Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea?

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
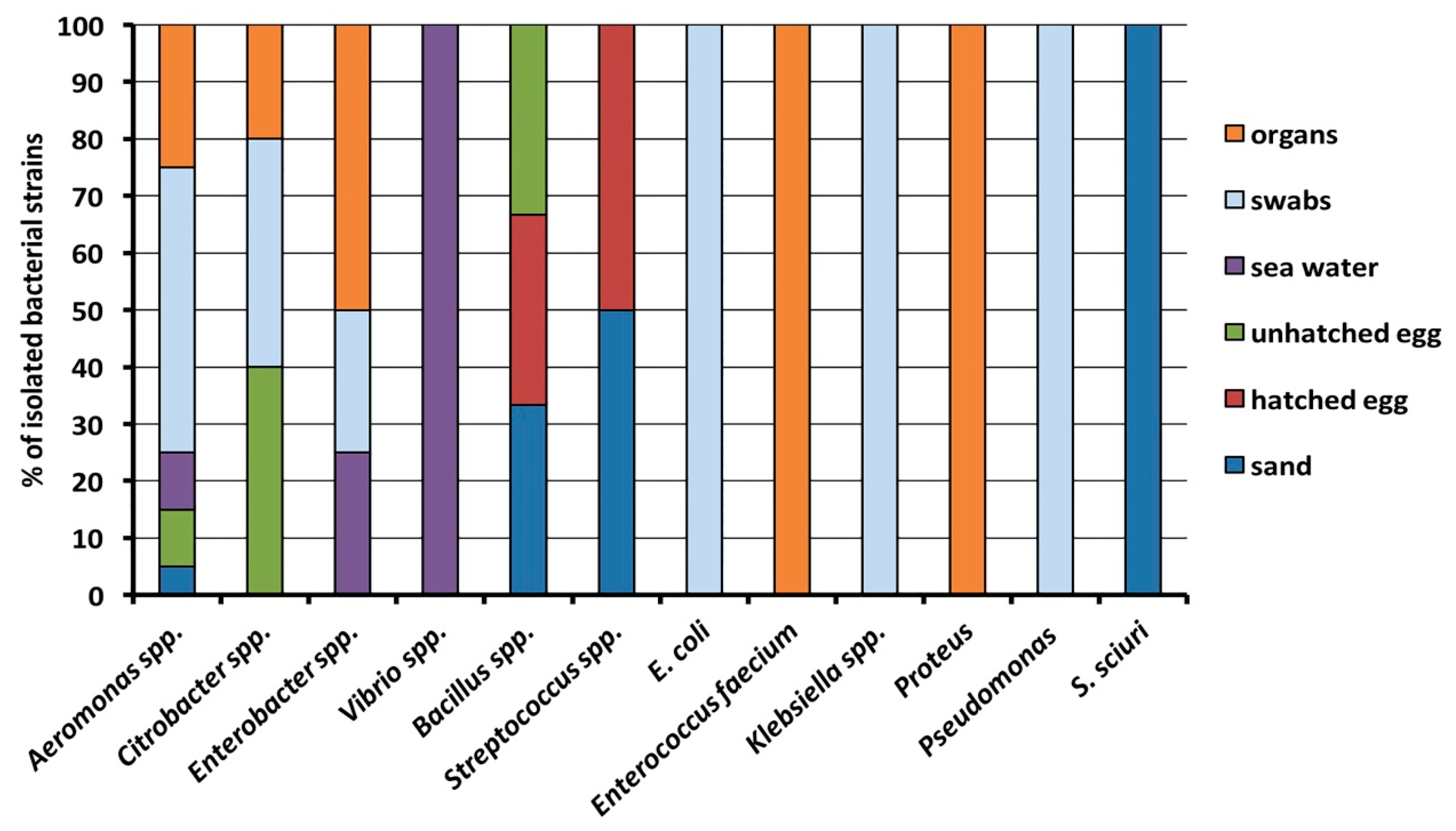
2.1. Bacterial Identification
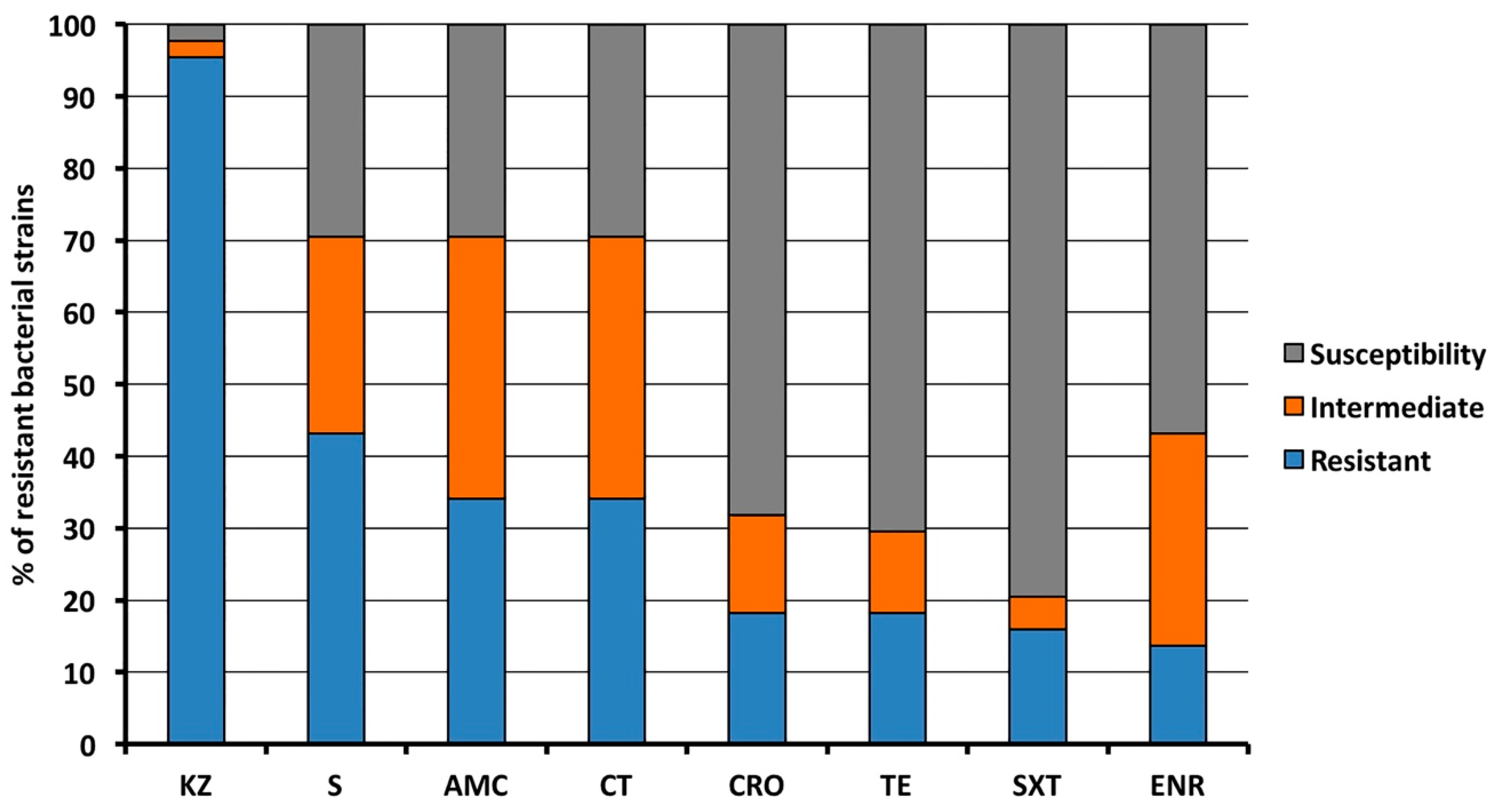
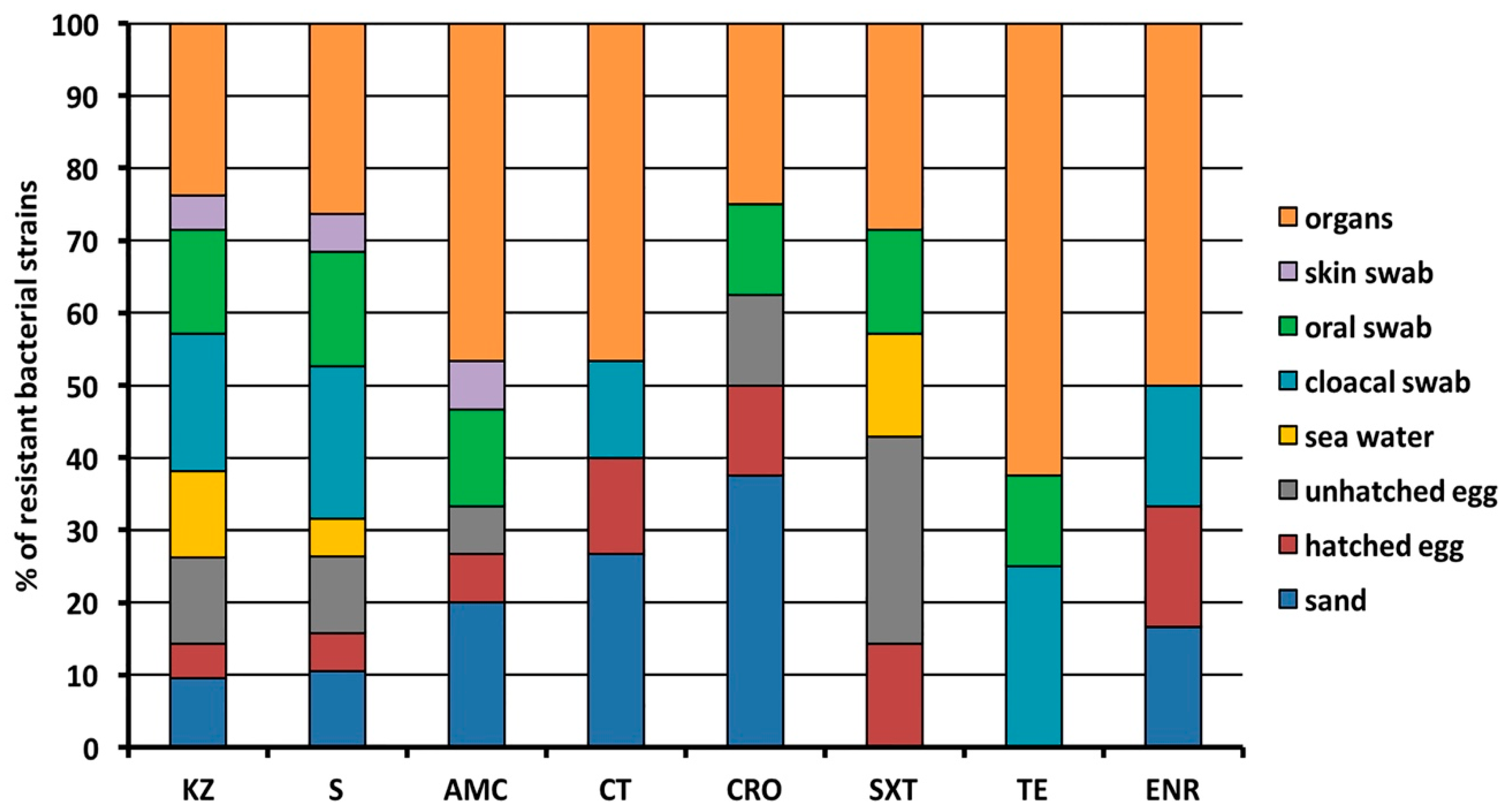
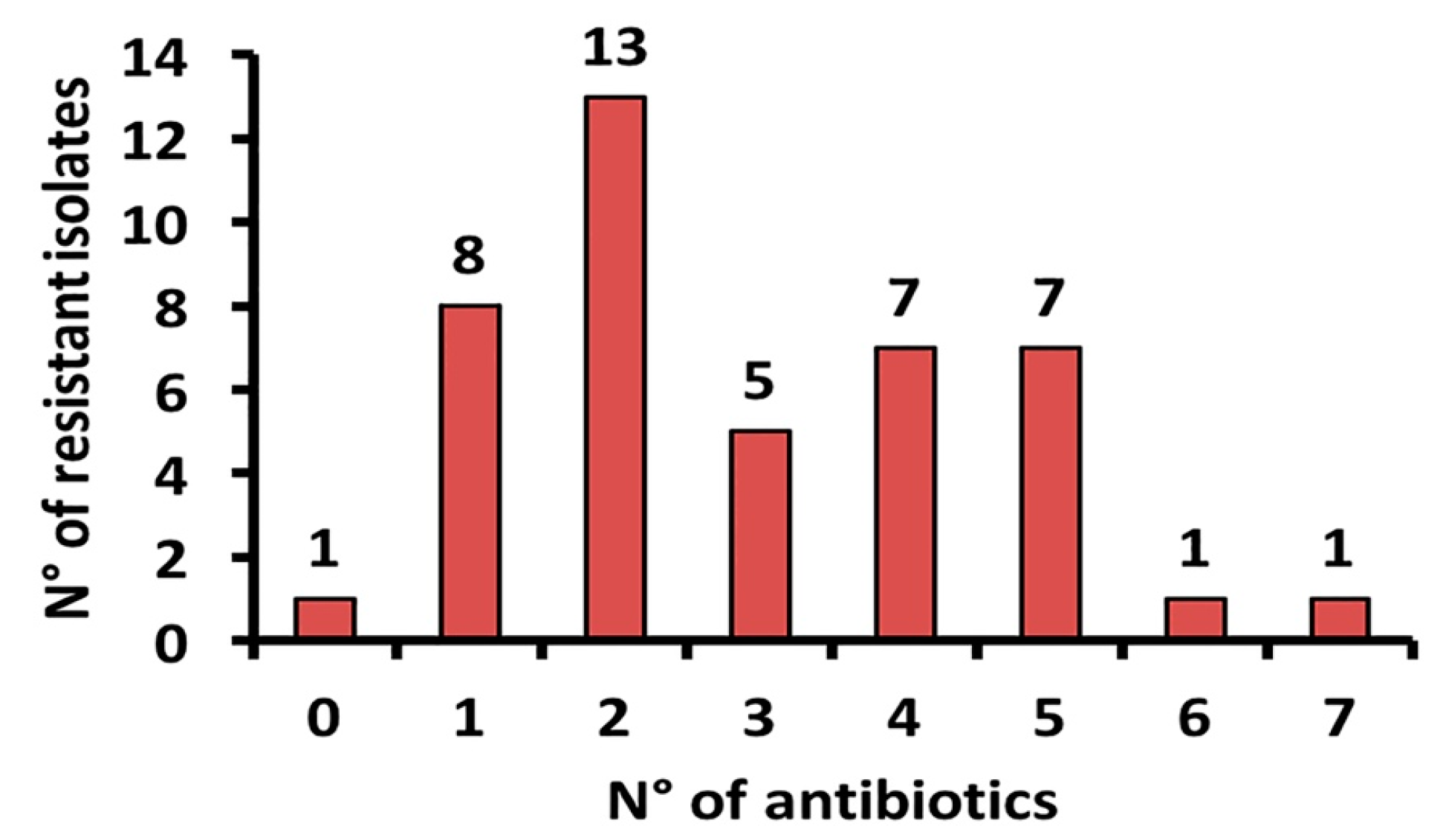
2.2. Antibacterial Resistance
2.3. Detection of Antibiotic and Heavy Metal Resistance Genes
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Bacterial Isolation
4.3. Antibiotic Susceptibility Test
4.4. Detection of Antibiotic and Heavy Metal Resistance Genes
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kraemer, S.A.; Ramachandran, A.; Perron, G.G. Antibiotic pollution in the environment: From microbial ecology to public policy. Microorganisms 2019, 7, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Carroll, D.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial resistance in wildlife: Implications for public health. Zoonoses Public Health 2015, 62, 534–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bahry, S.N.; Mahmoud, I.Y.; Al-Belushi, K.I.A.; Elshafie, A.E.; Al-Harthy, A.; Bakheit, C.K. Coastal sewage discharge and its impact on fish with reference to antibiotic resistant enteric bacteria and enteric pathogens as bio-indicators of pollution. Chemosphere 2009, 77, 1534–1539. [Google Scholar] [CrossRef]
- Al-Bahry, S.N.; Mahmoud, I.Y.; Al-Khaifi, A.; Elshafie, A.E.; Al-Harthy, A. Viability of multiple antibiotic resistant bacteria in distribution lines of treated sewage effluent used for irrigation. Water Sci. Technol. 2009, 60, 2939–2948. [Google Scholar] [CrossRef]
- Pontes, D.S.; Pinheiro, F.A.; Lima-Bittencourt, C.I.; Guedes, R.L.M.; Cursino, L.; Barbosa, F.; Santos, F.R.; Chartone-Souza, E.; Nascimento, A.M.A. Multiple antimicrobial resistance of gram-negative bacteria from natural oligotrophic lakes under distinct anthropogenic influence in a tropical region. Microb. Ecol. 2009, 58, 762–772. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy metal driven co-selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Fontaneto, D.; Doppelbauer, J.; Corno, G. Fitness and recovery of bacterial communities and antibiotic resistance genes in urban wastewaters exposed to classical disinfection treatments. Environ. Sci. Technol. 