Regulation of Geldanamycin Biosynthesis by Cluster-Situated Transcription Factors and the Master Regulator PhoP

1
Area de Microbiología, Departmento de Biología Molecular, Universidad de León, 24071 León, Spain
2
Instituto de Biotecnología (INBIOTEC). Av. Real 1, 24006 León, Spain
*
Author to whom correspondence should be addressed.
Antibiotics 2019, 8(3), 87; https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030087
Submission received: 5 June 2019
/
Revised: 25 June 2019
/
Accepted: 29 June 2019
/
Published: 30 June 2019
(This article belongs to the Special Issue Mechanism and Regulation of Antibiotic Synthesis in Streptomyces)
Abstract
:Geldanamycin and the closely related herbimycins A, B, and C are benzoquinone-type ansamycins with antitumoral activity. They are produced by Streptomyces hygroscopicus var. geldanus, Streptomyces lydicus and Streptomyces autolyticus among other Streptomyces strains. Geldanamycins interact with the Hsp-90 chaperone, a protein that has a key role in tumorigenesis of human cells. Geldanamycin is a polyketide antibiotic and the polyketide synthase contain seven modules organized in three geldanamycin synthases genes named gdmAI, gdmAII, and gdmAIII. The loading domain of GdmI activates AHBA, and also related hydroxybenzoic acid derivatives, forming geldanamycin analogues. Three regulatory genes, gdmRI, gdmRII, and gdmRIII were found associated with the geldanamycin gene cluster in S. hygroscopicus strains. GdmRI and GdmRII are LAL-type (large ATP binding regulators of the LuxR family) transcriptional regulators, while GdmRIII belongs to the TetR-family. All three are positive regulators of geldanamycin biosynthesis and are strictly required for expression of the geldanamycin polyketide synthases. In S. autolyticus the gdmRIII regulates geldanamycin biosynthesis and also expression of genes in the elaiophylin gene cluster, an unrelated macrodiolide antibiotic. The biosynthesis of geldanamycin is very sensitive to the inorganic phosphate concentration in the medium. This regulation is exerted through the two components system PhoR-PhoP. The phoRP genes of S. hygroscopicus are linked to phoU encoding a transcriptional modulator. The phoP gene was deleted in S. hygroscopicus var geldanus and the mutant was unable to grow in SPG medium unless supplemented with 5 mM phosphate. Also, the S. hygroscopicus pstS gene involved in the high affinity phosphate transport was cloned, and PhoP binding sequences (PHO boxes), were found upstream of phoU, phoRP, and pstS; the phoRP-phoU sequences were confirmed by EMSA and nuclease footprinting protection assays. The PhoP binding sequence consists of 11 nucleotide direct repeat units that are similar to those found in S. coelicolor Streptomyces avermitilis and other Streptomyces species. The available genetic information provides interesting tools for modification of the biosynthetic and regulatory mechanisms in order to increase geldanamycin production and to obtain new geldanamycin analogues with better antitumor properties.
1. Introduction: Antitumor Activity of Geldanamycin and its Derivatives
Geldanamycin and the closely related herbimycins A, B, and C are benzoquinone-type ansamycins [1] with antitumoral activity [2,3] (Figure 1A). These compounds are produced by different strains of S. hygroscopicus, Streptomyces lydicus, and by Streptomyces autolyticus, and gene clusters for geldanamycin have been found in other Streptomyces strains (Table 1). Both, geldanamycin and the herbimycins were discovered as weak antifungal and antibacterial antibiotics [4,5,6] but their major interest is as potent antitumor agents due to their ability to interact with the Hsp-90 chaperone complex in human cells [7,8]. The members of the Hsp-90 chaperone family play an important role in the tumorigenesis process in humans. Both geldanamycin and the herbimycins have potent antitumor activity at nanomolar concentration, particularly the former [9,10]. However, it was shown that the natural compounds are hepatotoxic [11]. In the last decades, great interest has focused on the discovery of new derivatives with lower toxicity by direct genetic modification of the known geldanamycin gene cluster and investigation on new producer strains [3,12,13,14]. Structurally similar compounds, such as 17-amino-17-demethoxy-geldanamycin, were found in a knock out mutant of S. autolyticus CGMCC 0516 [15]. Two other chemical derivatives of geldanamycin, 17-allylamino-17-demethoxygeldanamycin, and 17-(2-dimethylamino) ethylamino-17-demethoxy-geldanamycin have been tested in clinical trials [16,17].
2. Biosynthesis of Geldanamycins
Geldanamycin is a polyketide derived ansamycin [4] that is synthesized from precursors assembled by polyketide synthases. In general, the geldanamycin biosynthetic process can be divided in three large steps: (1) Biosynthesis of the precursor 3-amino-5-hydroxybenzoic acid (AHBA); (2) extension of the starter unit with polyketide elongation units, and (3) post-polyketide modifications [19].
2.1. Origin and Biosynthesis of the AHBA Unit
All ansamycins derive from a seven carbon and one nitrogen atom (mC7N) AHBA starter unit, that is elongated using malonyl-CoA or methylmalonyl-CoA units and finally form the ansamycin ring through a lactam bond. The AHBA unit is synthesized from glucose by the amino shikimate pathway which includes the three initial steps similar to those of the shikimate pathway (for the biosynthesis of aromatic amino acids). Previous information on the synthesis of the AHBA unit of rifamycin provides evidence indicating that seven genes, rifGHJKLMN, linked in a cluster, are required and sufficient to synthesize this compound in Amycolatopsis mediterranei. The gene rifK of this cluster, that encodes the AHBA synthase, has been extensively used to search for ansamycin gene clusters in other actinobacteria [25,26,27,28]. Homologs of these genes were searched in S. hygroscopicus 17997, producer of geldanamycin, and two different gene clusters were found. One of them, of the benzoquinone type, was shown to be involved in geldanamycin biosynthesis [18]. Genes of the second cluster, of the naphtoquinone type, did not complement mutants in geldanamycin biosynthesis in S. hygroscopicus [19].
The work of He et al. [19] provides evidence showing that the benzoquinone-type cluster in the S. hygroscopicus strain is, indeed, involved in geldanamycin biosynthesis while the naphtoquinone-type cluster is likely to be involved in biosynthesis of a rifamycin-type ansamycin. Similarly, two gene clusters encoding benzoquinone-type and naphtoquinone-type ansamycins have been found in Streptomyces collinus that produces ansatrienin and naphthomycin [26]. The presence of two related gene clusters in single Streptomyces species is relatively common. This is normally generated by gene duplication and subsequent specialization. Interestingly, there are differences in the organization of the AHBA gene cluster in the original S. hygroscopicus NRRL 3602 [18] and S. hygroscopicus 17997 [19]. This indicates that these two strains have evolved separately in modern times, although most likely both AHBA gene clusters derive from a common cluster in an ancestor of both strains.
2.2. Elongation Steps
After formation of the AHBA starter unit the biosynthesis of geldanamycin proceeds by elongation steps involving one malonyl-CoA unit, 4 methylmalonyl-CoA units, and 2 methoxy-malonyl-ACP elongation units. These elongation reactions are catalyzed by three polyketide synthases encoded by gdmAI, gdmAII, and gdmAIII, totaling seven modules. They are organized in three polyketide synthases (Figure 2, Table 2), containing the AT, KS, ACP, and KR domains. The first condensation step catalyzed by a PK type I is of interest, since it includes a loading domain in addition to the ACP domain of the PKS. The loading domain of this PKS activates AHBA and related aromatic units [29] by a mechanism that uses ATP and is similar to that performed by the A domain of NRPSs [30,31]. In this reaction the carboxyl group of the AHBA is activated as an acyl-phosphate and then is transferred to the ACP1 in the first PKS synthase. This activation and elongation is similar to the p-aminobenzoic acid (PABA) activation and elongation performed by the first module of the candicidin PKS [32]. Noteworthy, the loading domain has low substrate specificity and is able to activate several benzoic acid related compounds. This activation and first elongation are also very similar to those performed by the rifamycin producer LM-M1 bimodule of A. mediterranei. In this strain the protein encoded by the first PKS, named rifA, served as a model for the biosynthesis of rifamycin and other ansamycins in different Streptomyces; it contains an initial didomain consisting of an AHBA activating A domain and an ACP domain. This didomain, separated from the rest of the PKS protein, has been expressed in E. coli and shown to be able to activate AHBA and related benzoic acid derivatives [29]. The requirement of ATP and CoA in the in vitro (E. coli extracts) reaction excludes the possibility that the starter unit is provided to the PKS in an activated different form during its synthesis in the shikimate pathway. This loading enzyme is able to activate, in addition to the natural starter unit, other 3- or 5-substituted benzoic acid derivatives and 3,5-disubstituted derivatives. Therefore, as occurs in the case of rifamycin, the loading module in the GdmA PKS of the geldanamycin gene cluster has the potential to synthesize compounds similar to geldanamycin containing aromatic units related to AHBA. The subsequent elongation steps use the standard polyketide biosynthesis mechanisms.
Transcriptional studies of the geldanamycin polyketide synthase genes in S. hygroscopicus XM201 suggested that expression of the polyketide synthase genes was limiting for geldanamycin biosynthesis [21]. These authors replaced the native polyketide synthase promoter upstream of gdmA1 by a strong endogenous S. hygroscopicus XM201 promoter selected on the basis of its high transcriptional activity. Interestingly, the replacement of the native promoter led to overexpression of the gdmA genes (4 to 141-fold) leading to 39% increase of geldanamycin production. Then, in this overproducing strain, biosynthesis of the AHBA starter unit became limiting. Combined overexpression of the polyketide synthase gene and the amino shikimate gene cluster resulted in an increase of 88% in geldanamycin production [21].
