Chalcones Isolated from Arrabidaea brachypoda Flowers as Inhibitors of NorA and MepA Multidrug Efflux Pumps of Staphylococcus aureus

 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
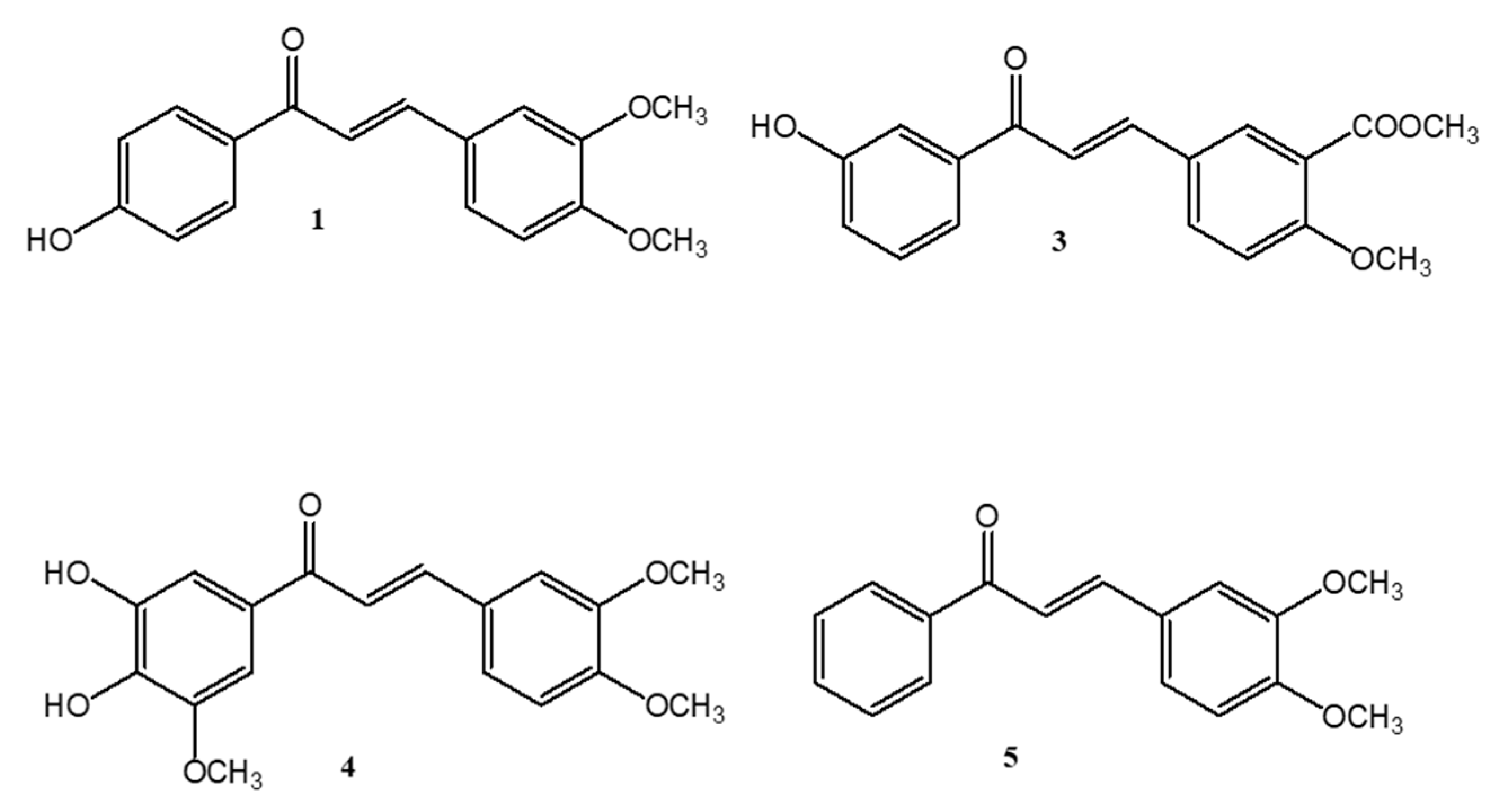
2.1. Compound Identification and Evaluation of the Antimicrobial Activity
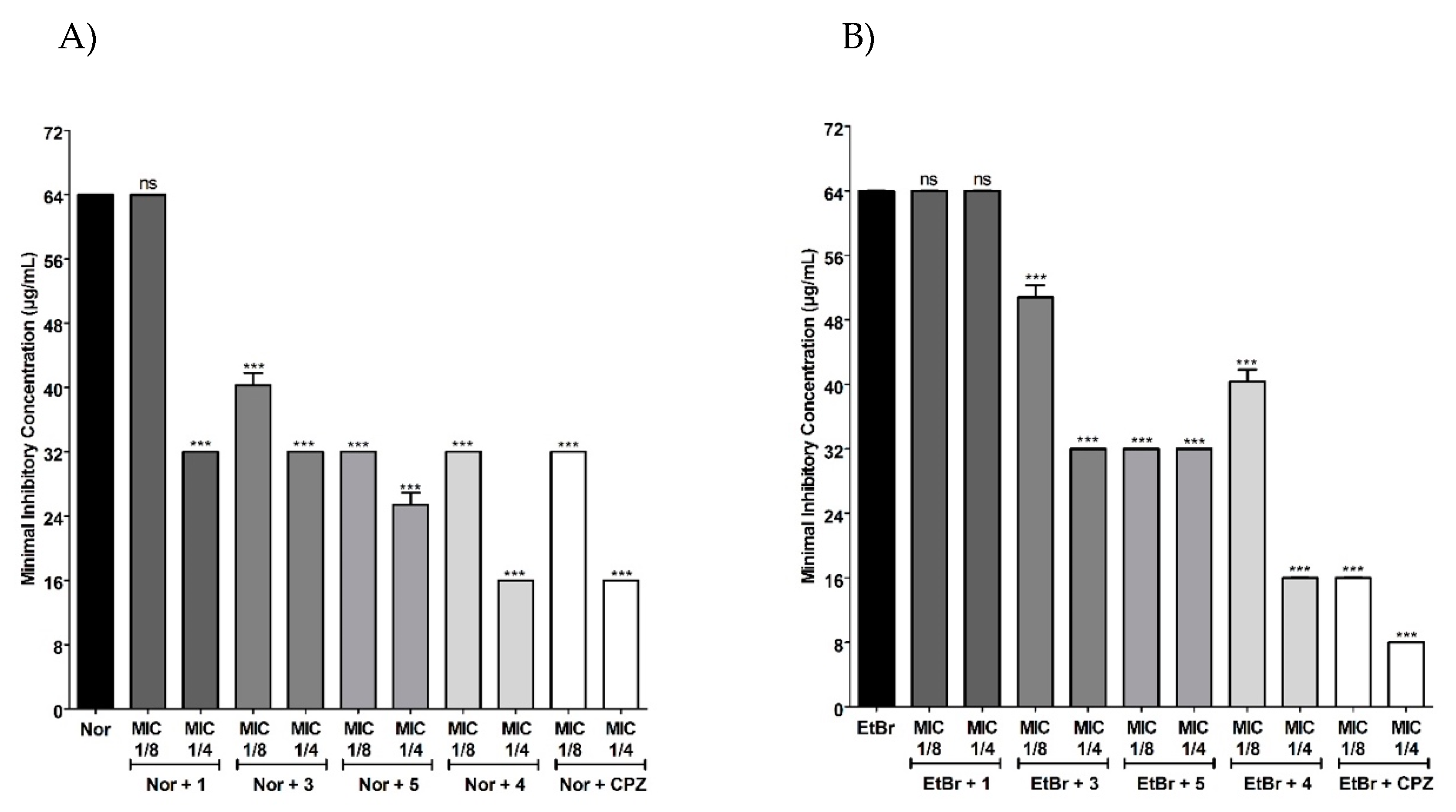
2.2. Evaluation of the Modulating Effect of Isolated Chalcones on the Resistance to Norfloxacin
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of the Ethanolic Extract and Obtaining the Diclorometanic Fraction
3.3. Isolation and Identification of Chalcones
3.4. Compound Identification
3.5. Strains and Drugs
3.6. Evaluation of the Antimicrobial Activity Assays
3.7. Modulation of the Antibiotic Resistance Assays
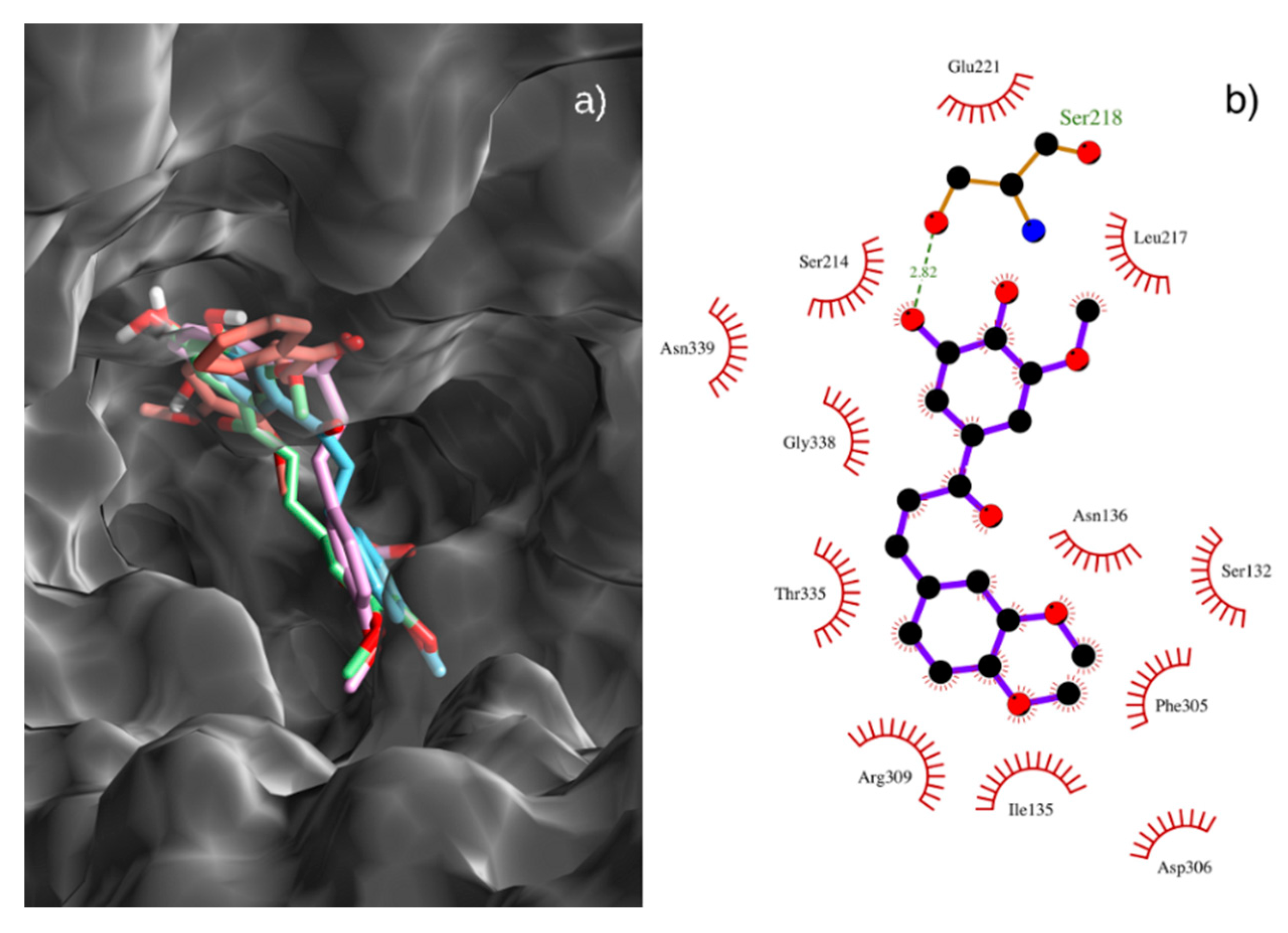
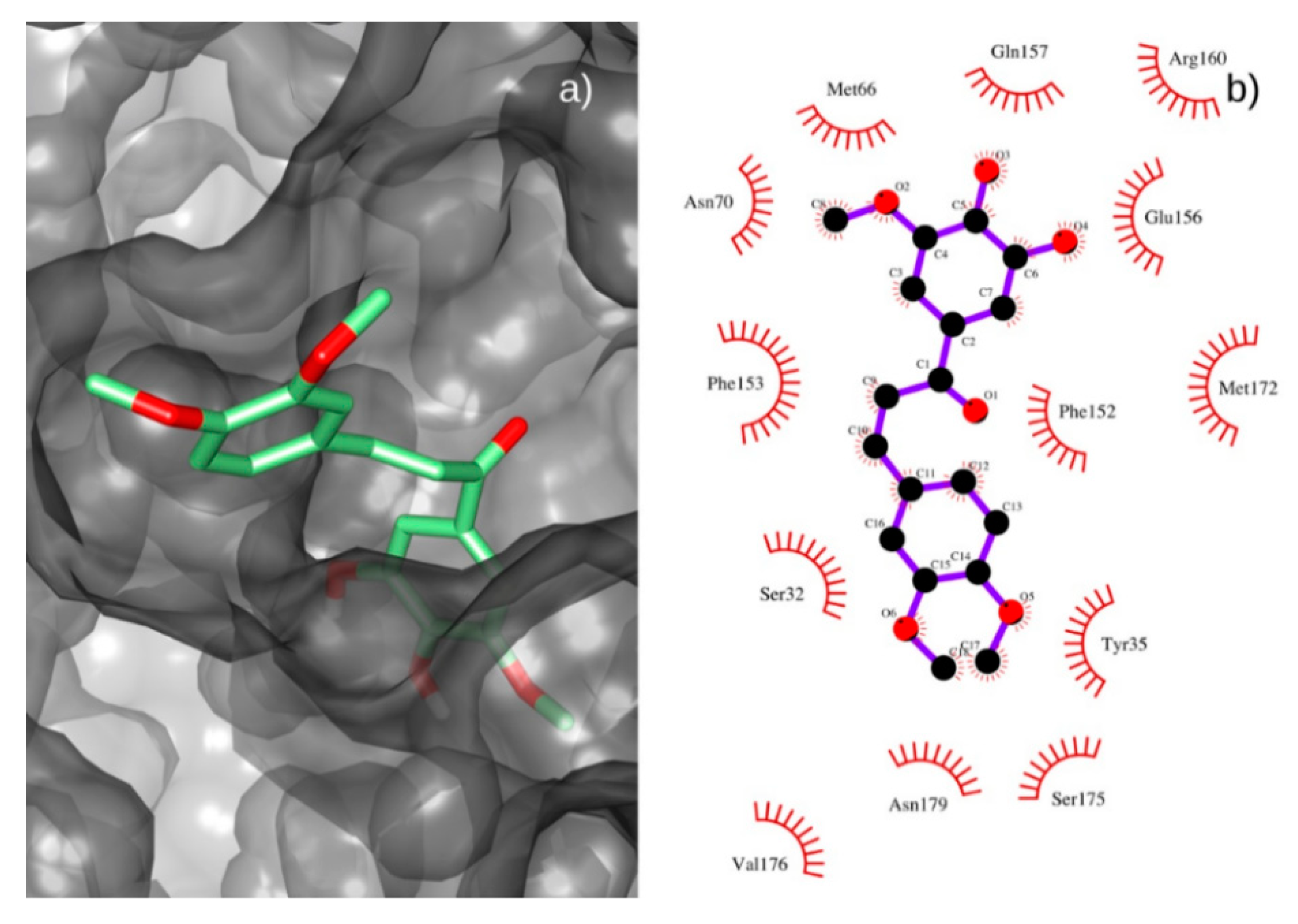
3.8. Docking Procedure
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brown, E.D.; Wright, G.D. Antibacterial drug discovery in the resistance era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Lira, A.C.; Lima, L.F.A.; Coutinho, H.D.M.; de Siqueira, J.P., Jr.; Barreto, H.M. Antimicrobial resistance in staphylococci isolated from canine pyoderma. Commun. Sci. 2012, 3, 181–185. [Google Scholar] [CrossRef]
- Chua, K.Y.L.; Howden, B.P.; Jiang, J.H.; Stinear, T.; Peleg, A.Y. Population genetics and the evolution of virulence in Staphylococcus aureus Infection. Genet. Evol. 2014, 21, 554–562. [Google Scholar] [CrossRef]
- Botelho, A.M.N.; Costa, M.O.C.; Moustafa, A.M.; Beltrame, C.O.; Ferreira, F.A.; Côrtes, M.F.; Costa, B.S.S.; Silva, D.N.S.; Bandeira, P.T.; Lima, N.C.B.; et al. Local diversification of Methicillin-Resistant Staphylococcus aureus ST239 in South America after its rapid worldwide dissemination. Front. Microbiol. 2019, 10, 82. [Google Scholar] [CrossRef]