2016, 50, 10153–10161. [Google Scholar] [CrossRef]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Duarte, D.J.; Oldenkamp, R.; Ragas, A.M.J. Modelling environmental antibiotic-resistance gene abundance: A meta-analysis. Sci. Total Environ. 2019, 659, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Wallace, C.C.; Yund, P.O.; Ford, T.E.; Matassa, K.A.; Bass, A.L. Increase in antimicrobial resistance in bacteria isolated from stranded marine mammals of the northwest atlantic. EcoHealth 2013, 10, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R.; Townsend, F.I.; Lane, S.M.; Dyar, E.; Hohn, A.A.; Rowles, T.K.; Staggs, L.A.; Wells, R.S.; Balmer, B.C.; Schwacke, L.H. Survey of antibiotic-resistant bacteria isolated from bottlenose dolphins tursiops truncatus in the southeastern USA. Dis. Aquat. Org. 2014, 108, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Foti, M.; Giacopello, C.; Bottari, T.; Fisichella, V.; Rinaldo, D.; Mammina, C. Antibiotic resistance of gram negatives isolates from loggerhead sea turtles (Caretta caretta) in the central mediterranean sea. Mar. Pollut. Bull. 2009, 58, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Al-Bahry, S.N.; Mahmoud, I.Y.; Al-Zadjali, M.; Elshafie, A.; Al-Harthy, A.; Al-Alawi, W. Antibiotic resistant bacteria as bio-indicator of polluted effluent in the green turtles, Chelonia mydas in Oman. Mar. Environ. Res. 2011, 71, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahasan, M.S.; Picard, J.; Elliott, L.; Kinobe, R.; Owens, L.; Ariel, E. Evidence of antibiotic resistance in enterobacteriales isolated from green sea turtles, Chelonia mydas on the great barrier reef. Mar. Pollut. Bull. 2017, 120, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Savoca, D.; Arculeo, M.; Barreca, S.; Buscemi, S.; Caracappa, S.; Gentile, A.; Persichetti, M.F.; Pace, A. Chasing phthalates in tissues of marine turtles from the mediterranean sea. Mar. Pollut. Bull. 2018, 127, 165–169. [Google Scholar] [CrossRef]
- Pace, A.; Rinaldi, L.; Ianniello, D.; Borrelli, L.; Cringoli, G.; Fioretti, A.; Hochscheid, S.; Dipineto, L. Gastrointestinal investigation of parasites and enterobacteriaceae in loggerhead sea turtles from italian coasts. BMC Vet. Res. 2019, 15, 370. [Google Scholar] [CrossRef]
