Novel Antimicrobial Peptide from Temporin L in The Treatment of Staphylococcus pseudintermedius and Malassezia pachydermatis in Polymicrobial Inter-Kingdom Infection

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility of M. pachydermatis
2.2. Resistance Acquisition Tests to Peptide 8
2.3. Antimicrobial Susceptibility of S. pseudintermedius
2.4. Synergistic Study
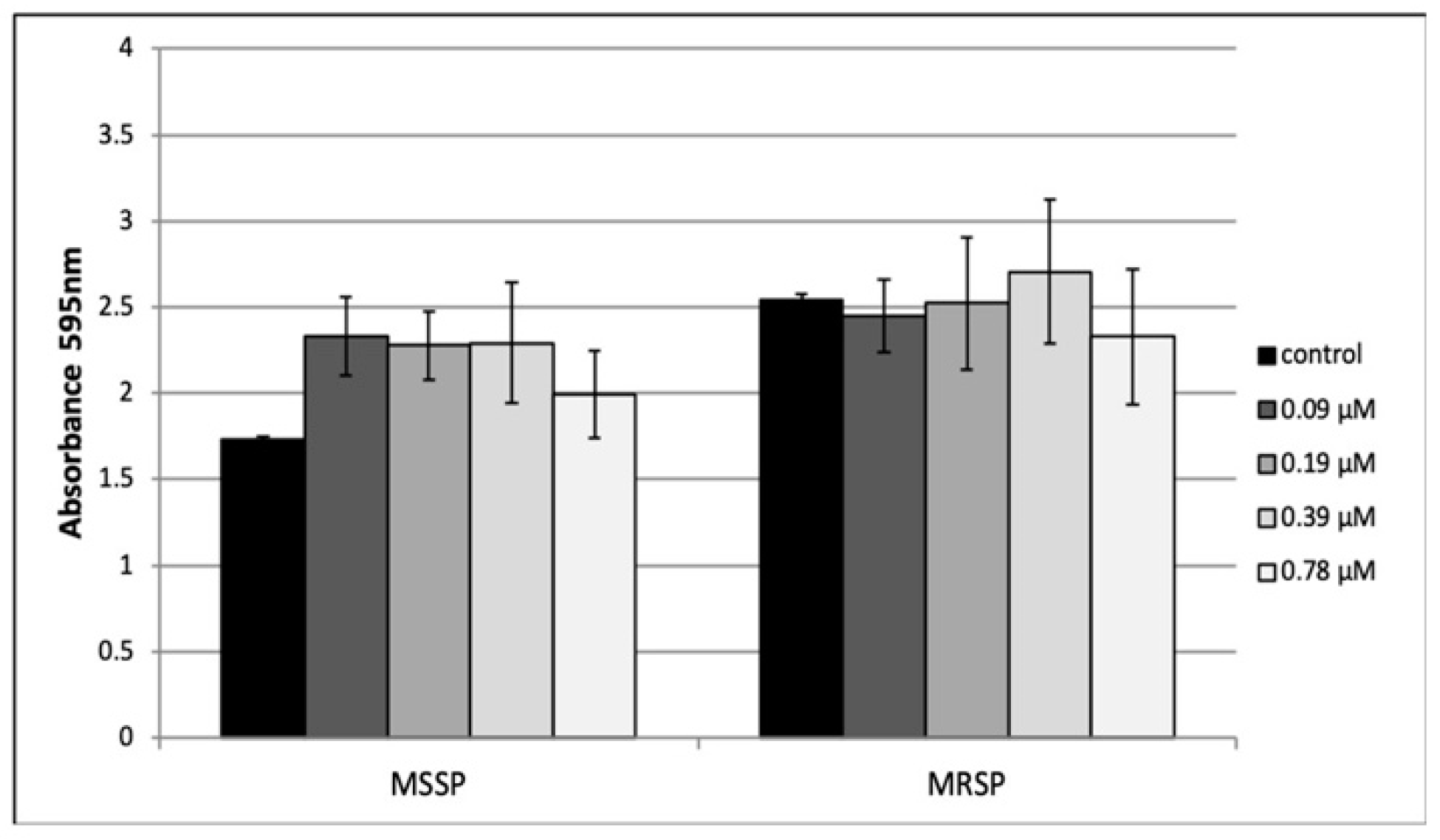
2.5. Effect of Peptide 8 on Biofilm Formation
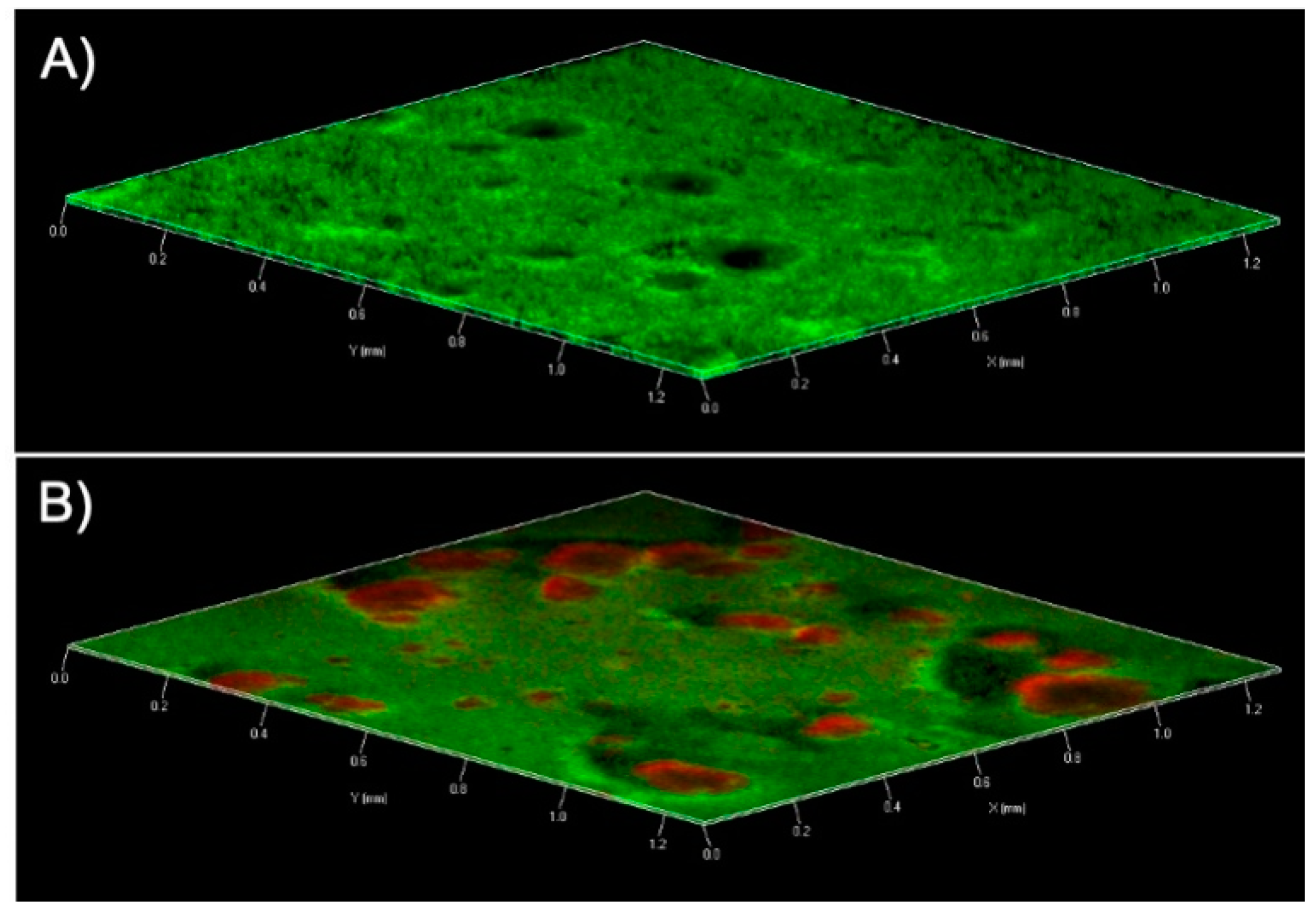
2.6. Effect of Peptide 8 on Mature Biofilm
2.7. Influence of Peptide 8 on MecA Gene Expression
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Peptides Synthesis
4.3. Microbial Strains and Culture
4.4. Molecular Analysis
4.5. Resistance Acquisition Tests to Peptide 8
4.6. Antimicrobial Activity Assay
4.7. Killing Rate
4.8. Checkerboard Method
4.9. Effect of Peptide 8 on Biofilm Formation
4.10. Effect of Peptide 8 on Mature Biofilm Biomass
4.11. Quantitation of Metabolic Activity of Mature Biofilm by XTT Assay
4.12. Confocal Laser Scanning Microscopy (CLSM)
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Peleg, A.Y.; Hogan, D.A.; Mylonakis, E. Medically important bacterial-fungal interactions. Nat. Rev. Microbiol. 2010, 8, 340–349. [Google Scholar] [CrossRef]
- Bond, R. Superficial veterinary mycoses. Clin. Dermatol. 2010, 28, 226–236. [Google Scholar] [CrossRef]
- Ilahi, A.; Hadrich, I.; Goudjil, S.; Kongolo, G.; Chazal, C.; Léké, A.; Ayadi, A.; Chouaki, T.; Ranque, S. Molecular epidemiology of a Malassezia pachydermatis neonatal unit outbreak. Med. Mycol. 2018, 56, 69–77. [Google Scholar] [CrossRef]
- Chryssanthou, E.; Broberger, U.; Petrini, B. Malassezia pachydermatis fungaemia in a neonatal intensive care unit. Acta Paediatr. 2001, 90, 323–327. [Google Scholar] [CrossRef]
- Al-Sweih, N.; Ahmad, S.; Joseph, L.; Khan, S.; Khan, Z. Malassezia pachydermatis fungemia in a preterm neonate resistant to fluconazole and flucytosine. Med. Mycol. Case Rep. 2014, 5, 9–11. [Google Scholar] [CrossRef]