2.3. Cyclization of the Lineal Polyketide to Progeldanamycin
After completion of the growing polyketide chain, it is released from the enzyme by intramolecular cyclization catalyzed by the amide synthase encoded by gdmF, a gene present in all geldanamycin producers (Figure 2, Table 2). It is noteworthy that the amide synthase of the geldanamycin gene cluster of S. hygroscopicus has much broader substrate specificity than the homologous enzyme of Actinnosynnema pretiosum [33], that produces ansamitocin, a cytotoxic compound. These authors isolated mutants of each of these two microorganisms blocked in the synthesis of the AHBA starting unit, to test whether they are able to incorporate other hydroxybenzoic acid compounds (which lacks the amino group of AHBA). Using these mutants, they demonstrated that at the difference of the A. pretiosum amide synthase, the enzyme of S. hygroscopicus was able to cyclize the linear polyketide intermediate containing hydroxybenzoic acid, forming a 20 membered lactone ring instead of the standard 19 membered lactam ring of geldanamycin. The difference of the amide synthases in these two organisms is apparently due to the protein structure that determines the accessibility of the substrate to the active center in the enzyme, formed by the triad Cys72, His110, and Asp125.
The incorporation of AHBA starter analogous units by S. hygroscopicus produced compounds similar to geldanamycin with potential antitumor activity [33]. However, none of them improved the antitumor activity of the native geldanamycin.
2.4. Post-Polyketide Modifications
The pro-geldanamycin intermediate is finally modified by a series of polyketide modifications that include hydroxylation at C-17 and oxidations at C-18 and C-21, followed by methylation of the hydroxyl group at C-17, introduction of the carbamoyl group at C-7 and finally formation of the double bond between C-4 and C-5. The C-17 hydroxylation and the C-18 and C-21 oxidations have remained unclear for many years; these reactions might be performed by at least one flavin-dependent oxidase, encoded by gdmM [15]. The O-methyltransferase that introduces a methyl group at the hydroxyl formed at C-17 was not initially found in the geldanamycin gene cluster but later Yin et al. [15] found a putative methyltransferase gene, gdmMT, located 17 kb away from the main geldanamycin gene cluster in S. autolyticus. The involvement of the gdmMT gene in the methylation of the hydroxyl group at C-17 was supported by in vivo and in vitro experiments. The O-carbamoyl transferase encoding gene, gdmN, cloned from S. hygroscopicus 17997 is very similar to the orthologous genes of other geldanamycin producers [34]. The gdmN-disrupted mutant synthesized C-7 decarbamoyl-geldanamycin confirming that the enzyme encoded by this gene is involved in the carbamoylation at C-7. Furthermore, these authors complemented the disrupted mutant with the wild type gdmN gene resulting in the formation of geldanamycin. Interestingly, the gdmN disrupted mutant was able to synthesize several novel geldanamycin compounds, including 4,5-dihydro-7-O-decarbamoyl-7-hydroxy-19-O-glycylgeldanamycin and 4,5-dihydro-7-O-decarbamoyl-7 hydroxygeldanamycin. These novel compounds showed less activity against human tumor cell lines but instead they showed better solubility [34]. Finally, desaturation resulting in the formation of the C-4/C-5 double bond was catalyszd by a P450 monooxygenase encoded by gdmP (see below).
3. Geldanamycin Gene Clusters in Different Streptomyces Species
Similar geldanamycin gene clusters have been found in other Streptomyces species (Table 1). Recent studies using different geldanamycin producers, as S. hygroscopicus sub. duamyceticus JCM4427 [20], have shed light on the post-polyketide modification reactions, although the use of different names for the same gene makes it difficult to compare the findings of different authors.
Two of the genes, named gel1 and gel7 by these authors (equivalent to gdmL and gdmM, respectively), encode flavin dependent oxygenases. Disruption of gel1 did not affect geldanamycin production, indicating that gel1 has no role in geldanamycin biosynthesis, whereas disruption of gel7 led to the formation of 17-demethoxy-reblastatin, an ansamycin containing a benzoquinone nucleus, suggesting that gel7 is involved in an oxidation of the benzoquinone ring. Complementation of the gel7 disrupted mutant with the wild type gel7 gene re-established the geldanamycin production. Shin et al. [20] proved that the hydroxylations at C-17 and C-21 are previous to the C-7 O-carbamoylation. They also described that gel16 (equivalent to gdmP), that is linked to a ferredoxin gene, encodes a P450 oxygenase, containing an heme-pocket. Disruption of the gel16 gene results in the formation of 4,5 dihydrogeldanamycin, and geldanamycin production was restored by complementation with a wild type gel16 gene suggesting that gel16 is involved in the formation of the geldanamycin C-4/5 double-bond. This was formally demonstrated by Rimal et al. [37] obtaining recombinant Gel-16 protein in E. coli and using it to perform the in vitro conversion of 4,5-dihydrogeldanamycin to geldanamycin. In silico protein–protein docking studies were performed by these authors to identify putative ferredoxin and ferredoxin reductases electron transporters cooperating with Gel16, involved in the dehydrogenation step. Although no hydroxylated intermediates were found in the gel16- disrupted mutant, the authors do not exclude that a hydroxylated intermediate is involved in the process of the double-bond formation.
In another geldanamycin producer, S. autolyticus JX-47, the complete genome consists of a 10 Mb lineal chromosome and 7 circular plasmids [22]. This strain was obtained from a laboratory at Yunnan (China) as an autolytimycin producing strain. Later, it was shown that this strain also produced geldanamycin, and a geldanamycin gene cluster was isolated in a bacterial artificial chromosome (BAC) library [22]. From this library, a 250 kb contiguous region of S. autolyticus JX-47 DNA, that included the complete geldanamycin cluster, was subcloned. In this strain, the central polyketide region of the geldanamycin gene cluster is 99% identical to that of the S. hygroscopicus 17997 strain and the encoded proteins range from 81% to 100% identity in amino acid sequence to the orthologous proteins of S. hygroscopicus. All genes of the geldanamycin cluster in S. autolyticus, have similar organization to those of S. hygroscopicus NRRL3602, except gdmL and gdmX, which are absent from the core region (Figure 2, Table 2).
4. Regulatory Genes in the Geldanamycin Gene Cluster
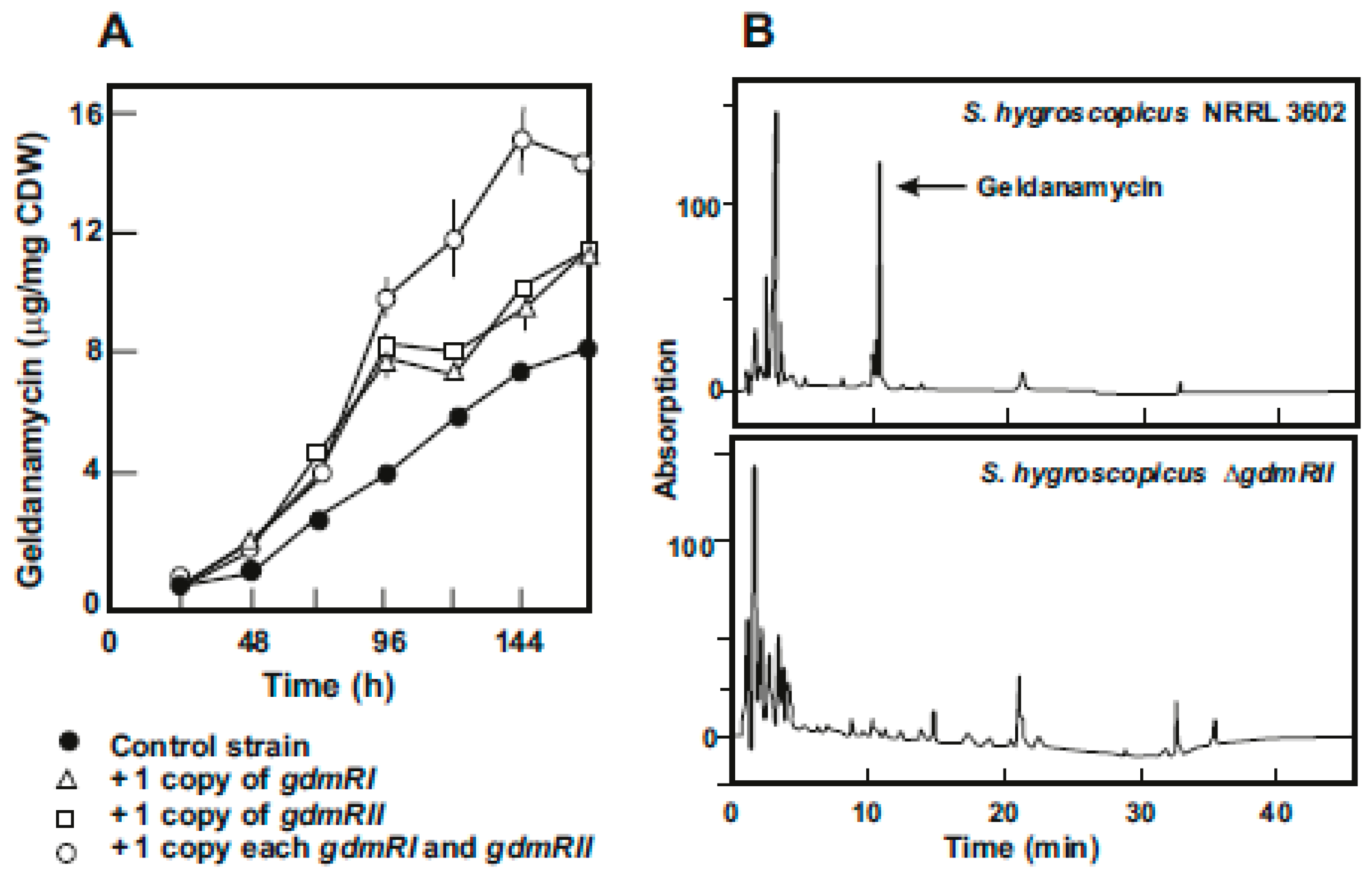
In the early studies on geldanamycin biosynthesis, Rascher et al. [18] found that in the geldanamycin gene cluster of S. hygroscopicus var. geldanus there were several putative regulatory genes. A similar finding was reported later in the cluster of S. hygroscopicus 17997 [35]. These authors cloned and characterized two geldanamycin regulatory genes, named gdmRI and gdmRII; both genes encode LAL type regulators which contain a Walker motif (ATP/GTP binding site) in the amino terminal region and a helix-turn-helix binding domain in the carboxyl end. Both GdmRI and GdmRII act as positive regulators of the geldanamycin biosynthesis as shown by gene disruption and complementation studies. In an independent parallel work, we isolated a mutant of S. hygroscopicus var. geldanus NRRL3602 deleted in gdmRII and observed that this mutant was completely blocked in geldanamycin biosynthesis (Figure 3).
