salmonella weltevreden and Salmonella chincol by pulsed field gel electrophoresis (PFGE) and enterobacterial repetitive intergenic consensuspolymerase chain reaction (ERIC-PCR). World J. Microbiol. Biotechnol. 2000, 16, 621–624. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Tested | Percentage of Resistance (%) | ||
|---|---|---|---|
| Cockle Samples, Malaysia (n = 37) | Clam Samples, Qatar (n = 23) | Mean Average | |
| Ampicillin | 65 | 74 | 70 |
| Bacitracin | 59 | 52 | 56 |
| Cefoperazone | 0 | 0 | 0 |
| Cephalothin | 73 | 57 | 65 |
| Clindamycin | 70 | 61 | 66 |
| Erythromycin | 57 | 45 | 51 |
| Kanamycin | 0 | 13 | 6 |
| Novobiocin | 57 | 35 | 46 |
| Penicillin | 95 | 91 | 93 |
| Streptomycin | 0 | 0 | 0 |
| Tetracycline | 0 | 0 | 0 |
| Vancomycin | 62 | 65 | 64 |
| Strain No. a | Locations | MAR Index | Antibiotic Patterns b | Plasmid Profiles c | RAPD | Strain Types d | ||
|---|---|---|---|---|---|---|---|---|
| RAPD 11 | RAPD 15 | |||||||
| 1 | Malaysia | 0.53 | BDaEKfNvPVa | A1 | P1 | C1 | F1 | 1 |
| 2 | Malaysia | 0.53 | BDaEKfNvPVa | A1 | P2 | C1 | F1 | 2 |
| 3 | Malaysia | 0.5 | AmpDaEKfNvP | A2 | P2 | C2 | F2 | 3 |
| 4 | Malaysia | 0.53 | BDaEKfNvPVa | A1 | P2 | C3 | F3 | 4 |
| 5 | Malaysia | 0.25 | AmpKfP | A3 | P2 | C4 | F4 | 5 |
| 6 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | P3 | C4 | F5 | 6 |
| 7 | Malaysia | 0.5 | BDaKfNvPVa | A5 | P4 | C5 | F6 | 7 |
| 8 | Malaysia | 0.5 | BDaKfNvPVa | A5 | - | C6 | F7 | 8 |
| 9 | Malaysia | 0.53 | AmpBDaKfNvPVa | A6 | - | C6 | F8 | 9 |
| 10 | Malaysia | 0.25 | AmpKfP | A3 | P5 | C7 | F9 | 10 |
| 11 | Malaysia | 0.35 | AmpKfP | A3 | P3 | C8 | F10 | 11 |
| 12 | Malaysia | 0.16 | PVa | A7 | P1 | C8 | F11 | 12 |
| 13 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | P6 | C9 | F12 | 13 |
| 14 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | - | C10 | F13 | 14 |
| 15 | Malaysia | 0.5 | AmpDaEKfNvP | A2 | - | C11 | F14 | 15 |
| 16 | Malaysia | 0.25 | AmpKfP | A3 | - | C12 | F15 | 16 |
| 17 | Malaysia | 0.25 | AmpKfP | A3 | - | C13 | F16 | 17 |
| 18 | Malaysia | 0.58 | AmpBDaKfNvPVa | A6 | P7 | C14 | F17 | 18 |
| 19 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | P8 | C15 | F18 | 19 |
| 20 | Malaysia | 0.25 | KfPVa | A8 | P9 | C15 | F19 | 20 |
| 21 | Malaysia | 0.58 | AmpBDaKfNvPVa | A6 | P10 | C16 | F20 | 21 |
| 22 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | - | C16 | F20 | 22 |
| 23 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | P2 | C16 | F21 | 23 |
| 24 | Malaysia | 0.25 | AmpPVa | A12 | - | C16 | F21 | 24 |
| 25 | Malaysia | 0.25 | AmKfP | A3 | P8 | C17 | F22 | 25 |
| 26 | Malaysia | 0.33 | BDaEP | A9 | P2 | C18 | F23 | 26 |
| 27 | Malaysia | 0.5 | BDaENvPVa | A10 | P3 | C19 | F24 | 27 |
| 28 | Malaysia | 0.33 | AmpBDaE | A11 | - | UT | F25 | 28 |
| 29 | Malaysia | 0.66 | AmpBDaEKfNvPVa | A4 | P3 | C20 | F25 | 29 |
| 30 | Malaysia | 0.33 | BDaEP | A9 | P3 | UT | F26 | 30 |
| 31 | Malaysia | 0.25 | AmpPVa | A12 | P1 | C21 | F27 | 31 |
| 32 | Malaysia | 0.5 | AmpDaEKfNvP | A2 | P11 | C22 | F28 | 32 |
| 33 | Malaysia | 0.5 | BDaENvPVa | A10 | P3 | C23 | F29 | 33 |
| 34 | Malaysia | 0.16 | KfVa | A13 | P3 | C24 | F29 | 34 |
| 35 | Malaysia | 0.5 | AmpBDaEPVa | A14 | - | C25 | F30 | 35 |
| 36 | Malaysia | 0.33 | BDaEP | A9 | - | C26 | F31 | 36 |
| 37 | Malaysia | 0.5 | AmpDaEKfNvP | A2 | P7 | C27 | F32 | 37 |