- Pace, A.; Dipineto, L.; Fioretti, A.; Hochscheid, S. Loggerhead sea turtles as sentinels in the western mediterranean: Antibiotic resistance and environment-related modifications of gram-negative bacteria. Mar. Pollut. Bull. 2019, 149, 110575. [Google Scholar] [CrossRef]
- Caracappa, S.; Persichetti, M.F.; Piazza, A.; Caracappa, G.; Gentile, A.; Marineo, S.; Crucitti, D.; Arculeo, M. Incidental catch of loggerhead sea turtles (Caretta caretta) along the sicilian coasts by longline fishery. PeerJ 2018, 6, e5392. [Google Scholar] [CrossRef] [Green Version]
- Pulcinella, J.; Bonanomi, S.; Colombelli, A.; Fortuna, C.M.; Moro, F.; Lucchetti, A.; Sala, A. Bycatch of loggerhead turtle (Caretta caretta) in the italian adriatic midwater pair trawl fishery. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Gambino, D.; Persichetti, M.F.; Gentile, A.; Arculeo, M.; Visconti, G.; Curro, V.; Caracappa, G.; Crucitti, D.; Piazza, A.; Mancianti, F.; et al. First data on microflora of loggerhead sea turtle (Caretta caretta) nests from the coastlines of sicily. Biol. Open 2020, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caracappa, S.; Pisciotta, A.; Persichetti, M.F.; Caracappa, G.; Alduina, R.; Arculeo, M. Nonmodal scutes patterns in the loggerhead sea turtle (Caretta caretta): A possible epigenetic effect? Can. J. Zool. 2016, 94, 379–383. [Google Scholar] [CrossRef]
- Fichi, G.; Cardeti, G.; Cersini, A.; Mancusi, C.; Guarducci, M.; Di Guardo, G.; Terracciano, G. Bacterial and viral pathogens detected in sea turtles stranded along the coast of Tuscany, Italy. Vet. Microbiol. 2016, 185, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, D.; Walsh, F. Antibiotic resistance genes across a wide variety of metagenomes. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef]
- Cardoso-Brito, V.; Raposo, A.C.S.; Pires, T.T.; Pinna, M.H.; Oria, A.P. Conjunctival bacterial flora and antimicrobial susceptibility of captive and free-living sea turtles in brazil. Vet. Ophthalmol. 2019, 22, 246–255. [Google Scholar] [CrossRef]
- Delli Paoli Carini, A.; Ariel, E.; Picard, J.; Elliott, L. Antibiotic resistant bacterial isolates from captive green turtles and in vitro sensitivity to bacteriophages. Int. J. Microbiol. 2017, 2017, 5798161. [Google Scholar] [CrossRef] [Green Version]
