
Changes in Hepatic Phospholipid Metabolism in Rats under UV Irradiation and Topically Treated with Cannabidiol



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Experiment
- Control group: petrolatum applied topically (20 min) every 12 h for 4 weeks;
- CBD group: CBD (120 mg/kg of body weight (b.w.); 2.5% w/w in petrolatum) applied topically (20 min) every 12 h for 4 weeks;
- UVA group: UVA radiation (365 nm) every 48 h (dose increasing from 0.5 to 5 J/cm2) for 4 weeks;
- UVA+CBD group: UVA radiation (365 nm) every 48 h (dose increasing from 0.5 to 5 J/cm2) and CBD (120 mg/kg of b.w.; 2.5% w/w in petrolatum) applied topically (20 min) every 12 h for 4 weeks;
- UVB group: UVB radiation (312 nm) every 48 h (dose increasing from 0.02 to 2 J/cm2) for 4 weeks;
- UVB+CBD group: UVB radiation (312 nm) every 48 h (dose increasing from 0.02 to 2 J/cm2) and CBD (120 mg/kg of b.w.; 2.5% w/w in petrolatum) applied topically (20 min) every 12 h for 4 weeks.
2.3. Tissue Preparation
2.4. Biochemical Studies
2.4.1. Determination of CBD and Its Metabolites Level
2.4.2. Determination of Antioxidants Activity/Level
2.4.3. Determination of Phospholipid Metabolism and Mediators
Phospholipid Peroxidation Products