- Alcalde-Rico, M.; Olivares-Pacheco, J.; Alvarez-Ortega, C.; Camara, M.; Martínez, J.L. Role of the multidrug resistance efflux pump MexCD-OprJ in the Pseudomonas aeruginosa quorum sensing response. Front. Microb. 2018, 9, 2752. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Cui, L.; Kuroda, M.; Ito, T. The emergence and evolution of methicillin-resistant Staphylococcus aureus. Trends Icrobial. 2001, 2001 9, 486–493. [Google Scholar] [CrossRef]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Peterson, E.; Kaur, P. Antibiotic resistance mechanisms in bacteria: Relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Front. Microb. 2018, 9, 2928. [Google Scholar] [CrossRef]
- Du, D.; Wang, K.X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.V.; Piddock, L.J.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Schindler, B.D.; Kaatz, G.W. Multidrug efflux pumps of Gram-positive bacteria. Drug Resistance Updat. 2016, 27, 1–13. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Blanco, P.; Alcalde-Rico, M.; Corona, F.; Reales-Calderón, J.A.; Sánchez, M.B.; Martínez, J.L. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resistance Updat. 2016, 28, 13–27. [Google Scholar] [CrossRef]
- Hassanzadeh, S.; Mashhadi, R.; Yousefi, M.; Askari, E.; Saniei, M.; Pourmand, M.R. Frequency of efflux pump genes mediating ciprofloxacin and antiseptic resistance in Methicillin-Resistant Staphylococcus aureus isolates. Microb. Pathog. 2017, 111, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R. Antibiotic effectiveness. Science 2014, 345, 1299–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachino, J.I.; Jin, W.; Kimura, K.; Kurosaki, H.; Sato, A.; Arakawa, Y. Sulfamoyl heteroarylcarboxylic acids as promising metallo-β-lactamase inhibitors for controlling bacterial carbapenem resistance. Mbio 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinoza, J.; Urzúa, A.; Sanhueza, L.; Walter, M.; Fincheira, P.; Muñoz, P.; Mendoza, L.; Wilkens, M. Essential oil, extracts, and sesquiterpenes obtained from the heartwood of Pilgerodendron uviferum act as potential inhibitors of the Staphylococcus aureus NorA multidrug efflux pump. Front. Microbiol. 