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253-e52. [Google Scholar] [CrossRef]
- Fungwithaya, P.; Brikshavana, P.; Chanchaithong, P.; Prapasarakul, N. Distribution of methicillin-resistant coagulase- positive staphylococci (MRCoPS) in a surgical unit and cystotomy operation sites in a veterinary teaching hospital. J. Vet. Med. Sci. 2017, 79, 359–365. [Google Scholar] [CrossRef]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Limatola, A.; Di Maro, S.; Merlino, F.; Mangoni, M.L.; Luca, V.; Di Grazia, A.; Gatti, S.; et al. Novel α-MSH peptide analogues with broad spectrum antimicrobial activity. PLoS ONE 2013, 8, e61614. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Grazia, A.D.; Cappiello, F.; Casciaro, B.; Luca, V. Naturally occurring peptides from Rana Temporaria: Antimicrobial properties and more. Curr. Top. Med. Chem. 2016, 16, 54–64. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.; Malfi, S.; Marcellini, L.; Barra, D.; Novellino, E.; et al. Structure−activity relationship, conformational and biological studies of Temporin L analogues. J. Med. Chem. 2011, 54, 1298–1307. [Google Scholar] [CrossRef]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.M.; Marcellini, L.; Luca, V.; Barra, D.; Novellino, E.; et al. The effect of d-amino acid substitution on the selectivity of temporin L towards target cells: Identification of a potent anti-Candida peptide. Biochim. Biophys. Acta 2013, 1828, 652–660. [Google Scholar] [CrossRef]
- Merlino, F.; Carotenuto, A.; Casciaro, B.; Martora, F.; Loffredo, M.R.; Di Grazia, A.; Yousif, A.M.; Brancaccio, D.; Palomba, L.; Novellino, E.; et al. Glycine-replaced derivatives of [Pro3,Dleu9]TL, a temporin L analogue: Evaluation of antimicrobial, cytotoxic and hemolytic activities. Eur. J. Med. Chem. 2017, 139, 750–761. [Google Scholar] [CrossRef]
