In Vitro Activity of Dalbavancin against Refractory Multidrug-Resistant (MDR) Staphylococcus aureus Isolates



Abstract
:1. Introduction
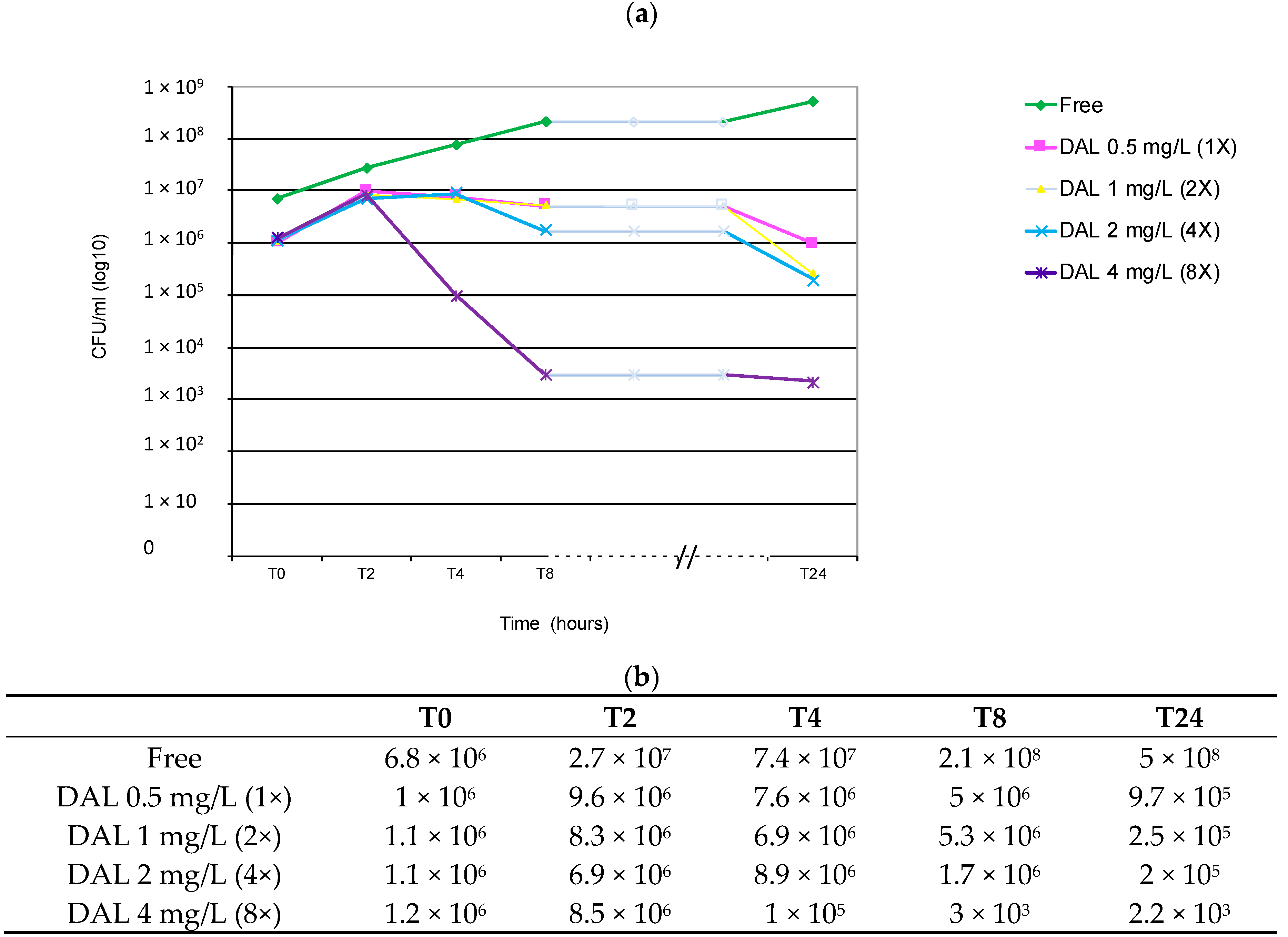
2. Results and Discussion
3. Methods Section
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hsueh, P.R.; Lee, S.Y.; Perng, C.L.; Chang, T.Y.; Lu, J.J. Clonal dissemination of meticillin-resistant and vancomycin-intermediate Staphylococcus aureus in a Taiwanese hospital. Int. J. Antimicrob. Agents 2010, 36, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Howden, B.P.; Peleg, A.Y.; Stinear, T.P. The evolution of vancomycin intermediate Staphylococcus aureus (VISA) and heterogenous-VISA. Infect. Genet. Evol. 2014, 21, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Thitiananpakorn, K.; Aiba, Y.; Tan, X.E.; Watanabe, S.; Kiga, K.; Sato’o, Y.; Boonsiri, T.; Li, F.Y.; Sasahara, T.; Taki, Y.; et al. Association of mprF mutations with cross-resistance to daptomycin and vancomycin in methicillin-resistant Staphylococcus aureus (MRSA). Sci. Rep. 2020, 10, 16107. [Google Scholar] [CrossRef] [PubMed]
- Klinker, K.P.; Borgert, S.J. Beyond Vancomycin: The Tail of the Lipoglycopeptides. Clin. Ther. 2015, 37, 2619–2636. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Mutakabbir, J.C.; Kebriaei, R.; Stamper, K.C.; Sheikh, Z.; Maassen, P.T.; Lev, K.L.; Rybak, M.J. Dalbavancin, Vancomycin and Daptomycin Alone and in Combination with Cefazolin against Resistant Phenotypes of Staphylococcus aureus in a Pharmacokinetic/Pharmacodynamic Model. Antibiotics 2020, 9, 696. [Google Scholar] [CrossRef] [PubMed]
- Xhemali, X.; Smith, J.R.; Kebriaei, R.; Rice, S.R.; Stamper, K.C.; Compton, M.; Singh, N.B.; Jahanbakhsh, S.; Rybak, M.J. Evaluation of dalbavancin alone and in combination with β-lactam antibiotics against resistant phenotypes of Staphylococcus aureus. J. Antimicrob. Chemother. 