2019, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Coêlho, M.L.; Ferreira, J.H.L.; de Siqueira Júnior, J.P.; Kaatz, G.W.; Barreto, H.M.; de Carvalho Melo Cavalcante, A.A. Inhibition of the NorA multi-drug transporter by oxygenated monoterpenes. Microb. Pathog. 2016, 99, 173–177. [Google Scholar] [CrossRef]
- Lorenzi, H.; Souza, H.M. Plantas Ornamentais no Brasil: Arbustivas, Herbáceas e Trepadeiras, 3rd ed.; Instituto Plantarum de Estudos da Flora, Ed.; Instituto Plantarum de Estudos da Flora: São Paulo, Brasil, 2001. [Google Scholar]
- Alcerito, T.; Barbo, F.E.; Negri, G.; Santos, D.Y.A.C.; Meda, C.I.; Young, M.C.M.; Chávez, D.; Blatt, C.T.T. Foliar epicuticular wax of Arrabidaea brachypoda: Flavonoids and antifungal activity. Biochem. Syst. Ecol. 2002, 30, 677–683. [Google Scholar] [CrossRef]
- Rodrigues, E.; Mendes, F.; Negri, G. Plants indicated by brazilian indians for disturbances of the central nervous system: A bibliographical survey. Cent. Ner. Syst. Agents Med. Chem. 2006, 6, 211–244. [Google Scholar] [CrossRef]
- K Sahu, N.; S Balbhadra, S.; Choudhary, J.; V Kohli, D. Exploring pharmacological significance of chalcone scaffold: A review. Curr. Med. Chem. 2012, 19, 209–225. [Google Scholar] [CrossRef]
- Rocha, V.; Quintino da Rocha, C.; Ferreira Queiroz, E.; Marcourt, L.; Vilegas, W.; Grimaldi, G.; Furrer, P.; Allémann, É.; Wolfender, J.-L.; Soares, M. Antileishmanial activity of dimeric flavonoids isolated from Arrabidaea brachypoda. Molecules 2018, 24, 1. [Google Scholar] [CrossRef] [Green Version]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Anti-cancer chalcones: Structural and molecular target perspectives. Eur. J. Med. Chem. 2015, 98, 69–114. [Google Scholar] [CrossRef]