- Buommino, E.; Carotenuto, A.; Antignano, I.; Bellavita, R.; Casciaro, B.; Loffredo, M.R.; Merlino, F.; Novellino, E.; Mangoni, M.L.; Nocera, F.P.; et al. The Outcomes of decorated prolines in the discovery of antimicrobial peptides from Temporin-L. Chem. Med. Chem. 2019, 14, 1283–1290. [Google Scholar] [CrossRef]
- Angiolella, L.; Carradori, S.; Maccallini, C.; Giusiano, G.; Supuran, C.T. Targeting Malassezia species for novel synthetic and natural antidandruff agents. Curr. Med. Chem. 2017, 24, 2392–2412. [Google Scholar] [CrossRef]
- Bellavita, R.; Raucci, F.; Merlino, F.; Piccolo, M.; Ferraro, M.G.; Irace, C.; Santamaria, R.; Iqbal, A.J.; Novellino, E.; Grieco, P.; et al. Temporin L-derived peptide as a regulator of the acute inflammatory response in zymosan-induced peritonitis. Biomed. Pharmacother. 2020, 123, 109788. [Google Scholar] [CrossRef]
- Gottschalk, S.; Thomsen, L.E. The Interaction of antimicrobial peptides with the membrane and intracellular targets of Staphylococcus aureus Investigated by ATP leakage, DNA-binding analysis, and the expression of a LexA-controlled gene, recA. Methods Mol. Biol. 2017, 1548, 297–305. [Google Scholar]
- Gupta, K.; Singh, S.; van Hoek, M.L. Short, synthetic cationic peptides have antibacterial activity against Mycobacterium smegmatis by forming pores in membrane and synergizing with antibiotics. Antibiotics 2015, 4, 358–378. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, L.; Falanga, A.; Del Genio, V.; Galdiero, S. A new hope: Self-assembling peptides with antimicrobial activity. Pharmaceutics 2019, 11, 166. [Google Scholar] [CrossRef] [Green Version]
- Zharkova, M.S.; Orlov, D.S.; Golubeva, O.Y.; Chakchir, O.B.; Eliseev, I.E.; Grinchuk, T.M.; Shamova, O.V. Application of antimicrobial peptides of the innate immune system in combination with conventional antibiotics-a novel way to combat antibiotic resistance? Front. Cell. Infect. Microbiol. 2019, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Brancaccio, D.; Pizzo, E.; Cafaro, V.; Notomista, E.; De Lise, F.; Bosso, A.; Gaglione, R.; Merlino, F.; Novellino, E.; Ungaro, F.; et al. Antimicrobial peptide Temporin-L complexed with anionic cyclodextrins results in a potent and safe agent against sessile bacteria. Int. J. Pharm. 2020, 584, 119437. [Google Scholar] [CrossRef]