- Zavala-Norzagaray, A.A.; Aguirre, A.A.; Velazquez-Roman, J.; Flores-Villasenor, H.; Leon-Sicairos, N.; Ley-Quinonez, C.P.; Hernandez-Diaz Lde, J.; Canizalez-Roman, A. Isolation, characterization, and antibiotic resistance of Vibrio spp. in sea turtles from northwestern mexico. Front. Microbiol. 2015, 6, 635. [Google Scholar] [CrossRef] [Green Version]
- Al-Bahry, S.; Mahmoud, I.; Elshafie, A.; Al-Harthy, A.; Al-Ghafri, S.; Al-Amri, I.; Alkindi, A. Bacterial flora and antibiotic resistance from eggs of green turtles Chelonia mydas: An indication of polluted effluents. Mar. Pollut. Bull. 2009, 58, 720–725. [Google Scholar] [CrossRef]
- Chuen-Im, T.; Suriyant, D.; Sawetsuwannakun, K.; Kitkumthorn, N. The occurrence of vibrionaceae, staphylococcaceae, and enterobacteriaceae in green turtle Chelonia mydas rearing seawater. J. Aquat. Anim. Health 2019, 31, 303–310. [Google Scholar] [CrossRef]
- Di Ianni, F.; Dodi, P.L.; Cabassi, C.S.; Pelizzone, I.; Sala, A.; Cavirani, S.; Parmigiani, E.; Quintavalla, F.; Taddei, S. Conjunctival flora of clinically normal and diseased turtles and tortoises. BMC Vet. Res. 2015, 11, 91. [Google Scholar] [CrossRef] [Green Version]
- Dumontet, S.; Krovacek, K.; Svenson, S.B.; Pasquale, V.; Baloda, S.B.; Figliuolo, G. Prevalence and diversity of Aeromonas and Vibrio spp. in coastal waters of southern Italy. Comp. Immunol. Microbiol. Infect. Dis. 2000, 23, 53–72. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The ocean as a global reservoir of antibiotic resistance genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-X.; Zhang, T.; Fang, H.H.P. Antibiotic resistance genes in water environment. Appl. Microbiol. Biot. 2009, 82, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Pedersen, K.; Larsen, J.L. Occurrence of antimicrobial resistance in fish-pathogenic and environmental bacteria associated with four danish rainbow trout farms. Appl. Environ. Microbiol. 2000, 66, 4908–4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, A.F.C.; Zhang, L.; Balfour, A.J.; Garside, R.; Hawkey, P.M.; Murray, A.K.; Ukoumunne, O.C.; Gaze, W.H. Exposure to and colonisation by antibiotic-resistant E. coli in UK coastal water users: Environmental surveillance, exposure assessment, and epidemiological study (beach bum survey). Environ. Int. 2018, 114, 326–333. [Google Scholar] [CrossRef]