Both, the results of He et al. [35] and our own experiments, indicate that disruption of one of the two LAL regulatory genes completely abolished geldanamycin biosynthesis. This means that both, gdmRI and gdmRII genes, are independent and are not simply duplicated regulatory genes. He et al. [35] showed that expression of these two genes is independent of each other; i.e., inactivation of one of these genes did not affect expression of the other one. These authors proved that inactivation of the gdmRI or gdmRII gene suppresses expression of the polyketide biosynthetic genes but does not affect transcription of post-polyketide modification genes and we confirmed that deletion of gdmRII resulted in lack of production of geldanamycin (Figure 3B). Using S. hygroscopicus NRRL3602, we amplified in an integrative monocopy vector either gdmRI, gdmRII, or both. The results indicated that production of geldanamycin increased about 33% with an additional copy of either gdmRI or gdmRII, but an important increase (more than 100%) was found when the wild-type strain was transformed with a combination of gdmRI and gdmRII (Figure 3A).
Later Kim et al. [36] studied the regulatory genes located close to the geldanamycin gene cluster from a different strain, S. hygroscopicus var. duamyceticus JCM4427, obtained from the Japanese culture collection of microorganisms (Table 1). These authors found up to five putative regulatory genes near the geldanamycin cluster; three of which were located downstream of the gdm cluster; these three genes, named gel14, gel17, and gel19, were characterized in detail [36]. Genes gel14 and gel17 encode LAL-type regulators that are identical to GdmRI and GdmRII described in S. hygroscopicus 17997 [35]. The identity between the two LAL-type regulator Gel14 and Gel17 is low (26%) and this explains why these two regulators are not functionally equivalent and work independently of each other in controlling geldanamycin biosynthesis [35]. On the other hand, gel19 encodes a TetR-type transcriptional factor that acts as a positive regulator of geldanamycin biosynthesis. The gel19 regulator belongs to the well-known family of TetR transcriptional factors, which are characterized as homodimers with an DNA binding helix-turn-helix domain at their N-terminal region and a ligand binding domain in the C-terminal end [39,40]. Studies of these regulatory genes and comparison of gene expression in mutants disrupted in each of these genes provide evidence indicating that regulation in strain S. hygroscopicus JCM4427 is somehow different from that found previously in S. hygroscopicus 17997. Indeed, in strain S. hygroscopicus JCM4427, gel17 and gel19 are required for expression of the gel14 LAL-type regulator and of the common promoter of the gelA genes (equivalent to gdmA, encoding the three polyketide synthases). Mutants in gel14 were able to express normally gel17 and gel19, whereas mutants in gel17 and gel19 did not express the LAL regulator gel14 or the polyketide synthase gene. In conclusion, the available evidence suggests that there is a cascade in which expression of gel14 is controlled by gel17 and gel19 and, in turn, gel14 is required for expression of GelA and controls initiation of transcription of gel8 (carbamoyl transferase) and gel16 (P450 oxygenase).
LAL-type regulators similar to Gel14 and Gel17 are encoded in many other polyketide gene clusters as, for instance PimM of the pimaricin producer Streptomyces natalensis [41,42], FkbN of the tacrolimus producers S. tsukubaensis and S. hygroscopicus var. ascomyceticus [43,44] or FscRI in the candicidin producer Streptomyces griseus [32,45], among others.
5. Engineering of Polyketide Biosynthesis in Geldanamycin Producing Strains
In the last decades the availability of genome sequences of different Streptomyces species has provided evidence indicating that many Streptomyces contain type I polyketides gene clusters [50,51]. The biosynthesis of these polyketides requires acyl-CoA precursor units including malonyl-CoA methylmalonyl-CoA, methoxymalonyl-CoA, allylmalonyl-CoA, among others. When these polyketides are produced simultaneously, they compete for the available common precursors. In the S. hygroscopicus 17997 strain, Li et al. [52] disrupted two polyketide genes different from that of geldanamycin and observed distinct effects; disruption of the polyketide cos-10 gene did not affect geldanamycin production but in contrast, disruption of the polyketide pg-10 gene doubled the production of geldanamycin supporting the conclusion that there is competition between the biosynthesis of geldanamycin and that of Pg-10 polyketide, which does not occur with other polyketides. At this time, the molecular mechanism of the specific competition between geldanamycin and polyketide Pg-10 is not yet clear.
Several important efforts have been made to obtain novel geldanamycin derivatives by modification of the biosynthetic pathway [53]. Moreover, Kim et al. [13] obtained, by combined mutagenesis of the geldanamycin PKS and modification of the tailoring enzymes, a C15-hydroxylated, C17-demethoxy non-quinone geldanamycin analogue, DHQ3, that has 4.6-fold higher Hsp90 ATPase activity inhibition than geldanamycin
6. Phosphate Control of Geldanamycin Biosynthesis
Growth of the Streptomyces species and the biosynthesis of antibiotics and other secondary metabolites is controlled by the concentration of phosphate in the medium [54]. However, there are important differences in the sensitivity to phosphate of the biosynthesis of distinct secondary metabolites [55,56,57].
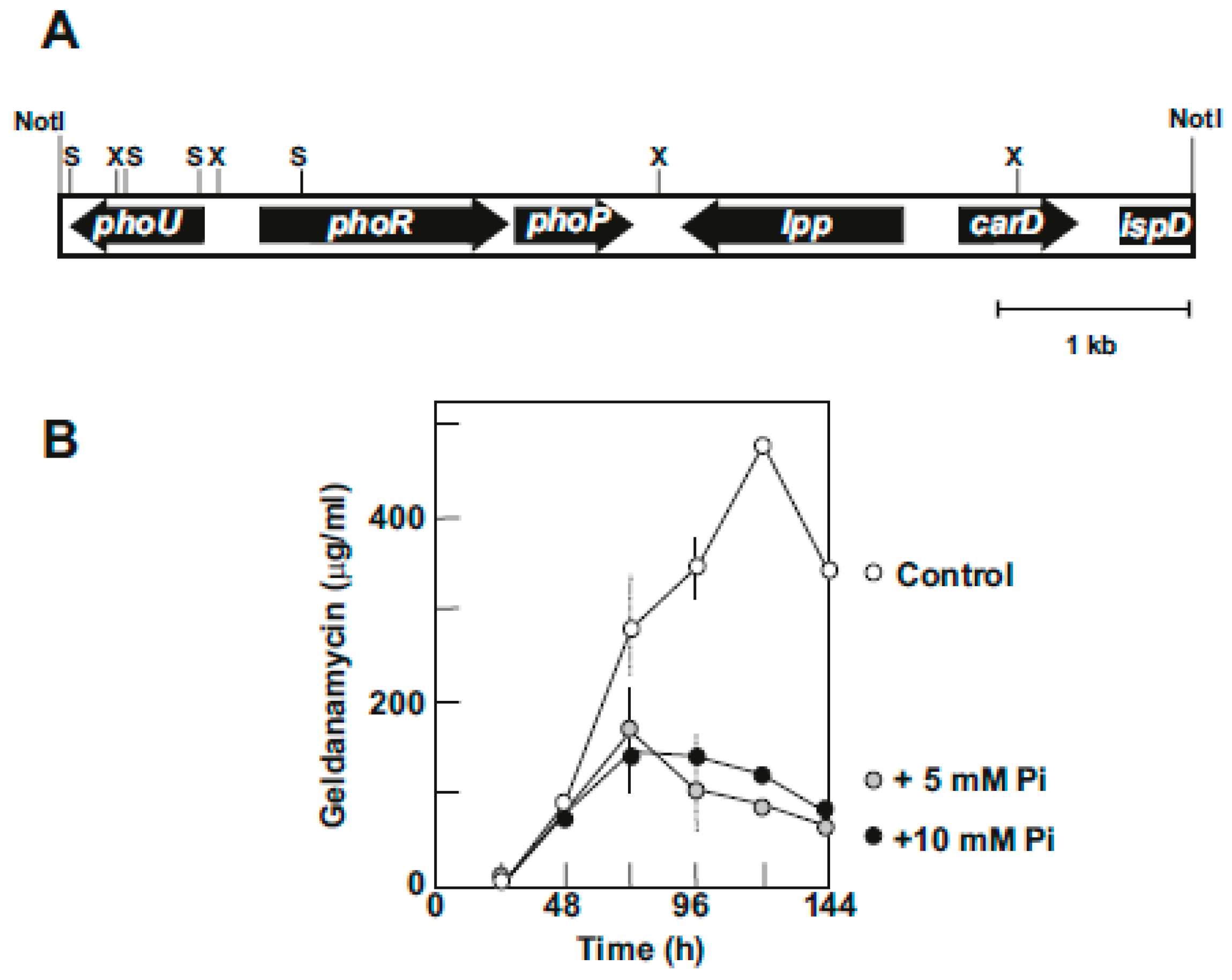
Our studies on the effect of inorganic phosphate in production of geldanamycin by S. hygroscopicus var. geldanus NRRL3602 showed that the biosynthesis of geldanamycin is highly sensitive to inorganic phosphate (Figure 4B). In soy-peptone-glucose (SPG) medium, which supports high geldanamycin production (above 500 μg/mL at 120 h), inorganic phosphate at 5 mM or higher concentration reduces geldanamycin production by 80%.