2019, 74, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Campanile, F.; Bongiorno, D.; Perez, M.; Mongelli, G.; Sessa, L.; Benvenuto, S.; Gona, F. AMCLI—S. aureus Survey Participants, Varaldo, P.E., Stefani, S. Epidemiology of Staphylococcus aureus in Italy: First nationwide survey, 2012. J. Glob. Antimicrob. Resist. 2015, 3, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno, D.; Mongelli, G.; Stefani, S.; Campanile, F. Burden of Rifampicin- and Methicillin-Resistant Staphylococcus aureus in Italy. Microb. Drug Resist. 2018, 24, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Cui, L.; Katayama, Y.; Kozue, K.; Hiramatsu, K. Impact of rpoB mutations on reduced vancomycin susceptibility in Staphylococcus aureus. J. Clin. Microbiol. 2011, 49, 2680–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, M.; Sekizuka, T.; Matsui, H.; Ohsuga, J.; Ohshima, T.; Hanaki, H. IS256-Mediated Overexpression of the WalKR Two-Component System Regulon Contributes to Reduced Vancomycin Susceptibility in a Staphylococcus aureus Clinical Isolate. Front. Microbiol. 2019, 10, 1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedenbach, D.J.; Bell, J.M.; Sader, H.S.; Fritsche, T.R.; Jones, R.N.; Turnidge, J.D. Antimicrobial susceptibility of Gram-positive bacterial isolates from the Asia-Pacific region and an in vitro evaluation of the bactericidal activity of daptomycin, vancomycin, and teicoplanin: A SENTRY Program Report (2003–2004). Int. J. Antimicrob. Agents 2007, 30, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Capone, A.; Cafiso, V.; Campanile, F.; Parisi, G.; Mariani, B.; Petrosillo, N.; Stefani, S. In vivo development of daptomycin resistance in vancomycin-susceptible methicillin-resistant Staphylococcus aureus severe infections previously treated with glycopeptides. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 625–631. [Google Scholar] [CrossRef] [PubMed]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 10.0; 2020. Available online: http://www.eucast.org (accessed on 1 January 2020).