- Aguirre, A.A.; Gardner, S.C.; Marsh, J.C.; Delgado, S.G.; Limpus, C.J.; Nichols, W.J. Hazards associated with the consumption of sea turtle meat and eggs: A review for health care workers and the general public. EcoHealth 2006, 3, 141–153. [Google Scholar] [CrossRef]
- Sultan, I.; Rahman, S.; Jan, A.T.; Siddiqui, M.T.; Mondal, A.H.; Haq, Q.M.R. Antibiotics, resistome and resistance mechanisms: A bacterial perspective. Front. Microbiol. 2018, 9, 2066. [Google Scholar] [CrossRef] [Green Version]
- Segawa, T.; Takeuchi, N.; Rivera, A.; Yamada, A.; Yoshimura, Y.; Barcaza, G.; Shinbori, K.; Motoyama, H.; Kohshima, S.; Ushida, K. Distribution of antibiotic resistance genes in glacier environments. Environ. Microbiol. Rep. 2013, 5, 127–134. [Google Scholar] [CrossRef]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.L.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef]
- Santestevan, N.A.; de Angelis Zvoboda, D.; Prichula, J.; Pereira, R.I.; Wachholz, G.R.; Cardoso, L.A.; de Moura, T.M.; Medeiros, A.W.; de Amorin, D.B.; Tavares, M.; et al. Antimicrobial resistance and virulence factor gene profiles of Enterococcus spp. isolates from wild Arctocephalus australis (south american fur seal) and Arctocephalus tropicalis (subantarctic fur seal). World J. Microbiol. Biotechnol. 2015, 31, 1935–1946. [Google Scholar] [CrossRef]
- Prichula, J.; Pereira, R.I.; Wachholz, G.R.; Cardoso, L.A.; Tolfo, N.C.; Santestevan, N.A.; Medeiros, A.W.; Tavares, M.; Frazzon, J.; d’Azevedo, P.A.; et al. Resistance to antimicrobial agents among enterococci isolated from fecal samples of wild marine species in the southern coast of brazil. Mar. Pollut. Bull. 2016, 105, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Al-Bahry, S.N.; Al-Zadjali, M.A.; Mahmoud, I.Y.; Elshafie, A.E. Biomonitoring marine habitats in reference to antibiotic resistant bacteria and ampicillin resistance determinants from oviductal fluid of the nesting green sea turtle, Chelonia mydas. Chemosphere 2012, 87, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Burgmann, H.; Sorum, H.; Norstrom, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Arizza, V.; Vecchioni, L.; Caracappa, S.; Sciurba, G.; Berlinghieri, F.; Gentile, A.; Persichetti, M.F.; Arculeo, M.; Alduina, R. New insights into the gut microbiome in loggerhead sea turtles Caretta caretta stranded on the mediterranean coast. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Galluzzo, P.; Buffa, P.G.; Carlino, E.; Spezia, O.; Alduina, R. Comparison of antibiotic resistance profile and biofilm production of Staphylococcus aureus isolates derived from human specimens and animal-derived samples. Antibiotics 2019, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Gaglio, S.; Galluzzo, P.; Cascone, G.; Piraino, C.; Lo Presti, V.D.; Alduina, R. Antibiotic resistance profiling, analysis of virulence aspects and molecular genotyping of Staphylococcus aureus isolated in Sicily, Italy. Foodborne Pathog. Dis. 2018, 15, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, A.; Eckert, E.M.; D’Urso, S.; Bertoni, R.; Gillan, D.C.; Wattiez, R.; Corno, G. Co-occurrence of integrase 1, antibiotic and heavy metal resistance genes in municipal wastewater treatment plants. Water Res. 2016, 94, 208–214. [Google Scholar] [CrossRef]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef]
- Bibbal, D.; Dupouy, V.; Ferre, J.P.; Toutain, P.L.; Fayet, O.; Prere, M.F.; Bousquet-Melou, A. Impact of three ampicillin dosage regimens on selection of ampicillin resistance in enterobacteriaceae and excretion of blatem genes in swine feces. Appl. Environ. Microbiol. 2007, 73, 4785–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, E.; Jofre, J.; Balcazar, J.L. Prevalence of antibiotic resistance genes and bacterial community composition in a river influenced by a wastewater treatment plant. PLoS ONE 2013, 8, e78906. [Google Scholar] [CrossRef] [PubMed]
- Marti, E.; Balcazar, J.L. Real-time pcr assays for quantification of qnr genes in environmental water samples and chicken feces. Appl. Environ. Microbiol. 2013, 79, 1743–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, R.; Kim, S.C.; Carlson, K.H.; Pruden, A. Effect of river landscape on the sediment concentrations of antibiotics and corresponding antibiotic resistance genes (arg). Water Res. 2006, 40, 2427–2435. [Google Scholar] [CrossRef]