6.1. Cloning of the phoU- phoRP Gene Cluster of S. hygroscopicus NRRL3602
Phosphate control of both primary metabolism and the biosynthesis of secondary metabolites is mediated by the two components PhoR-PhoP system [58,59].
A cosmid library of total DNA of S. hygroscopicus var geldanus in the superCos1 vector (Stratagene) was constructed and screened by hybridization with a 1.2 kb DNA probe containing the phoRP genes of S. coelicolor [60]. A cosmid, Cos17d1, showing a high hybridization signal was selected and a 7.0 kb fragment of the insert was subcloned in pBluescript and sequenced. Five orfs were found in this insert that include phoR, phoP, phoU, lpp, carD, and the incomplete ispD gene (Figure 4A).
phoR-phoP were located in the opposite orientation to phoU; the three genes are expressed from a divergent promoter region as occurs in other Streptomyces species [56,57]. The lpp gene encode a lipoprotein, the carD gene encodes a CarD family transcriptional regulator that binds RNA polymerase, and the incomplete ispD encodes a cytidyltransferase.
The S. hygroscopicus sensor kinase PhoR is a protein of 432 amino acids that contains the characteristic Boxes H, N, D/F, and G [61] present in all PhoR proteins. PhoR has a high identity in amino acid sequence to the orthologous proteins of S. coelicolor (85%), S. avermitilis (86%), and S. natalensis (88%).
The response regulator PhoP has 223 amino acids and is 98% identical to the homologous protein of S. coelicolor and S. avermitilis. PhoP is extremely well conserved in all Streptomyces species [62] supporting its important role on the PhoR-PhoP mediated response [63]. The PhoP protein has the conserved aminoacids Asp6 and Asp49 in the amino terminal region, lysine K98 for phosphorylation, as part of the receiver domain, and a DNA binding domain (DBD) in the carboxyl terminal end (amino acids 190-201). The distance between phoR and phoP is only 5 nucleotides, strongly suggesting that these two genes are co-transcribed as reported previously in S. coelicolor [60,64].
The divergent phoU gene encodes a 275 amino acids protein 92% identical to the PhoU of S. natalensis and 93% to those of S. coelicolor and S. avermitilis. In S. coelicolor, phoU encodes a modulator of the PhoRP expression that exerts self-control of phosphate regulation [64]. In addition, the pstS gene, encoding the high affinity phosphate transport was cloned by hybridization with a probe of the orthologous S. coelicolor gene.
6.2. Disruption and Characterization of the PhoP Gene: Effect on Growth and Geldanamycin Production
In order to study the effect of PhoP on growth and production of geldanamycin, the PhoP gene was disrupted by an apramycin resistance gene in cosmid Cos17-D1 in E. coli and the disrupted phoP gene was then replaced in S. hygroscopicus by the REDIRECT technique. The mutation was confirmed by PCR amplification and sequencing of a 2.6 kb XhoI fragment containing the gene replacement. The nucleotide sequence of this fragment confirmed the disruption of phoP.
To study the effect of the inactivation of phoP on phosphate utilization, growth and geldanamycin production, the phoP mutant was grown in SPG, a medium that is an excellent nutrient for polyketide antibiotics production.
Interestingly, the disruption of phoP had a very strong effect on growth of S. hygroscopicus. The mutant was unable to grow in SPG medium in the absence of phosphate supplementation, although the SPG medium contains organic phosphate i.e,. it behaved as an inorganic phosphate auxotroph. When SPG medium was supplemented with 2.5, 5, 9, or 15 mM phosphate, there was an increasing recovery of growth at the 5-, 9- and 15-mM concentration, but the mutant showed very limited growth in 2.5 mM phosphate supplemented medium. This strong dependence of growth has not been observed in S. coelicolor or S. natalensis phoP mutants using the same medium. These two Streptomyces species grow in the absence of inorganic phosphate because they were able to hydrolyze organic phosphate present in the soy-peptone medium, whereas S. hygroscopicus seems unable to do so. This might be due to the absence of some extracellular phosphatases in S. hygroscopicus, as described also in S. tsukubaensis [62]. Although S. hygroscopicus phoP mutant grows well at 5 mM phosphate, the production of geldanamycin was higher at 15 mM which is a phosphate concentration normally inhibitory for antibiotic biosynthesis, as is the case in S. natalensis for pimaricin biosynthesis [55], suggesting that the phoP mutant is deregulated in PhoP mediated control of geldanamycin biosynthesis.
6.3. Identification of PhoP Binding Sequences in S. hygroscopicus Genes
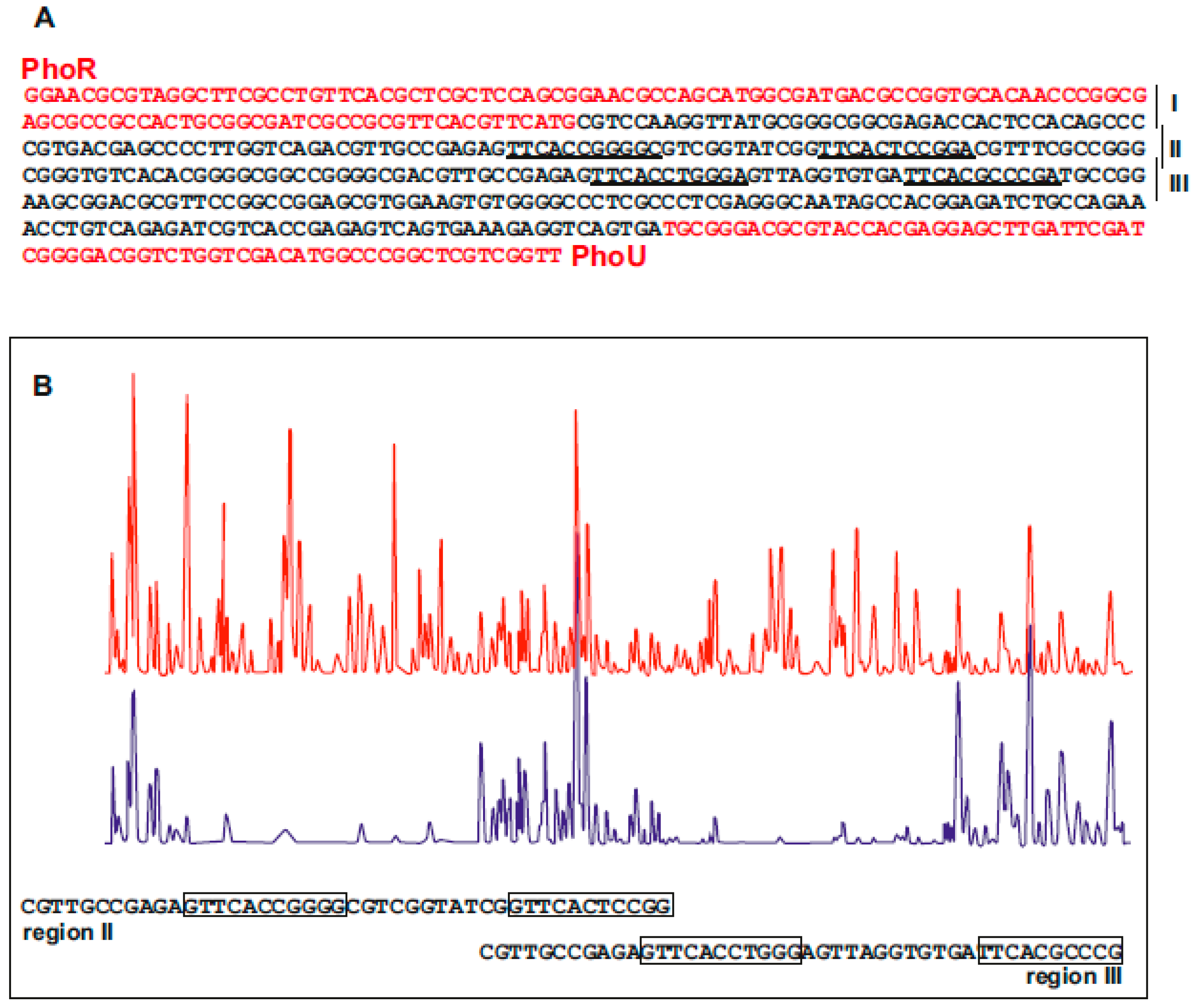
It is known that PhoP binds an 11 nucleotides direct repeat sequence in the genome of different Streptomyces species [56,60,65]. Although the genome of S. hygroscopicus var geldanus was not available, we searched for PhoP binding boxes in the promoter regions of phoU, phoRP, and pstS genes that were cloned as indicated above. Bioinformatic analysis of the promoter regions of these genes allowed us to identify the putative PHO boxes in these promoters. The intergenic sequence phoRP-phoU (Figure 5A) was divided in three segments (regions I, II, III) that were subjected to EMSA analysis using S. coelicolor PhoPDBD linked to-GST (0.2 to 3.2 pMols) obtained as a recombinant protein in E. coli [60]. The results showed a clear retardation in the mobility assays of the DNA fragments in regions II and III but no gel shift was observed in region I. Bioinformatic analysis of these regions and of the pstS gene promoter showed the presence of putative PHO boxes for PhoP binding. To confirm the nature of these PhoP binding sequences, a DNAse-footprinting analysis was performed on a 389 bp DNA fragment carrying the regions II and III upstream of the phoRP-phoU promoter and PhoPDBD-GST protein at 2 μM concentration. A DNA protection footprinting analysis showed two protected regions (Figure 5B) containing the 11-nucleotides repeats (PHO boxes) detected also by bioinformatic analysis. Alignment of the PHO boxes of phoU-phoRP and pstS genes provided a DNA-binding sequence (GTTCACCCGCC), similar to that of S. coelicolor and S. avermitilis [56], although with minor differences in the frequency of alternative nucleotides.