- Sharma, N.; Mohanakrishnan, D.; Shard, A.; Sharma, A.; Sharma, A.; Sinha, A.; Sahal, D. Stilbene–chalcone hybrids: Design, synthesis, and evaluation as a new class of antimalarial scaffolds that trigger cell death through stage specific apoptosis. J. Med. Chem. 2012, 55, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.M.D.S.; Oliveira, A.B.M.; Leal, A.L.A.B.; Oliveira, F.A.D.A.; Portela, A.L.; Lima Neto, J.D.S.; Siqueira-Júnior, J.P.; Kaatz, G.W.; Rocha, C.Q.; Barreto, H.M. Antimicrobial activity and inhibition of the NorA efflux pump of Staphylococcus aureus by extract and isolated compounds from Arrabidaea brachypoda. Microb. Pathog. 2020, 140, 103935. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.M.; Mytelka, D.S.; Dunwiddie, C.T.; Persinger, C.C.; Munos, B.H.; Lindborg, S.R.; Schacht, A.L. How to improve R&D productivity: The pharmaceutical industry’s grand challenge. Nat. Rev. Drug Discov. 2010, 9, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.K.A.; Fontenelle, R.O.S.; Magalhães, F.E.A.; Bandeira, P.N.; Menezes, S.; Santos, J.E.S.A.; Dos Santos, H. Potential farmacológico de chalconas: Uma breve revisão. Rev. Virtual de Quím. 2018, 10, 1455–1473. [Google Scholar]
- Ramírez Escobedo, M.E.; Barajas Bermúdez, L.; Pérez Berumen, C.; Sáenz Galindo, A.; Silva Belmares, S.Y. Síntesis y actividad biológica de chalconas. Rev. Mex. de Cienc. Farm. 2012, 43, 7–14. [Google Scholar]
- Rao, G.P.; Kumar, P.; Singh, M.; Singh, H.N.; Pandey, O.P. Efficacy of chalcone, hydrazide and oxadiazole derivatives against fungal pathogens of sugarcane. Sugar Cane 1994, 17–22. [Google Scholar]
- Thirunarayanan, G.; Vanangamudi, G. Synthesis, spectral studies, antimicrobial and insect antifeedant activities of some substituted styryl 4′-fluorophenyl ketones. Arabian J. Chem. 2014, 7, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Kachadourian, R.; Day, B.J. Flavonoid-induced glutathione depletion: Potential implications for cancer treatment. Free Radic. Biol. Med. 2006, 41, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Houghton, P.J.; Howes, M.J.; Lee, C.C.; Steventon, G. Uses and abuses of in vitro tests in ethnopharmacology: Visualizing an elephant. J. Ethnopharmacol. 