- Roosa, S.; Wattiez, R.; Prygiel, E.; Lesven, L.; Billon, G.; Gillan, D.C. Bacterial metal resistance genes and metal bioavailability in contaminated sediments. Environ. Pollut. 2014, 189, 143–151. [Google Scholar] [CrossRef]
- Luo, Y.; Mao, D.; Rysz, M.; Zhou, Q.; Zhang, H.; Xu, L.; Alvarez, P.J.J. Trends in antibiotic resistance genes occurrence in the haihe river, China. Environ. Sci. Technol. 2010, 44, 7220–7225. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | blaTEM | qnrS | sulII | tetA | blaCTXM | czcA | arsB | int1 |
|---|---|---|---|---|---|---|---|---|
| Dead Turtle (n = 3) | 2 | ND 1 | 1 | ND | ND | 1 | ND | 2 |
| Live Turtle (n = 3) | 2 | 1 | 2 | 1 | ND | 1 | ND | 3 |
| Sand (n = 3) | 3 | ND | ND | ND | ND | 3 | ND | 3 |
| Eggshell (n = 3) | 2 | ND | 1 | ND | ND | 1 | 2 | 3 |
| Yolk (n = 2) | 2 | ND | ND | ND | ND | ND | ND | 1 |
| Seawater (n = 2) | 2 | ND | 1 | ND | ND | 2 | ND | 2 |
| Sample | Location | Bacterial Isolates |
|---|---|---|
| Bottom sand | Linosa | Staphylococcus aureus; Aeromonas hydrophyla/cavie |
| Top sand | Bacillus sp.; Streptococcus sp. | |
| Hatched egg | Bacillus sp.; Streptococcus sp. | |
| Unhatched egg | Aeromonas hydrophyla/cavie; Citrobacter freundii; Bacillus sp. | |
| Yolk | Aeromonas hydrophyla/cavie; Citrobacter freundii; | |
| Seawater | South coast | Enterobacter sp.; Vibrio alginolyticus; Aeromonas sp. |
| North coast | Vibrio neocaledonicus; Vibrio sp. |
| Sample | Sex | CCL 1 | Weight (Kg) | Stranding Location | Isolate |
|---|---|---|---|---|---|
| Cloacal swab | F | 68 | 29 | Siracusa (SR) | Aeromonas spp. |
| F | 41 | 18 | Citrobacter spp. | ||
| F | 38 | 19 | Lipari (ME) | Escherichia coli | |
| - | 13 | 4 | Augusta (SR) | Aeromonas spp. | |
| - | 25 | 8 | Enterobacter spp. | ||
| - | 30 | 16 | Aeromonas spp. | ||
| M | 25 | 11 | Aeromonas spp. | ||
| - | 46 | 23 | Milazzo (ME) | Aeromonas spp. | |
| Oral swab | F | 54 | 23 | Milazzo (ME) | Pseudomonas spp. |
| Citrobacter spp. | |||||
| Aeromonas spp. | |||||
| F | 38 | 14 | Lipari (ME) | Aeromonas spp. | |
| M | 54 | 23 | Palermo (PA) | Aeromonas spp. | |
| M | 48 | 26 | Filicudi (ME) | Aeromonas spp. | |
| Skin swab | F | 31 | 10 | Ustica (PA) | Klebsiella spp. |
| - | 28 | 10 | Porticello (PA) | Aeromonas spp. | |