- Merlino, F.; Tomassi, S.; Yousif, A.M.; Messere, A.; Marinelli, L.; Grieco, P.; Novellino, E.; Cosconati, S.; Di Maro, S. Boosting Fmoc solid-phase peptide synthesis by ultrasonication. Org. Lett. 2019, 21, 6378–6382. [Google Scholar] [CrossRef]
- Buommino, E.; Nocera, F.P.; Parisi, A.; Rizzo, A.; Donnarumma, G.; Mallardo, K.; Fiorito, F.; Baroni, A.; De Martino, L. Correlation between genetic variability and virulence factors in clinical strains of Malassezia pachydermatis of animal origin. New Microbiol. 2016, 39, 216–223. [Google Scholar]
- Papich, M.G.; Simjee, S.; Apley, M.; Frana, S.T.; Knapp, C.C.; Lubbers, B.V.; Rose, M.; Schwarz, S.; Silley, P.; Sweeney, M.T.; et al. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; NCCLS: Wayne, PA, USA, 2018. [Google Scholar]
- Cafarchia, C.; Figueredo, L.A.; Favuzzi, V.; Surico, M.R.; Colao, V.; Iatta, R.; Montagna, M.T.; Otranto, D. Assessment of the antifungal susceptibility of Malassezia pachydermatis in various media using a CLSI protocol. Vet. Microbiol. 2012, 159, 536–540. [Google Scholar] [CrossRef]
- Olajuyigbe, O.O.; Afolayan, A.J. In vitro antibacterial and time-kill evaluation of the Erythrina caffrathunb. Extract against bacteria associated with diarrhea. Sci. World J. 2012, 738314. [Google Scholar]
- Pillai, S.K.; Moellering, R.C.; Eliopoulo, G.M. Antimicrobial combinations. In Antibiotics in Laboratory Medicine, 5th ed; Lorian, V., Ed.; The Lippincott Williams & Wilkins Co.: Philadelphia, PA, USA, 2005; pp. 365–440. [Google Scholar]
- Barra, F.; Roscetto, E.; Soriano, A.A.; Vollaro, A.; Postiglione, I.; Pierantoni, G.M.; Palumbo, G.; Catania, M.R. Photodynamic and antibiotic therapy in combination to fight biofilms and resistant surface bacterial infections. Int. J. Mol. Sci. 2015, 16, 20417–20430. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar]