| 38 | Qatar | 0.66 | AmpBDaEKKfNvP | A15 | P12 | C28 | F33 | 38 |
| 39 | Qatar | 0.41 | AmpKNvPVa | A16 | P13 | C28 | F34 | 39 |
| 40 | Qatar | 0.5 | AmpDaEKfNvP | A2 | - | C29 | F35 | 40 |
| 41 | Qatar | 0.41 | AmpDaKPVa | A17 | - | C30 | F36 | 41 |
| 42 | Qatar | 0.25 | AmpPVa | A12 | P14 | C31 | F37 | 42 |
| 43 | Qatar | 0.5 | AmpBDaEPVa | A14 | - | C32 | F38 | 43 |
| 44 | Qatar | 0.25 | AmpPVa | A12 | P3 | C34 | F40 | 44 |
| 45 | Qatar | 0.5 | AmpBDaEPVa | A14 | P6 | C35 | F41 | 45 |
| 46 | Qatar | 0.25 | AmpKfP | A3 | - | C16 | F21 | 46 |
| 47 | Qatar | 0.58 | AmpBDaKfNvPVa | A6 | - | C36 | F42 | 47 |
| 48 | Qatar | 0.41 | AmpBDaEP | A18 | P15 | C36 | F43 | 48 |
| 49 | Qatar | 0.25 | AmpKfP | A3 | - | C37 | F43 | 49 |
| 50 | Qatar | 0.5 | AmpBDaEPVa | A14 | - | C38 | F44 | 50 |
| 51 | Qatar | 0.58 | BDaEKfNvPVa | A1 | P16 | C39 | F45 | 51 |
| 52 | Qatar | 0.58 | BDaEKfNvPVa | A1 | P17 | C40 | F46 | 52 |
| 53 | Qatar | 0.58 | BDaEKfNvPVa | A1 | P18 | C41 | F47 | 53 |
| 54 | Qatar | 0.5 | AmpBDaEPVa | A14 | P16 | C42 | F48 | 54 |
| 55 | Qatar | 0.5 | AmpBDaEPVa | A14 | - | C43 | UT | 55 |
| 56 | Qatar | 0.25 | AmpKfP | A3 | P17 | C44 | F49 | 56 |
| 57 | Qatar | 0.25 | AmpKfP | A3 | P18 | C45 | F50 | 57 |
| 58 | Qatar | 0.16 | KfVa | A13 | - | ND | F50 | 58 |
| 59 | Qatar | 0.16 | KfVa | A13 | P19 | C46 | F50 | 59 |
| 60 | Qatar | 0.58 | BDaEKfNvPVa | A1 | P15 | C47 | F50 | 60 |
| MAR Index | Percentage of Occurrence (%) | |
|---|---|---|
| Isolates from Malaysia (Cockles) (n = 37) | Isolates from Qatar (Clams) (n = 23) | |
| 0.1 | - | - |
| 0.2 | 22 | 32 |
| 0.3 | 16 | - |
| 0.4 | 3 | 13 |
| 0.5 | 35 | 30 |
| 0.6 | 5 | 17 |
| 0.7 | 19 | 4 |
| 0.8 | - | - |
| 0.9 | - | - |
| 1.0 | - | - |
| Antibiotic Tested | Mean Averages (%) Isolates Resistant within Two Sample Sources | No. of V. vulnificus Isolates Resistant | |
|---|---|---|---|
| With Plasmid (40) | Without Plasmid (20) | ||
| Ampicillin | 70 | 36 (90%) | 15 (75%) |
| Bacitracin | 56 | 23 (58%) | 11 (55%) |
| Cefoperazone | 0 | 0 | 0 |
| Cephalothin | 65 | 28 (70%) | 12 (60%) |
| Clindamycin | 66 | 27 (68%) | 13 (65%) |
| Erythromycin | 51 | 22 (55%) | 9 (45%) |
| Kanamycin | 6 | 2 (5%) | 1 (5%) |
| Novobiocin | 46 | 23 (58%) | 6 (30%) |
| Penicillin | 93 | 39 (98%) | 17 (85%) |
| Streptomycin | 0 | 0 | 0 |
| Tetracycline | 0 | 0 | 0 |
| Vancomycin | 64 | 27 (68%) | 11 (55%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
M. Kurdi Al-Dulaimi, M.; Abd. Mutalib, S.; Abd. Ghani, M.; Mohd. Zaini, N.A.; Ariffin, A.A. Multiple Antibiotic Resistance (MAR), Plasmid Profiles, and DNA Polymorphisms among Vibrio vulnificus Isolates. Antibiotics 2019, 8, 68. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020068
M. Kurdi Al-Dulaimi M, Abd. Mutalib S, Abd. Ghani M, Mohd. Zaini NA, Ariffin AA. Multiple Antibiotic Resistance (MAR), Plasmid Profiles, and DNA Polymorphisms among Vibrio vulnificus Isolates. Antibiotics. 2019; 8(2):68. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020068
Chicago/Turabian StyleM. Kurdi Al-Dulaimi, Mohammed, Sahilah Abd. Mutalib, Maaruf Abd. Ghani, Nurul Aqilah Mohd. Zaini, and Ahmad Azuhairi Ariffin. 2019. "Multiple Antibiotic Resistance (MAR), Plasmid Profiles, and DNA Polymorphisms among Vibrio vulnificus Isolates" Antibiotics 8, no. 2: 68. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020068