| Organs | F | 51 | 34 | Milazzo (ME) | Aeromonas spp. |
| Enterococcus faecium | |||||
| - | 27 | 4 | Aeromonas spp. | ||
| - | 38 | 35 | Proteus vulgaris | ||
| F | 68 | 29 | Siracusa (SR) | Aeromonas spp. | |
| F | 44 | 31 | Terrasini (PA) | Aeromonas spp. | |
| F | 48 | 21 | Aeromonas spp. | ||
| Citrobacter spp. | |||||
| M | 43 | 19 | Messina (ME) | Enterobacter spp. | |
| M | 23 | 3.5 | Enterobacter spp. |
| Target Name | Primer Sequence (5′-3′) | Annealing Temperature | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| 16SrDNA | CGGTGAATACGTTCYCGG GGHTACCTTGTTACGACTT | 55 | 142 | [49] |
| tetA | GCTACATCCTGCTTGCCTTC CATAGATCGCCGTGAAGAGG | 64 | 210 | [50] |
| blaTEM | TTCCTGTTTTTGCTCACCCAG CTCAAGGATCTTACCGCTGTTG | 60 | 112 | [51] |
| blaCTXM | CTATGGCACCACCAACGATA ACGGCTTTCTGCCTTAGGTT | 60 | 103 | [52] |
| qnrS | GACGTGCTAACTTGCGTGAT TGGCATTGTTGGAAACTTG | 62 | 118 | [53] |
| sulII | TCCGGTGGAGGCCGGTATCTGG CGGGAATGCCATCTGCCTTGAG | 60 | 191 | [54] |
| czcA | TCGACGGBGCCGTGGTSMTBGTCGAGAA GTVAWSGCCAKCGGVBGGAACA | 63 | 232 | [55] |
| arsB | GTSAARCCSTTYTCGATGGC GCRAASGCSAHSAYCATGAT | 56.5 | 226 | [55] |
| int1 | GGCTTCGTGATGCCTGCTT CATTCCTGGCCGTGGTTCT | 59 | 148 | [56] |
| 16S rDNA | CCTACGGGNBGCASCAG GACTACNVGGGTATCTAATCC | 55 | 464 | [57] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alduina, R.; Gambino, D.; Presentato, A.; Gentile, A.; Sucato, A.; Savoca, D.; Filippello, S.; Visconti, G.; Caracappa, G.; Vicari, D.; et al. Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea? Antibiotics 2020, 9, 116. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030116
Alduina R, Gambino D, Presentato A, Gentile A, Sucato A, Savoca D, Filippello S, Visconti G, Caracappa G, Vicari D, et al. Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea? Antibiotics. 2020; 9(3):116. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030116
Chicago/Turabian StyleAlduina, Rosa, Delia Gambino, Alessandro Presentato, Antonino Gentile, Arianna Sucato, Dario Savoca, Serena Filippello, Giulia Visconti, Giulia Caracappa, Domenico Vicari, and et al. 2020. "Is Caretta Caretta a Carrier of Antibiotic Resistance in the Mediterranean Sea?" Antibiotics 9, no. 3: 116. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9030116