- Vollaro, A.; Catania, M.R.; Iesce, M.R.; Sferruzza, R.; D’Abrosca, B.; Donnarumma, G.; De Filippis, A.; Cermola, F.; DellaGreca, M.; Buommino, E. Antimicrobial and anti-biofilm properties of novel synthetic lignan-like compounds. New Microbiol. 2019, 42, 21–28. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cmpds | MIC Values (μM) | MMC Values (μM) | MMC/MIC Ratio |
|---|---|---|---|
| 1 | 25 | 100 | 4 |
| 2 | 25 | 100 | 4 |
| 3 | 25 | 100 | 4 |
| 4 | 25 | 100 | 4 |
| 5 | 25 | 100 | 4 |
| 6 | 25 | 100 | 4 |
| 7 | 12.5 | 25 | 2 |
| 8 | 6.25 | 6.25 | 1 |
| 9 | 25 | 100 | 4 |
| 10 | 12.5 | 25 | 2 |
| 11 | 25 | 100 | 4 |
| Strains | Log10 CFU/Ml After 1 Generation Subculture Including TL48 | Log10 CFU/Ml After 15 Generation Subcultures Including TL48 |
|---|---|---|
| M. pachydermatis Untreated | 8.10 ± 0.21 | 8.26 ± 0.29 |
| M. pachydermatis + Cmpd 8 | 5.80 ± 0.25 | 2.10 ± 0.12 |
| Strains | MIC | MBC | Oxacillin | Vancomycin | MBC/MIC Ratio | FICindex |
|---|---|---|---|---|---|---|
| MSSP | 1.56 | 3.12 | <5 | <1.4 | 2 | n.d. |
| MRSP | 6.25 | 12.5 | 25 | <1.4 | 2 | 0.37 |
| Strains | Log10CFU/mL | |||||
|---|---|---|---|---|---|---|
| 0 h | 1 h | 2 h | 4 h | 6 h | 24 h | |
| MSSP untreated | 5.50 ± 0.21 | 5.74 ± 0.21 | 5.80 ± 0.25 | 5.87 ± 0.19 | 7.10 ± 0.31 | 9.8 ± 0.31 |
| MSSP+TL48 | 5.45 ± 0.25 | 0.60 ± 0.12 | 0.58 ± 0.32 | 0.89 ± 0.15 | 0.46 ± 0.15 | 4.97 ± 0.30 |
| MRSP untreated | 5.30 ± 0.21 | 5.37 ± 0.32 | 5.84 ± 0.28 | 5.91 ± 0.29 | 6.85 ± 0.25 | 9.36 ± 0.25 |
| MRS+TL48 | 5.30 ± 0.18 | 2.47 ± 0.18 | 2.38 ± 0.19 | 1.30 ± 0.21 | 0.77 ± 0.12 | 4.86 ± 0.28 |
| Cmpd | Sequence |
|---|---|
| 1 | F V P W F S K F l Gly10 R I L |
| 2 | F V P W F S K F l Pro10 R I L |
| 3 | F V P W F S K F l dPro10 R I L |
| 4 | F V P W F S K F l Hyp10R I L |
| 5 | F V P W F S K F l dHyp10 R I L |