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; M100; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Campanile, F.; Borbone, S.; Perez, M.; Bongiorno, D.; Cafiso, V.; Bertuccio, T.; Purrello, S.; Nicolosi, D.; Scuderi, C.; Stefani, S. Heteroresistance to glycopeptides in Italian meticillin-resistant Staphylococcus aureus (MRSA) isolates. Int. J. Antimicrob. Agents 2010, 36, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI) 2015 (M07-A10). Methods for Dilution Antimicrobial Susceptibility Tests Forbacteria that Grow Aerobically; Approved Standard, 10th ed.; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- National Committee for Clinical Laboratory Standards (NCCLS) 1999 (M26-A). Methods for Determining Bactericidal Activity of Antimicrobial Agents; Approved Guidelines; NCCLS: Wayne, PA, USA, 1999. [Google Scholar]
- Sader, H.S.; Streit, J.M.; Mendes, R.E. Update on the in vitro activity of dalbavancin against indicated species (Staphylococcus aureus, Enterococcus faecalis, β-hemolytic streptococci, and Streptococcus anginosus group) collected from United States hospitals in 2017–2019. Diagn. Microbiol. Infect. Dis. 2020, 99, 115195. [Google Scholar] [CrossRef] [PubMed]
- Van Hal, S.J.; Paterson, D.L. Systematic review and meta-analysis of the significance of heterogeneous vancomycin-intermediate Staphylococcus aureus isolates. Antimicrob. Agents Chemother. 2011, 55, 405–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sader, H.S.; Mendes, R.E.; Duncan, L.R.; Pfaller, M.A.; Flamm, R.K. Antimicrobial Activity of Dalbavancin against Staphylococcus aureus with Decreased Susceptibility to Glycopeptides, Daptomycin, and/or Linezolid from U.S. Medical Centers. Antimicrob. Agents Chemother. 2018, 62, e02397-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramatsu, K.; Kayayama, Y.; Matsuo, M.; Aiba, Y.; Saito, M.; Hishinuma, T.; Iwamoto, A. Vancomycin-intermediate resistance in Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2014, 2, 213–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrothers, T.J.; Chittenden, J.T.; Critchley, I. Dalbavancin Population Pharmacokinetic Modeling and Target Attainment Analysis. Clin. Pharmacol. Drug Dev. 2020, 9, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strains | n. of Strains | MIC Range (mg/L) | MIC50 (mg/L) | MIC90 (mg/L) | n. −% (R) |
|---|---|---|---|---|---|
| MSSA | 23 | ≤0.007–0.125 | 0.03 | 0.125 | 0 |
| MRSA/VSSA (HA/CA-MRSA) | 25 | 0.015–0.25 | 0.06 | 0.125 | 1 (4%) |
| MRSA/hVISA (HA/CA-MRSA) | 22 | ≤0.007–0.125 | 0.06 | 0.125 | 0 |
| MRSA/DNS (hVISA + VISA) | 4 (3hVISA + 1VISA) | 0.06–2 | 0.06 | 0.125 | 1 (25%) |
| MRSA/RIF-R (hVISA + VSSA) | 50 (31hVISA + 19VSSA) | 0.015–0.5 | 0.125 | 0.25 | 9 (18%) |
| Tot S. aureus | 124 | ≤0.007–2 | 0.06 | 0.125 | 11 (8.8%) |
| Subcategory | Strain | ST-MRSA-SCCmec | spa-Type | Dalbavancin MIC (mg/L) |
|---|---|---|---|---|
| MSSA | ATCC29213 | - | - | 0.06 |
| MRSA-hVISA | Mu3 | ST5-hVISA-II | t002 | 0.125 |
| MRSA-VISA | Mu50 | ST5-hVISA-II | t002 | 1 |
| MRSA-hVISA | NRS22 | ST45-hVISA-II | t266 | 0.03 |
| MRSA-VISA (DNS) | NRS402 | ST5-VISA-II | t002 | 1 |
| MRSA-VRSA | VRS1 | ST5-VRSA Tn1546 vanA | - | ≥4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bongiorno, D.; Lazzaro, L.M.; Stefani, S.; Campanile, F. In Vitro Activity of Dalbavancin against Refractory Multidrug-Resistant (MDR) Staphylococcus aureus Isolates. Antibiotics 2020, 9, 865. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120865
Bongiorno D, Lazzaro LM, Stefani S, Campanile F. In Vitro Activity of Dalbavancin against Refractory Multidrug-Resistant (MDR) Staphylococcus aureus Isolates. Antibiotics. 2020; 9(12):865. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120865
Chicago/Turabian StyleBongiorno, Dafne, Lorenzo Mattia Lazzaro, Stefania Stefani, and Floriana Campanile. 2020. "In Vitro Activity of Dalbavancin against Refractory Multidrug-Resistant (MDR) Staphylococcus aureus Isolates" Antibiotics 9, no. 12: 865. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9120865