2007, 110, 391–400. [Google Scholar] [CrossRef]
- Kaatz, G.W.; Seo, S.M.; Ruble, C.A. Efflux-mediated fluoroquinolone resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1993, 37, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Markham, P.N.; Westhaus, E.; Klyachko, K.; Johnson, M.E.; Neyfakh, A.A. Multiple novel inhibitors of the NorA multidrug transporter of Staphylococcus aureus multiple novel inhibitors of the NorA multidrug transporter of Staphylococcus aureus. Antimicrob. Agents Chemother. 1999, 43, 2404–2408. [Google Scholar] [CrossRef] [Green Version]
- Thai, K.-M.; Ngo, T.-D.; Phan, T.-V.; Tran, T.-D.; Nguyen, N.-V.; Nguyen, T.-H.; Le, M.-T. Virtual screening for novel Staphylococcus aureus NorA efflux pump inhibitors from natural products. Med. Chem. 2015, 11, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Cirino, I.C.S.; Menezes-Silva, S.M.P.; Silva, H.T.D.; de Souza, E.L.; Siqueira-Júnior, J.P. The Essential oil from Origanum vulgare L. and its individual constituents Carvacrol and Thymol enhance the effect of Tetracycline against Staphylococcus aureus. Chemotherapy 2014, 60, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.M.; de Macedo, E.V.; Oliveira, F.A.A.; Ferreira, J.H.L.; Gutierrez, S.J.C.; Peláez, W.J.; Lima, F.C.A.; de Siqueira Júnior, J.P.; Coutinho, H.D.M.; Kaatz, G.W.; et al. Inhibition of the NorA efflux pump of Staphylococcus aureus by synthetic riparins. J. Appl. Microbiol. 2016, 121, 1312–1322. [Google Scholar] [CrossRef]