| 6 | F V P W F S K F l dNle10R I L |
| 7 | F V P W F S K F l Lys10R I L |
| 8 | F V P W F S K F l dLys10R I L |
| 9 | F V P W F S K F l Trp10R I L |
| 10 | F V P W F S K F l dTrp10R I L |
| 11 | F V P W F S K F l Aic10R I L |
| Gene | Sense and Antisense Sequences | Conditions | Bp |
|---|---|---|---|
| mecA | 5′-TCCACCCTCAAACAGGTGAA-3′ 5′-TGGAACTTGTTGAGCAGAGGT-3′ | 95 °C for 5′ 94 °C 30″, 55°C 4″s, 72 °C 30″ for 33 cycles 72 °C for 7′ | 139 |
| mecI | 5′-TCATCTGCAGAATGGGAAGTT-3′ 5′-TTGGACTCCAGTCCTTTTGC-3′ | 103 | |
| mecR1 | 5′-AGCACCGTTACTATCTGCACA-3′ 5′-AGAATAAGCTTGCTCCCGTTCA-3′ | 142 | |
| rRNA16S | 5′-CGGTCCAGACTCCTACGGGAGGCAGCA-3′ 5′-GCGTGGACTACCAGGGTATCTAATCC-3′ | 450 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellavita, R.; Vollaro, A.; Catania, M.R.; Merlino, F.; De Martino, L.; Nocera, F.P.; DellaGreca, M.; Lembo, F.; Grieco, P.; Buommino, E. Novel Antimicrobial Peptide from Temporin L in The Treatment of Staphylococcus pseudintermedius and Malassezia pachydermatis in Polymicrobial Inter-Kingdom Infection. Antibiotics 2020, 9, 530. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090530
Bellavita R, Vollaro A, Catania MR, Merlino F, De Martino L, Nocera FP, DellaGreca M, Lembo F, Grieco P, Buommino E. Novel Antimicrobial Peptide from Temporin L in The Treatment of Staphylococcus pseudintermedius and Malassezia pachydermatis in Polymicrobial Inter-Kingdom Infection. Antibiotics. 2020; 9(9):530. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090530
Chicago/Turabian StyleBellavita, Rosa, Adriana Vollaro, Maria Rosaria Catania, Francesco Merlino, Luisa De Martino, Francesca Paola Nocera, Marina DellaGreca, Francesca Lembo, Paolo Grieco, and Elisabetta Buommino. 2020. "Novel Antimicrobial Peptide from Temporin L in The Treatment of Staphylococcus pseudintermedius and Malassezia pachydermatis in Polymicrobial Inter-Kingdom Infection" Antibiotics 9, no. 9: 530. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9090530