In summary, our studies on the phosphate control of geldanamycin biosynthesis indicate regulation by the phosphate concentration in the medium as occurs with many other polyketides, but it is clearly more sensitive to the inorganic phosphate concentration than other polyketides.
7. Future Outlook
The ansamycin geldanamycin and its derivatives have great potential as antitumor agents due to their interaction with the Hsp-90 chaperone in human cells. Clinical assays have shown that geldanamycin produces hepatoxicity and side effects affecting healthy human celIs. Novel geldanamycin derivatives obtained by metabolic engineering have been developed. Research on novel strains producing less toxic geldanamycin analogs is required. Moreover, at this time geldanamycin is an important tool for research in human tumorigenesis, tumor cell dissemination and cell apoptosis [67]. Biosynthesis of geldanamycin is well known but the regulatory mechanisms that control expression of geldanamycin biosynthesis genes needs further research. There are several regulatory mechanisms that control gene expression limiting the production of this antitumor agent; expression of the three positive regulators gdmRI, gdmRII, and gdmRIII need to be optimized to obtain maximal geldanamycin production; the nucleotide sequences recognized by each of these transcriptional regulators have to be elucidated in order to search for additional target genes. Geldanamycin biosynthesis is extremely sensitive to the phosphate concentration in the culture medium. Phosphate is limiting for growth but, at high concentration, strongly represses the biosynthetic genes and therefore it has an important regulatory role both in growth and geldanamycin production. Moreover, other pleiotropic regulators that control nitrogen metabolism, carbon source utilization, and coordination of metabolism are not known in these Streptomyces species and need to be investigated. The balance between utilization of nitrogen and carbon sources is very important in the biosynthesis of geldanamycin because of the involvement of the mC7N precursor unit that requires an amino group to form the AHBA starter unit. The availability of the genome sequence of both S. hygroscopicus XM-201 [21] and S. autolyticus CGMCC 0516 [23] provides new useful “omics” information to dissect the genome of these geldanamycin producing strains. Transcriptomic analysis of expression of different genes involved both in the biosynthesis of structural components of geldanamycin and in the control of the biosynthesis of this antitumor agent needs to be emphasized. Finally, metabolomic studies leading to removal of side-product contaminants are also required to further increase the production of geldanamycin and related.
8. Conclusions
The biosynthesis of geldanamycin in different producer strains is regulated by several transcriptional factors. Three of them gdmRI, gdmRII and gdmRIII are situated close to the geldanamycin gene cluster. Moreover, the biosynthesis of geldanamycin is strongly regulated by phosphate; this regulation is mediated by the two-component system PhoR-PhoP. The PhoP binding sequence in S. hygroscopicus have been identified in this article.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. Investigation, A.R.; writing—review and editing, J.F.M. and P.L.; writing—original draft preparation, P.L.; supervision, J.F.M.; funding acquisition, J.F.M.
Funding
This research was funded in 2007 by the Spanish Ministry of Education and Science, grant number FIT-010000-2007-76.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sasaki, K.; Rinehart, K.L.; Slomp, G.; Grostic, M.F.; Olson, E.C. Geldanamycin. I. Structure assignment. J. Am. Chem. Soc. 1970, 92, 7591–7593. [Google Scholar] [PubMed]
- Fukuyo, Y.; Hunt, C.R.; Horikoshi, N. Geldanamycin and its anti-cancer activities. Cancer Lett. 2010, 290, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Keum, G.; Pae, A.N. Discovery and development of heat shock protein 90 inhibitors as anticancer agents: a review of patented potent geldanamycin derivatives. Expert Opin. Ther. Pat. 2013, 23, 919–943. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, C.; Meulman, P.A.; Wnuk, R.J.; Peterson, D.H. Geldanamycin, a new antibiotic. J. Antibiot. 1970, 23, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Iwai, Y.; Nakagawa, A.; Sadakane, N.; Omura, S.; Oiwa, H.; Matsumoto, S.; Takahashi, M.; Ikai, T.; Ochiai, Y. Herbimycin B, a new benzoquinonoid ansamycin with anti-TMV and herbicidal activities. J. Antibiot. 1980, 33, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Iwai, Y.; Takahashi, Y.; Sadakane, N.; Nakagawa, A.; Oiwa, H.; Hasegawa, Y.; Ikai, T. Herbimycin, a new antibiotic produced by a strain of Streptomyces. J. Antibiot. 1979, 32, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Whitesell, L.; Mimnaugh, E.G.; De Costa, B.; Myers, C.E.; Neckers, L.M. Inhibition of heat shock protein HSP90-pp60v-src heteroprotein complex formation by benzoquinone ansamycins: essential role for stress proteins in oncogenic transformation. Proc. Natl. Acad. Sci. USA 1994, 91, 8324–8328. [Google Scholar] [CrossRef] [PubMed]
- Franke, J.; Eichner, S.; Zeilinger, C.; Kirschning, A. Targeting heat-shock-protein 90 (Hsp90) by natural products: geldanamycin, a show case in cancer therapy. Nat. Prod. Rep. 2013, 30, 1299–1323. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Yasuda, H.; Onodera, K. Growth inhibition of virus transformed cells in vitro and antitumor activity in vivo of geldanamycin and its derivatives. J. Antibiot. 1979, 32, 849–851. [Google Scholar] [CrossRef]
- Uehara, Y.; Murakami, Y.; Suzukake-Tsuchiya, K.; Moriya, Y.; Sano, H.; Shibata, K.; Omura, S. Effects of herbimycin, derivatives on src oncogene function in relation to antitumor activity. J. Antibiot. 1988, 41, 831–834. [Google Scholar] [CrossRef]
- Supko, J.G.; Hickman, R.L.; Grever, M.R.; Malspeis, L. Preclinical pharmacologic evaluation of geldanamycin as an antitumor agent. Cancer Chemother. Pharmacol. 1995, 36, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Ryu, J.S.; Jin, Y.; Kim, W.; Kaur, N.; Chung, S.J.; Jeon, Y.J.; Park, J.T.; Bang, J.S.; Lee, H.S.; et al. Synthesis and anticancer activity of geldanamycin derivatives derived from biosynthetically generated metabolites. Org. Biomol. Chem. 2008, 6, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Lee, D.; Hong, S.S.; Na, Z.; Shin, J.C.; Roh, S.H.; Wu, C.Z.; Choi, O.; Lee, K.; Shen, Y.M.; et al. Rational biosynthetic engineering for optimization of geldanamycin analogues. ChemBioChem 2009, 10, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Kitson, R.R.; Chang, C.H.; Xiong, R.; Williams, H.E.; Davis, A.L.; Lewis, W.; Dehn, D.L.; Siegel, D.; Roe, S.M.; Prodromou, C.; et al. Synthesis of 19-substituted geldanamycins with altered conformations and their binding to heat shock protein Hsp90. Nat. Chem. 2013, 5, 307–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.; Lu, T.; Zhao, L.X.; Chen, Y.; Huang, S.X.; Lohman, J.R.; Xu, L.H.; Jiang, C.L.; Shen, B. The missing C-17 O-methyltransferase in geldanamycin biosynthesis. Org. Lett. 2011, 13, 3726–3729. [Google Scholar] [CrossRef] [PubMed]