- Braga Ribeiro, A.M.; Sousa, J.N.; Costa, L.M.; Oliveira, F.A.A.; dos Santos, R.C.; Silva Nunes, A.S.; da Silva, W.O.; Marques Cordeiro, P.J.; de Sousa Lima Neto, J.; de Siqueira-Júnior, J.P.; et al. Antimicrobial activity of Phyllanthus amarus Schumach. & Thonn and inhibition of the NorA efflux pump of Staphylococcus aureus by Phyllanthin. Microb. Pathog. 2019, 130, 242–246. [Google Scholar] [CrossRef]
- Maia, G.L.A.; Falcão-Silva, V.S.; Aquino, P.G.V.; Araújo-Júnior, J.X.; Tavares, J.F.; Silva, M.S.; Rodrigues, L.C.; Siqueira-Júnior, J.P.; Barbosa-Filho, J.M. Flavonoids from Praxelis clematidea R.M. King and Robinson modulate bacterial drug resistance. Molecules 2011, 16, 4828–4835. [Google Scholar] [CrossRef]
- Gibbons, S.; Udo, E.E. The effect of reserpine, a modulator of multidrug efflux pumps, on the in vitro activity of tetracycline against clinical isolates of methicillin resistant Staphylococcus aureus (MRSA) possessing the tet(K) determinant. Phytother. Res. 2000, 14, 139–140. [Google Scholar] [CrossRef]
- Ross, J.I.; Farrell, A.M.; Eady, E.A.; Cove, J.H.; Cunliffe, W.J. Characterisation and molecular cloning of the novel macrolide-streptogramin B resistance determinant from Staphylococcus epidermidis. J. Antimicrob. Chemother. 1989, 24, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Kaatz, G.W.; McAleese, F.; Seo, S.M. Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob. Agents Chemother. 2005, 49, 1857–1864. [Google Scholar] [CrossRef] [Green Version]
- Tennent, J.M.; Lyon, B.R.; Midgley, M.; Jones, G.; Purewal, A.S.; Skurray, R.A. Physical and Biochemical Characterization of the qacA gene encoding antiseptic and disinfectant resistance in Staphylococcus aureus. Microbiology 1989, 135, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Littlejohn, T.G.; DiBerardino, D.; Messerotti, L.J.; Spiers, S.J.; Skurray, R.A. Structure and evolution of a family of genes encoding antiseptic and disinfectant resistance in Staphylococcus aureus. Gene 1991, 101, 59–66. [Google Scholar] [CrossRef]