- Ivy, P.S.; Schoenfeldt, M. Clinical trials referral resource: current clinical trials of 17-AG and 17-DMAG. Oncology 2004, 18, 610, 615, 619–620. [Google Scholar]
- Schnur, R.C.; Corman, M.L.; Gallaschun, R.J.; Cooper, B.A.; Dee, M.F.; Doty, J.L.; Muzzi, M.L.; Moyer, J.D.; DiOrio, C.I.; Barbacci, E.G.; et al. Inhibition of the oncogene product p185erbB-2 in vitro and in vivo by geldanamycin and dihydrogel danamycin derivatives. J. Med. Chem. 1995, 38, 3806–3812. [Google Scholar] [CrossRef]
- Rascher, A.; Hu, Z.; Viswanathan, N.; Schirmer, A.; Reid, R.; Nierman, W.C.; Lewis, M.; Hutchinson, C.R. Cloning and characterization of a gene cluster for geldanamycin production in Streptomyces hygroscopicus NRRL 3602. FEMS Microbiol. Lett. 2003, 218, 223–230. [Google Scholar] [CrossRef]
- He, W.; Wu, L.; Gao, Q.; Du, Y.; Wang, Y. Identification of AHBA biosynthetic genes related to geldanamycin biosynthesis in Streptomyces hygroscopicus 17997. Curr. Microbiol. 2006, 52, 197–203. [Google Scholar] [CrossRef]
- Shin, J.C.; Na, Z.; Lee, D.H.; Kim, W.C.; Lee, K.; Shen, Y.M.; Paik, S.G.; Hong, Y.S.; Lee, J.J. Characterization of tailoring genes involved in the modification of geldanamycin polyketide in Streptomyces hygroscopicus JCM4427. J. Microbiol. Biotechnol. 2008, 18, 1101–1108. [Google Scholar]
- Wang, X.; Ning, X.; Zhao, Q.; Kang, Q.; Bai, L. Improved PKS Gene Expression With Strong Endogenous Promoter Resulted in Geldanamycin Yield Increase. Biotechnol. J. 2017, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Ouyang, Y.; Wang, G.; Li, X. Streptomyces autolyticus JX-47 large-insert bacterial artificial chromosome library construction and identification of clones covering geldanamycin biosynthesis gene cluster. Curr. Microbiol. 2011, 63, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Jiang, M.; Ren, Z.; Dong, Y.; Lu, T. The complete genome sequence of Streptomyces autolyticus CGMCC 0516, the producer of geldanamycin, autolytimycin, reblastatin and elaiophylin. J. Biotechnol. 2017, 252, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Boudjeko, T.; Tchinda, R.A.M.; Mina Zitouni, M.; Nana, J.A.V.T.; Lerat, S.; Beaulieu, C. Streptomyces cameroonensis sp. nov., a Geldanamycin Producer That Promotes Theobroma cacao Growth. Microbes Environ. 2017, 32, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhao, L.; Liu, H.W.; Sherman, D.H. A gene cluster for macrolide antibiotic biosynthesis in Streptomyces venezuelae: architecture of metabolic diversity. Proc. Natl. Acad. Sci. USA 1998, 95, 12111–12116. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; von Bamberg, D.; Hale, V.; Breuer, M.; Hardt, B.; Müller, R.; Floss, H.G.; Reynolds, K.A.; Leistner, E. Biosynthesis of ansatrienin (mycotrienin) and naphthomycin. Identification and analysis of two separate biosynthetic gene clusters in Streptomyces collinus Tü 1892. Eur. J. Biochem. 1999, 261, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.W.; Bai, L.; Clade, D.; Hoffmann, D.; Toelzer, S.; Trinh, K.Q.; Xu, J.; Moss, S.J.; Leistner, E.; Floss, H.G. The biosynthetic gene cluster of the maytansinoid antitumor agent ansamitocin from Actinosynnema pretiosum. Proc. Natl. Acad. Sci. USA 2002, 99, 7968–7973. [Google Scholar] [CrossRef]
- August, P.R.; Tang, L.; Yoon, Y.J.; Ning, S.; Müller, R.; Yu, T.W.; Taylor, M.; Hoffmann, D.; Kim, C.G.; Zhang, X.; et al. Biosynthesis of the ansamycin antibiotic rifamycin: deductions from the molecular analysis of the rif biosynthetic gene cluster of Amycolatopsis mediterranei S699. Chem. Biol. 1998, 5, 69–79. [Google Scholar] [CrossRef]
- Admiraal, S.J.; Walsh, C.T.; Khosla, C. The loading module of rifamycin synthetase is an adenylation-thiolation didomain with substrate tolerance for substituted benzoates. Biochemistry 2001, 40, 6116–6123. [Google Scholar] [CrossRef]
- Marahiel, M.A.; Stachelhaus, T.; Mootz, H.D. Modular peptide synthetases involved in non-ribosomal peptide synthesis. Chem. Rev. 1997, 97, 2651–2673. [Google Scholar] [CrossRef]
- Martín, J.F. Alpha-aminoadipyl-cysteinyl-valine synthetases in beta-lactam producing organisms. From Abraham’s discoveries to novel concepts of non-ribosomal peptide synthesis. J. Antibiot. 2000, 53, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Aparicio, J.F. Biosynthesis Enzymology of the Polyenes Pimaricin and Candicidin. Methods Enzymol. 2009, 459, 215–242. [Google Scholar] [PubMed]
- Eichner, S.; Eichner, T.; Floss, H.G.; Fohrer, J.; Hofer, E.; Sasse, F.; Zeilinger, C.; Kirschning, A. Broad substrate specificity of the amide synthase in S. hygroscopicus--new 20-membered macrolactones derived from geldanamycin. J. Am. Chem. Soc. 2012, 134, 1673–1679. [Google Scholar] [CrossRef]
- Li, Y.; He, W.; Wang, Y.; Wang, Y.; Shao, R. A new post-PKS modification process in the carbamoyltransferase gene inactivation strain of Streptomyces hygroscopicus 17997. J. Antibiot. 2008, 61, 347–355. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Lei, J.; Liu, Y.; Wang, Y. The LuxR family members GdmRI and GdmRII are positive regulators of geldanamycin biosynthesis in Streptomyces hygroscopicus 17997. Arch. Microbiol. 2008, 189, 501–510. [Google Scholar] [CrossRef]
- Kim, W.; Lee, J.J.; Paik, S.G.; Hong, Y.S. Identification of three positive regulators in the geldanamycin PKS gene cluster of Streptomyces hygroscopicus JCM4427. J. Microbiol. Biotechnol. 2010, 20, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Rimal, H.; Yu, S.C.; Lee, B.; Hong, Y.S.; Oh, T.J. Characterization of Gel16 as a Cytochrome P450 in Geldanamycin Biosynthesis and in-silico Analysis for an Endogenous Electron Transport System. J. Microbiol. Biotechnol. 2019, 29, 44–54. [Google Scholar] [CrossRef]
- Pérez-Redondo, R.; Santamarta, I.; Bovenberg, R.; Martín, J.F.; Liras, P. The enigmatic lack of glucose utilization in Streptomyces clavuligerus is due to inefficient expression of the glucose permease gene. Microbiology 2010, 156, 1527–1537. [Google Scholar] [CrossRef]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef]
- Cuthbertson, L.; Nodwell, J.R. The TetR family of regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef]
- Santos-Aberturas, J.; Payero, T.D.; Vicente, C.M.; Guerra, S.M.; Cañibano, C.; Martín, J.F.; Aparicio, J.F. Functional conservation of PAS-LuxR transcriptional regulators in polyene macrolide biosynthesis. Metab. Eng. 2011, 13, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Santos-Aberturas, J.; Vicente, C.M.; Guerra, S.M.; Payero, T.D.; Martín, J.F.; Aparicio, J.F. Molecular control of polyene macrolide biosynthesis: direct binding of the regulator PimM to eight promoters of pimaricin genes and identification of binding boxes. J. Biol. Chem. 2011, 286, 9150–9161. [Google Scholar] [CrossRef] [PubMed]
- Goranovič, D.; Blažič, M.; Magdevska, V.; Horvat, J.; Kuščer, E.; Polak, T.; Santos-Aberturas, J.; Martínez-Castro, M.; Barreiro, C.; Mrak, P.; et al. FK506 biosynthesis is regulated by two positive regulatory elements in Streptomyces tsukubaensis. BMC Microbiol. 2012, 12, 238. [Google Scholar]
- Ordóñez-Robles, M.; Rodríguez-García, A.; Martín, J.F. Target genes of the Streptomyces tsukubaensis FkbN regulator include most of the tacrolimus biosynthesis genes, a phosphopantetheinyl transferase and other PKS genes. Appl. Microbiol. Biotechnol. 2016, 100, 8091–8103. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Huang, X.; Zhou, X.; Bai, L.; He, J.; Jeong, K.J.; Lee, S.Y.; Deng, Z. Organizational and mutational analysis of a complete FR-008/candicidin gene cluster encoding a structurally related polyene complex. Chem. Biol. 2003, 10, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef]
- Jiang, M.; Yin, M.; Wu, S.; Han, X.; Ji, K.; Wen, M.; Lu, T. GdmRIII, a TetR Family Transcriptional Regulator, Controls Geldanamycin and Elaiophylin Biosynthesis in Streptomyces autolyticus CGMCC0516. Sci. Rep. 2017, 7, 4803. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Wang, L.; Zhang, G.; Fan, K.; Tan, H.; Yang, K. A novel role of ‘pseudo’γ-butyrolactone receptors in controlling γ-butyrolactone biosynthesis in Streptomyces. Mol. Microbiol. 2011, 82, 236–250. [Google Scholar] [CrossRef]
- Martínez-Burgo, Y.; Álvarez-Álvarez, R.; Rodríguez-García, A.; Liras, P. The Pathway-Specific Regulator ClaR of Streptomyces clavuligerus Has a Global Effect on the Expression of Genes for Secondary Metabolism and Differentiation. Appl. Environ. Microbiol. 2015, 81, 6637–6648. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef]
- Omura, S.; Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Takahashi, C.; Shinose, M.; Takahashi, Y.; Horikawa, H.; Nakazawa, H.; Osonoe, T.; et al. Genome sequence of an industrial microorganism Streptomyces avermitilis: deducing the ability of producing secondary metabolites. Proc. Natl. Acad. Sci. USA 2001, 98, 12215–12220. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Li, M.; He, W.Q.; Wang, Y.G.; Shao, R.G. Inactivation of putative PKS genes can double geldanamycin yield in Streptomyces hygroscopicus 17997. Genet. Mol. Res. 2013, 12, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Piagentini, M.; Rascher, A.; Tian, Z.Q.; Buchanan, G.O.; Regentin, R.; Hu, Z.; Hutchinson, C.R.; McDaniel, R. Engineered biosynthesis of geldanamycin analogs for Hsp90 inhibition. Chem. Biol. 2004, 11, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Demain, A.L. Control of antibiotic biosynthesis. Microbiol. Rev. 1980, 44, 230–251. [Google Scholar] [PubMed]
- Mendes, M.V.; Tunca, S.; Antón, N.; Recio, E.; Sola-Landa, A.; Aparicio, J.F.; Martín, J.F. The two-component PhoR-PhoP system of Streptomyces natalensis: Inactivation or deletion of phoP reduces the negative phosphate regulation of pimaricin biosynthesis. Metab. Eng. 2007, 9, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.F.; Rodríguez-García, A.; Liras, P. The master regulator PhoP coordinates phosphate and nitrogen metabolism, respiration, cell differentiation and antibiotic biosynthesis: comparison in Streptomyces coelicolor and Streptomyces avermitilis. J. Antibiot. 2017, 70, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Robles, M.; Santos-Beneit, F.; Rodríguez-García, A.; Martín, J.F. Analysis of the Pho regulon in Streptomyces tsukubaensis. Microbiol. Res. 2017, 205, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Moura, R.S.; Martín, J.F. The two-component PhoR-PhoP system controls both primary metabolism and secondary metabolite biosynthesis in Streptomyces lividans. Proc. Natl. Acad. Sci. USA 2003, 100, 6133–6138. [Google Scholar] [CrossRef]
- Martín, J.F. Phosphate control of the biosynthesis of antibiotics and other secondary metabolites is mediated by the PhoR-PhoP system: An unfinished story. J. Bacteriol. 2004, 186, 5197–5201. [Google Scholar]
- Sola-Landa, A.; Rodríguez-García, A.; Franco-Domínguez, E.; Martín, J.F. Binding of PhoP to promoters of phosphate-regulated genes in Streptomyces coelicolor: identification of PHO boxes. Mol. Microbiol. 2005, 56, 1373–1385. [Google Scholar] [CrossRef]
- Jung, K.; Fried, L.; Behr, S.; Heermann, R. Histidine kinases and response regulators in networks. Curr. Opin. Microbiol. 2012, 15, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castro, M.; Barreiro, C.; Martín, J.F. Analysis and validation of the pho regulon in the tacrolimus-producer strain Streptomyces tsukubaensis: differences with the model organism Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2018, 102, 7029–7045. [Google Scholar]
- Martín, J.F.; Santos-Beneit, F.; Rodríguez-García, A.; Sola-Landa, A.; Smith, M.C.; Ellingsen, T.E.; Nieselt, K.; Burroughs, N.J.; Wellington, E.M. Transcriptomic studies of phosphate control of primary and secondary metabolism in Streptomyces coelicolor. Appl. Microbiol. Biotechnol. 2012, 95, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Martín-Martín, S.; Rodríguez-García, A.; Santos-Beneit, F.; Franco-Domínguez, E.; Sola-Landa, A.; Martín, J.F. Self-control of the PHO regulon: the PhoP-dependent protein PhoU controls negatively expression of genes of PHO regulon in Streptomyces coelicolor. J. Antibiot. 2017. (In press) [Google Scholar]
- Allenby, N.E.; Laing, E.; Bucca, G.; Kierzek, A.M.; Smith, C.P. Diverse control of metabolism and other cellular processes in Streptomyces coelicolor by the PhoP transcription factor: genome-wide identification of in vivo targets. Nucleic Acids Res. 2012, 40, 9543–9556. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Rodríguez-García, A.; Amin, R.; Wohlleben, W.; Martín, J.F. Competition between the GlnR and PhoP regulators for the glnA and amtB promoters in Streptomyces coelicolor. Nucleic Acids Res. 2013, 41, 1767–1782. [Google Scholar] [CrossRef]
- Solárová, Z.; Mojžiš, J.; Solár, P. Hsp90 inhibitor as a sensitizer of cancer cells to different therapies. Int. J. Oncol. 2015, 46, 907–926. [Google Scholar]
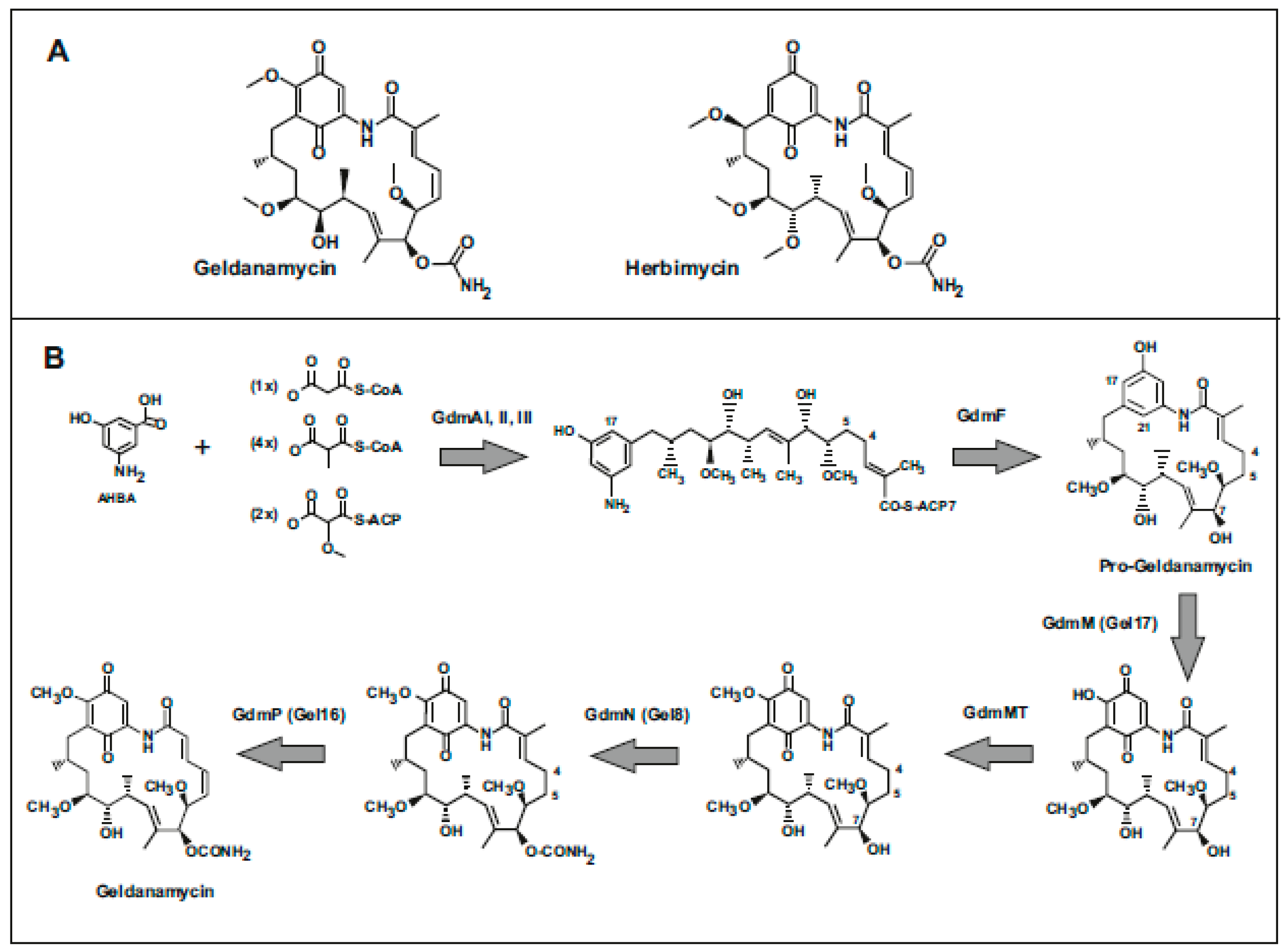
Figure 1.
Geldanamycin structure and biosynthesis pathway. (A) Structures of geldanamycin and herbimycin. (B) Geldanamycin biosynthetic pathway. The enzymes involved in every step are indicated.
Figure 1.
Geldanamycin structure and biosynthesis pathway. (A) Structures of geldanamycin and herbimycin. (B) Geldanamycin biosynthetic pathway. The enzymes involved in every step are indicated.