- CLSI. Clinical and Laboratory Standards Institute. Document M07-A9, Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard—Ninth Edition; CLSI: Wayne, NJ, USA, 2012; Volume 32, ISBN 1562387839. [Google Scholar]
- Neyfakh, A.A.; Borsch, C.M.; Kaatz, G.W. Fluoroquinolone resistance protein NorA of Staphylococcus aureus is a multidrug efflux transporter. Antimicrob. Agents Chemother. 1993, 37, 128–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hospital, A.; Andrio, P.; Fenollosa, C.; Cicin-Sain, D.; Orozco, M.; Gelpí, J.L. MDWeb and MDMoby: An integrated web-based platform for molecular dynamics simulations. Bioinformatics 2012, 28, 1278–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 456–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chalcones (Exact Mass) | m/z; MSn (% Base Peak) |
|---|---|
| 283.0980 | 283[M − H]−; MS2[268]: 253(12), 239(100) |
| 311.0919 | 311[M − H]−; MS2[253]: 28(100), 269(100) |
| 329.1025 | 329[M − H]−; MS2[269]: 135(100); MS3[135]: 193(100) |
| 267.1021 | 267[M − H]−; MS2[252]: 221(100) |
| Compound | Molecular Formula | Acurate Mass (amu) | Exact Mass | [M − H]− | Δm (ppm) |
|---|---|---|---|---|---|
| 1 | C17H16O4 | 283.0958 | 283.0980 | 283 | 2.2 |
| 3 | C18H16O5 | 311.0918 | 311.0919 | 311 | 1.0 |
| 4 | C18H18O6 | 329.1028 | 329.1025 | 329 | −3.0 |
| 5 | C17H16O3 | 267.1019 | 267.1021 | 267 | 2.0 |
| MIC (μg/mL) | ||||
|---|---|---|---|---|
| Microbial Strains | 1 | 3 | 4 | 5 |
| Staphylococcus aureus ATCC 25923 | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus IS-58 | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus RN-4220 | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus SA1199-B | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus K2068 | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus K4414 | 1024 | 1024 | 1024 | 1024 |
| Staphylococcus aureus K4100 | ≥1024 | ≥1024 | ≥1024 | ≥1024 |
| Efflux Pump Superfamily | Strain (Drug) | MIC (μg/mL) | ||
|---|---|---|---|---|
| Antibiotic/EtBr Alone | + Chalcone 4 (MIC 1/8) | + Chalcone 4 (MIC 1/4) | ||
| TetK (MFS) | IS-58 (Tet) | 64 | 64 (0,0) * | 64 (0,0) |
| IS-58 (EtBr) | 16 | 8 (2.0) | 4 (4.0) | |
| MsrA (ABC) | RN4220 (Ery) | 32 | 32 (0,0) | 32 (0,0) |
| RN4220 (EtBr) | 32 | 4 (8.0) | 4 (8.0) | |
| MepA (MATE) | K2068 (Nor) | 4 | 2 (2.0) | 1 (4.0) |
| K2068 (EtBr) | 8 | 4 (2.0) | 4 (2.0) | |
| QacA/B (MFS) | K4414 (BrEt) | 32 | 32 (0,0) | 16 (2.0) |
| QacC (SMR) | K4100 (BrEt) | 128 | 128 (0,0) | 128 (0,0) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rezende-Júnior, L.M.; Andrade, L.M.d.S.; Leal, A.L.A.B.; Mesquita, A.B.d.S.; Santos, A.L.P.d.A.d.; Neto, J.d.S.L.; Siqueira-Júnior, J.P.; Nogueira, C.E.S.; Kaatz, G.W.; Coutinho, H.D.M.; et al. Chalcones Isolated from Arrabidaea brachypoda Flowers as Inhibitors of NorA and MepA Multidrug Efflux Pumps of Staphylococcus aureus. Antibiotics 2020, 9, 351. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9060351
Rezende-Júnior LM, Andrade LMdS, Leal ALAB, Mesquita ABdS, Santos ALPdAd, Neto JdSL, Siqueira-Júnior JP, Nogueira CES, Kaatz GW, Coutinho HDM, et al. Chalcones Isolated from Arrabidaea brachypoda Flowers as Inhibitors of NorA and MepA Multidrug Efflux Pumps of Staphylococcus aureus. Antibiotics. 2020; 9(6):351. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9060351
Chicago/Turabian StyleRezende-Júnior, Luís Mário, Leila Maria de Sousa Andrade, Antonio Linkoln Alves Borges Leal, Avilnete Belem de Souza Mesquita, Ana Lurdes Portela de Araújo dos Santos, José de Sousa Lima Neto, José Pinto Siqueira-Júnior, Carlos Emídio Sampaio Nogueira, Glenn William Kaatz, Henrique Douglas Melo Coutinho, and et al. 2020. "Chalcones Isolated from Arrabidaea brachypoda Flowers as Inhibitors of NorA and MepA Multidrug Efflux Pumps of Staphylococcus aureus" Antibiotics 9, no. 6: 351. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9060351