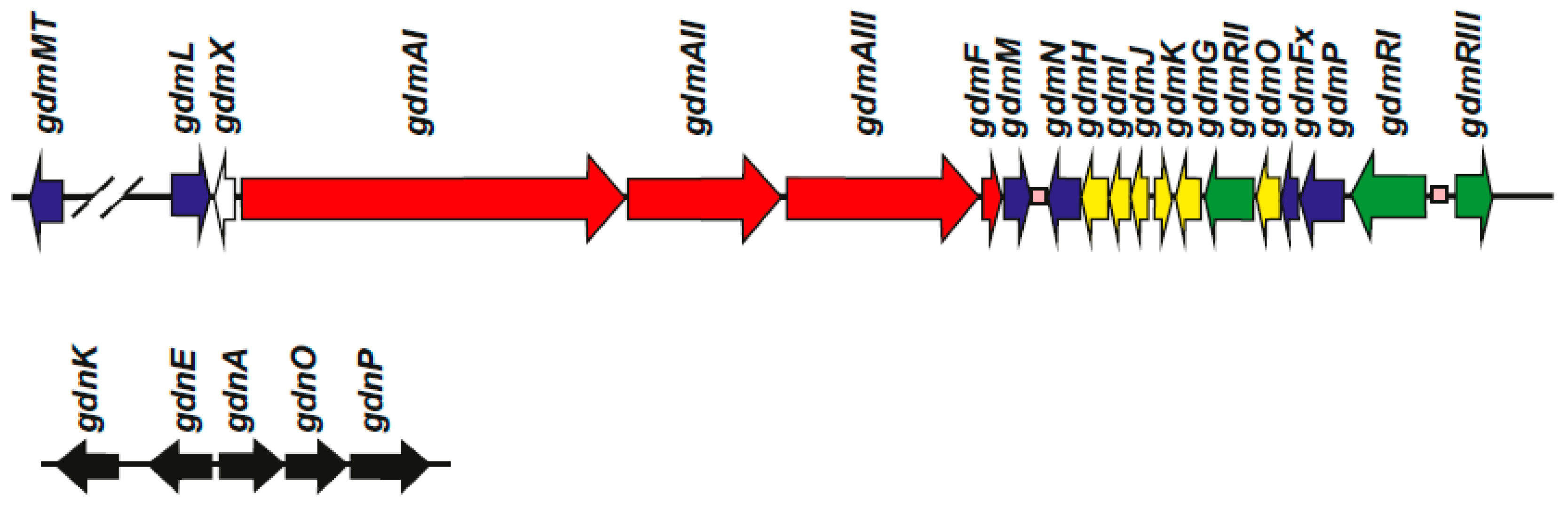
Figure 2.
Cluster of geldanamycin genes in S. hygroscopicus NRRL3602. Cluster of geldanamycin genes in S. hygroscopicus NRRL3602 [18]. The name of each gene is indicated above it. Similar gene clusters have been found in other S. hygroscopicus strains and in S. autolyticus with the minor differences described in the text. Genes for the geldanamycin precursor AHBA [19], named gdnKEAOP, and located separately in the genome, are shown below. The names of the geldanamycin genes given by different authors and function of the encoded enzymes are shown in Table 2.
Figure 2.
Cluster of geldanamycin genes in S. hygroscopicus NRRL3602. Cluster of geldanamycin genes in S. hygroscopicus NRRL3602 [18]. The name of each gene is indicated above it. Similar gene clusters have been found in other S. hygroscopicus strains and in S. autolyticus with the minor differences described in the text. Genes for the geldanamycin precursor AHBA [19], named gdnKEAOP, and located separately in the genome, are shown below. The names of the geldanamycin genes given by different authors and function of the encoded enzymes are shown in Table 2.

Figure 3.
Effect of the regulatory genes on geldanamycin production. (A) Production of geldanamycin by S. hygroscopicus var. geldanus NRRL3602 transformed with the empty plasmid pRA (black circles), and the pRA plasmid [38] carrying insertions with the genes gdmRI (white triangles), gdmRII (white squares), and gdmRI+gdmRII (white circles). (B) HPLC analysis of geldanamycin production by S. hygroscopicus var. geldanus NRRL3602 (upper panel) and S. hygroscopicus ΔgdmRII (lower panel) in which gdmRII was deleted. The arrow indicates the geldanamycin peak.
Figure 3.
Effect of the regulatory genes on geldanamycin production. (A) Production of geldanamycin by S. hygroscopicus var. geldanus NRRL3602 transformed with the empty plasmid pRA (black circles), and the pRA plasmid [38] carrying insertions with the genes gdmRI (white triangles), gdmRII (white squares), and gdmRI+gdmRII (white circles). (B) HPLC analysis of geldanamycin production by S. hygroscopicus var. geldanus NRRL3602 (upper panel) and S. hygroscopicus ΔgdmRII (lower panel) in which gdmRII was deleted. The arrow indicates the geldanamycin peak.

Figure 4.
Phosphate control of geldanamycin production. (A) Organization of the pho genes and the surrounding region in a NotI DNA fragment of S. hygroscopicus var. geldanus NRRL 3602. Restriction sites for XhoI (X) and SalI (S) are indicated. (B) Production of geldanamycin in SPG medium (white circles) and SPG medium supplemented with 5 mM (gray circles) and 10 mM inorganic phosphate (black circles).
Figure 4.
Phosphate control of geldanamycin production. (A) Organization of the pho genes and the surrounding region in a NotI DNA fragment of S. hygroscopicus var. geldanus NRRL 3602. Restriction sites for XhoI (X) and SalI (S) are indicated. (B) Production of geldanamycin in SPG medium (white circles) and SPG medium supplemented with 5 mM (gray circles) and 10 mM inorganic phosphate (black circles).

Figure 5.
Localization of PHO boxes in the intergenic region phoRP-PhoU of S. hygroscopicus var. geldanus NRRL 3602. (A) Sequence of the intergenic region phoRP-phoU showing in red the nucleotides belonging to the phoP and phoU genes. The regions I, II, and III used in EMSA analysis are indicated. (B) Footprinting analysis of DNA containing regions II and III. The upper line (in red) corresponds to the parental strain DNA without the PhoPDBD-GST protein. The lower line (in blue) corresponds to the same DNA sequence protected by addition of PhoPDBD-GST protein. PhoPDBD-GST protein (2 μM) was used in the assay as described by Sola-Landa et al. [66].
Figure 5.
Localization of PHO boxes in the intergenic region phoRP-PhoU of S. hygroscopicus var. geldanus NRRL 3602. (A) Sequence of the intergenic region phoRP-phoU showing in red the nucleotides belonging to the phoP and phoU genes. The regions I, II, and III used in EMSA analysis are indicated. (B) Footprinting analysis of DNA containing regions II and III. The upper line (in red) corresponds to the parental strain DNA without the PhoPDBD-GST protein. The lower line (in blue) corresponds to the same DNA sequence protected by addition of PhoPDBD-GST protein. PhoPDBD-GST protein (2 μM) was used in the assay as described by Sola-Landa et al. [66].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Strains producers of geldanamycin.
| Streptomyces Strains | Reference |
|---|---|
| Streptomyces hygroscopicus var geldanus NRRL3602 | [18] |
| Streptomyces hygroscopicus 17997 | [19] |
| Streptomyces hygroscopicus sub. duamyceticus JCM 4427 | [20] |
| Streptomyces hygroscopicus XM 201 | [21] |
| Streptomyces autolyticus JX-47 | [22] |
| Streptomyces autolyticus CGMCC 0516, | [23] |
| Streptomyces cameroonensis sp. | [24] |
| Streptomyces species containing geldanamycin biosynthesis genes identified bioinformatically | |
| Streptomyces violaceusniger Tu 4113 Streptomyces sp. RTd22 Streptomyces lydicus strain 103 Streptomyces rapamycinicus NRRL 5491 Streptomyces iranensis Streptomyces albus DSM 41398 Streptomyces bingchenggensis BCW-1 | Cited in [23] |
Table 2.
Genes in the geldanamycin cluster and function of the encoded enzymes.
| Genes | Function | Reference |
|---|---|---|
| gdmMT | O-methyltransferase | [15] |
| gdmL = gel1 | Flavin-dependent oxygenase | [20] |
| gdmX | Unknown | [18] |
| gdmAI, AII, AIII = gelAI, AII, AIII | Polyketide synthases I, II and III 1 | [18] 2 |
| gdmF | Amide synthase | [33] |
| gdmM = gel7 | Flavin-dependent oxidase | [20] |
| gdmN = gel8 | Carbamoyl transferase | [20,34] |
| gdmH | Methoxymalonyl-ACP biosynthesis | [18] |
| gdmI | Methoxymalonyl-ACP biosynthesis | [18] |
| gdmJ | Methoxymalonyl-ACP biosynthesis | [18] |
| gdmK | Methoxymalonyl-ACP biosynthesis | [18] |
| gdmG | O-methyl transferase for methoxy-malonyl-ACP biosynthesis | [18] |
| gdmRII = gel17 | LAL-type regulator | [35,36] |
| gdmO3 | Amino dehydroquinate synthase | [18] |
| gdmFx | Ferredoxin | [20] |
| gdmP = gel16 | P450 monooxygenase | [20] |
| gdmRI = gel14 | LAL-type regulator | [35,36] |
| gdmRIII = gel 19 | TetR-family positive regulator | [36] |
1. The GdmAI, AII, AIII proteins are translated from a putative monocistronic mRNA encoded by the gdmA gene. The original gdm gene designation was proposed by Rascher et al. [18]. 2. Other gene designations (gel genes) correspond to articles on the characterization of the genes. 3. Other genes for AHBA biosynthesis have been cloned in S. hygroscopicus 17997 by He et al. [19], but except the amino dehydroquinate synthase (gdmO) were not located in the geldanamycin cluster described by Rasher et al. [18].
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martín, J.F.; Ramos, A.; Liras, P. Regulation of Geldanamycin Biosynthesis by Cluster-Situated Transcription Factors and the Master Regulator PhoP. Antibiotics 2019, 8, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030087
AMA Style
Martín JF, Ramos A, Liras P. Regulation of Geldanamycin Biosynthesis by Cluster-Situated Transcription Factors and the Master Regulator PhoP. Antibiotics. 2019; 8(3):87. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030087
Chicago/Turabian StyleMartín, Juan F., Angelina Ramos, and Paloma Liras. 2019. "Regulation of Geldanamycin Biosynthesis by Cluster-Situated Transcription Factors and the Master Regulator PhoP" Antibiotics 8, no. 3: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8030087
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.