Enzymes Metabolizing Phospholipids
Eicosanoids
Endocannabinoids
2.5. Statistical Analysis
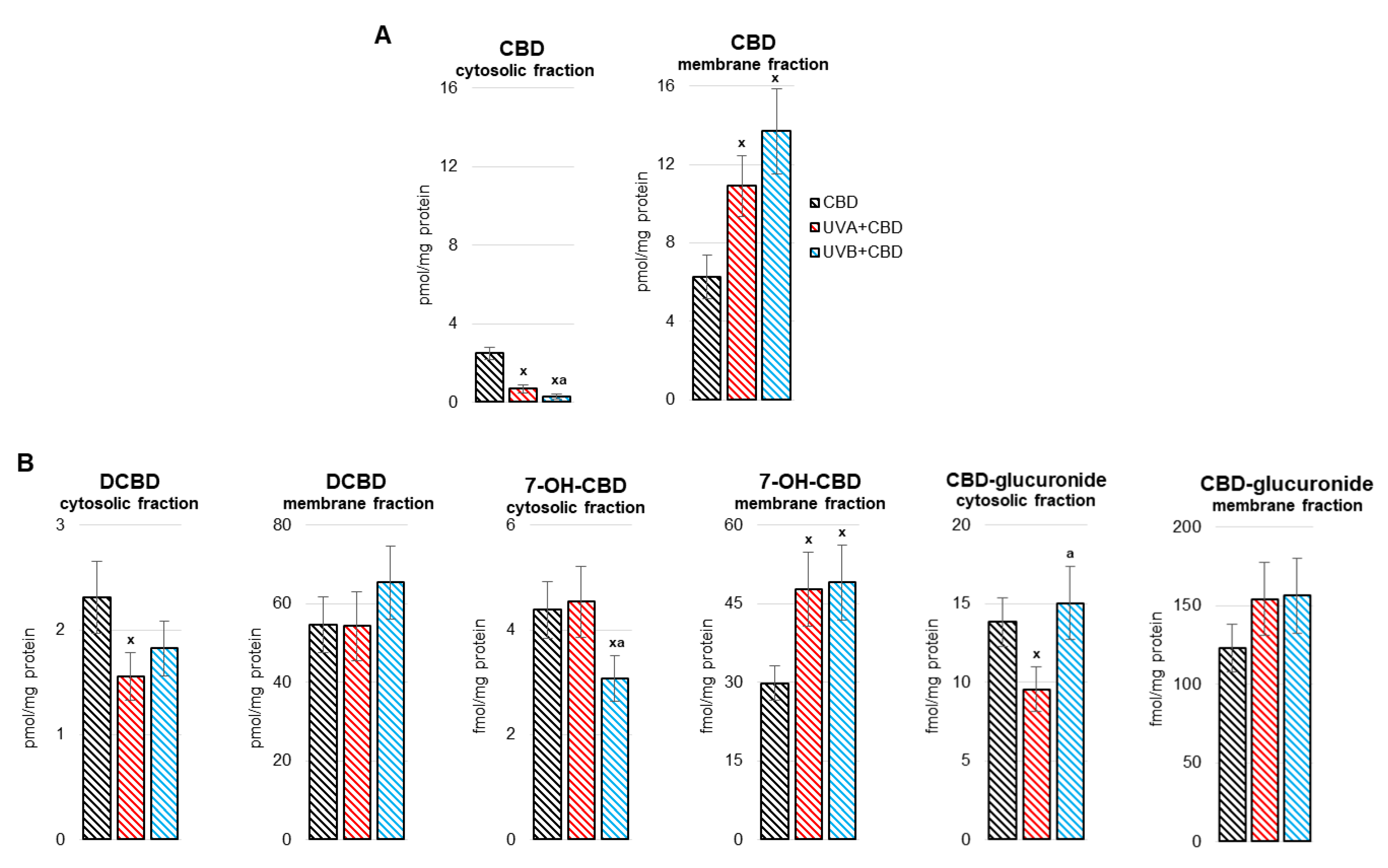
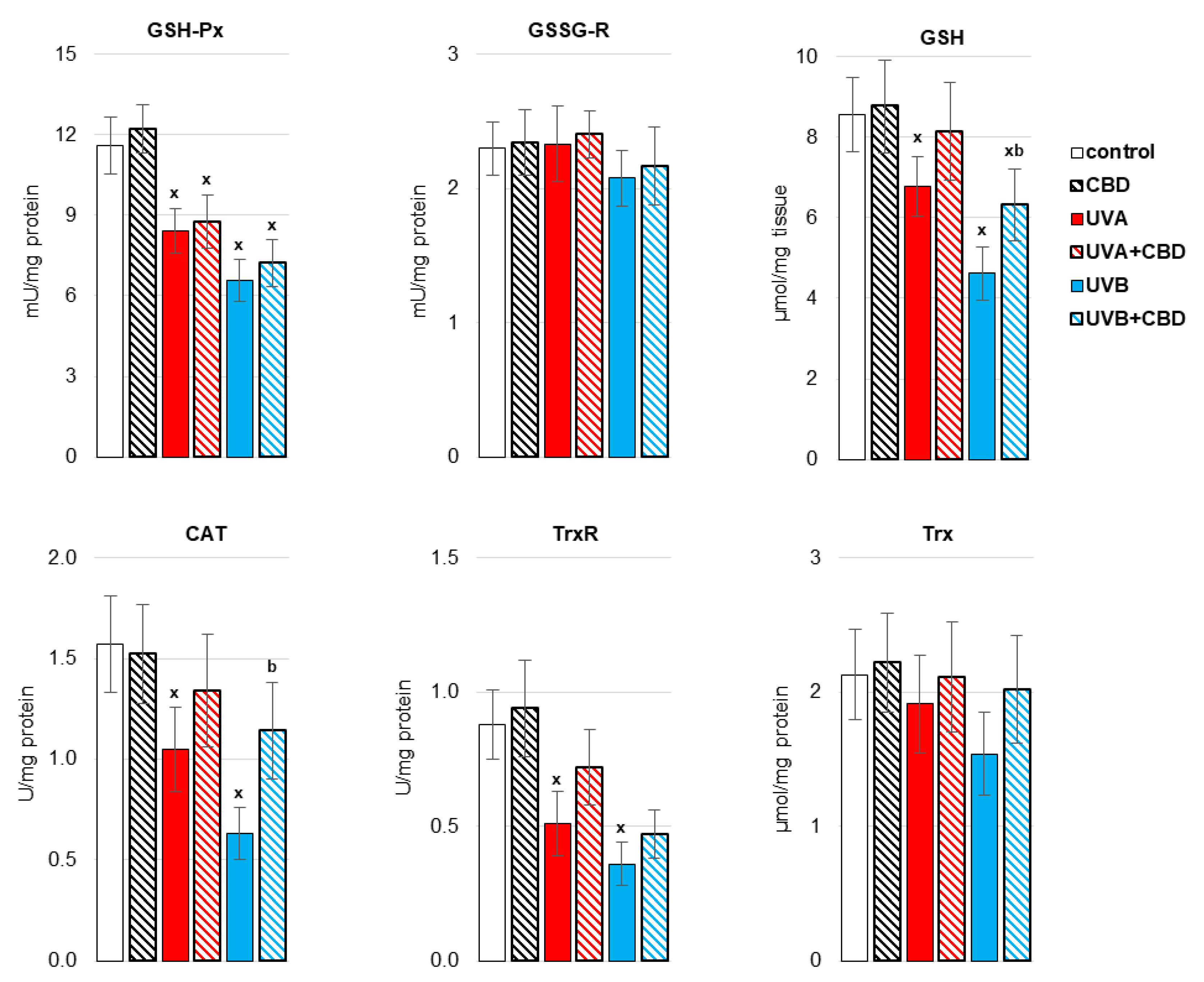
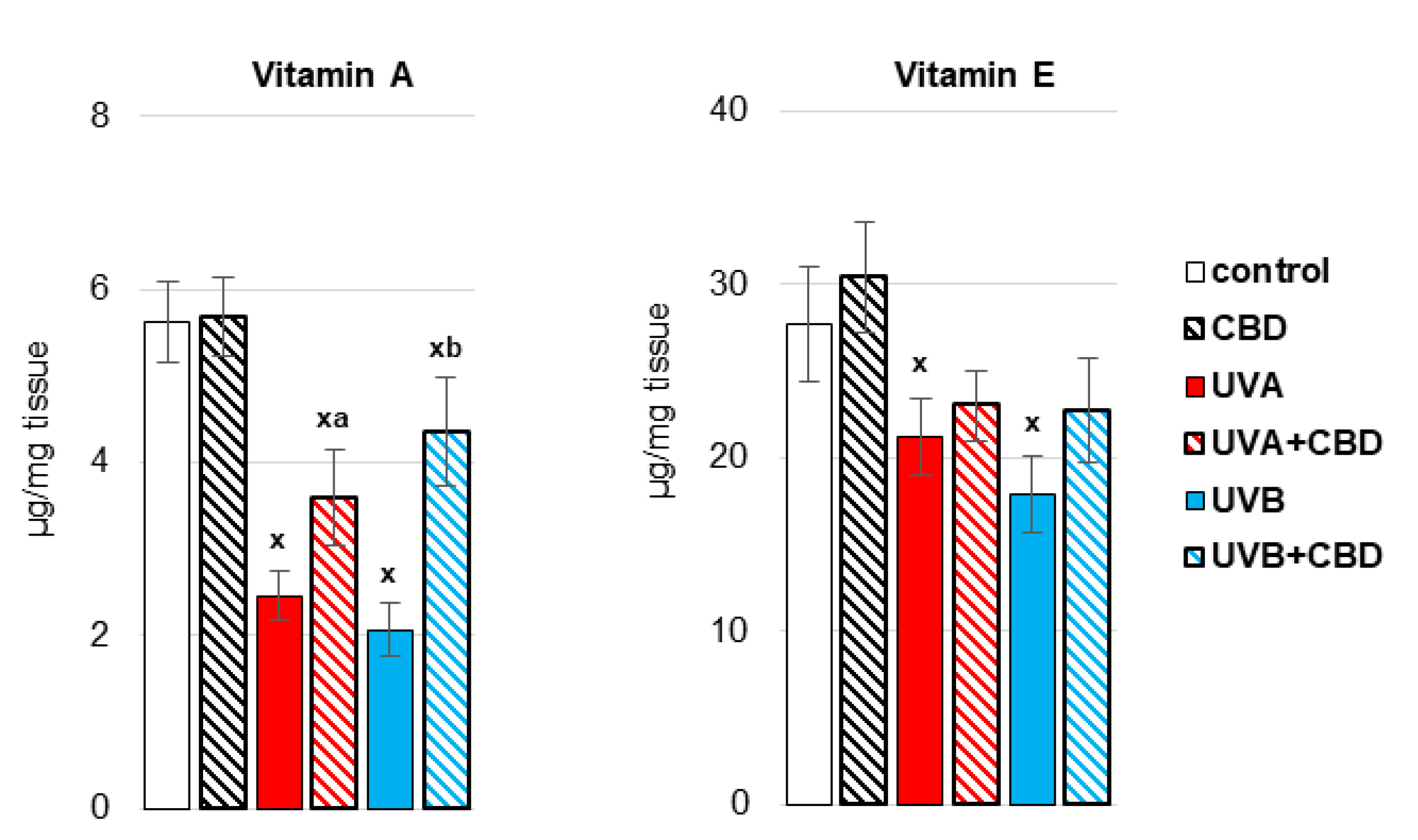
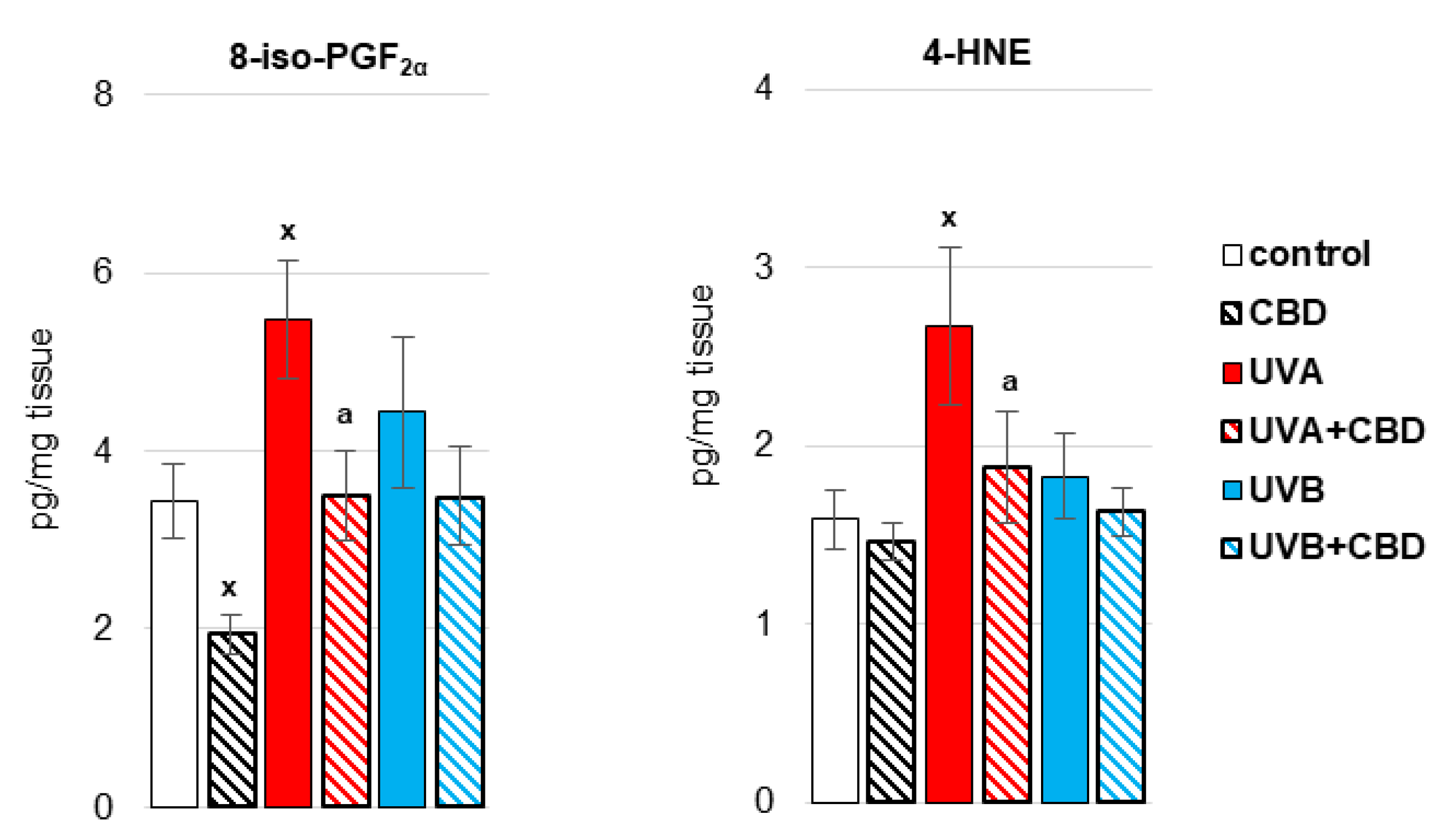
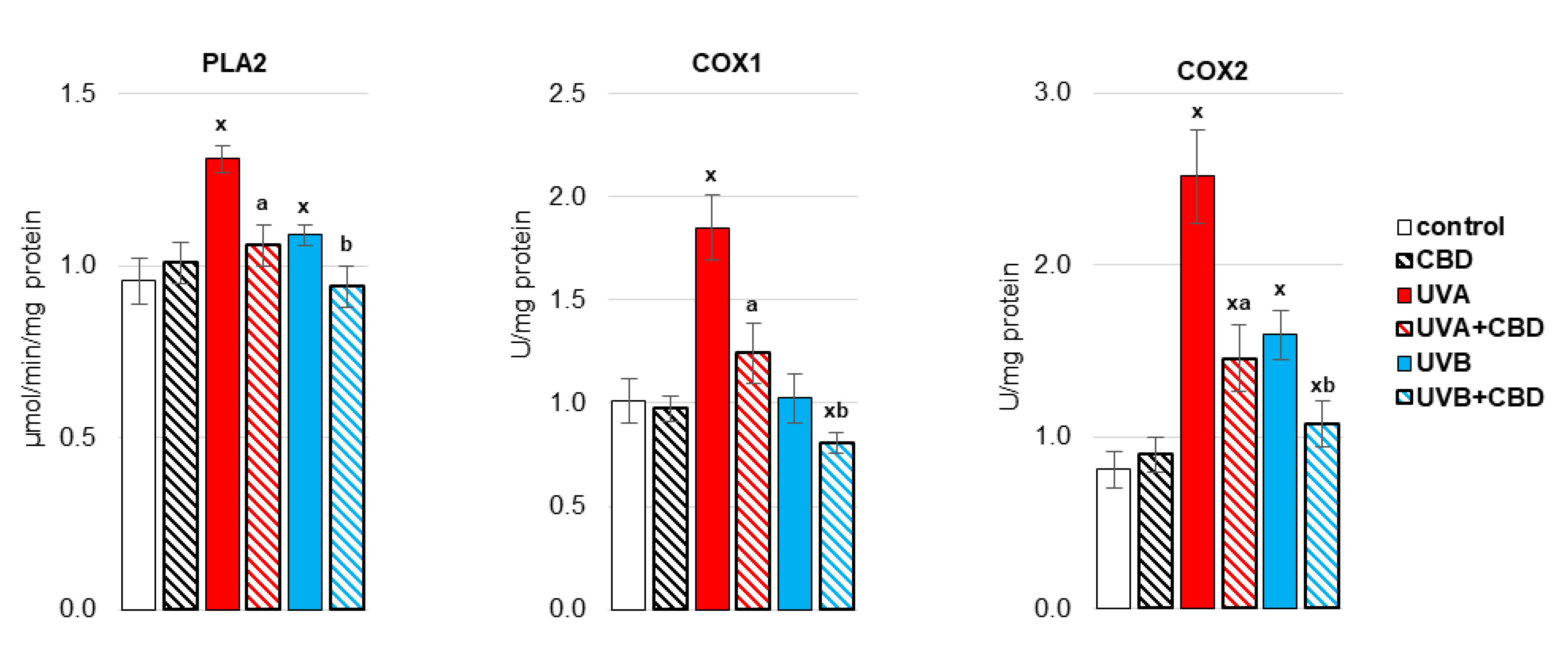
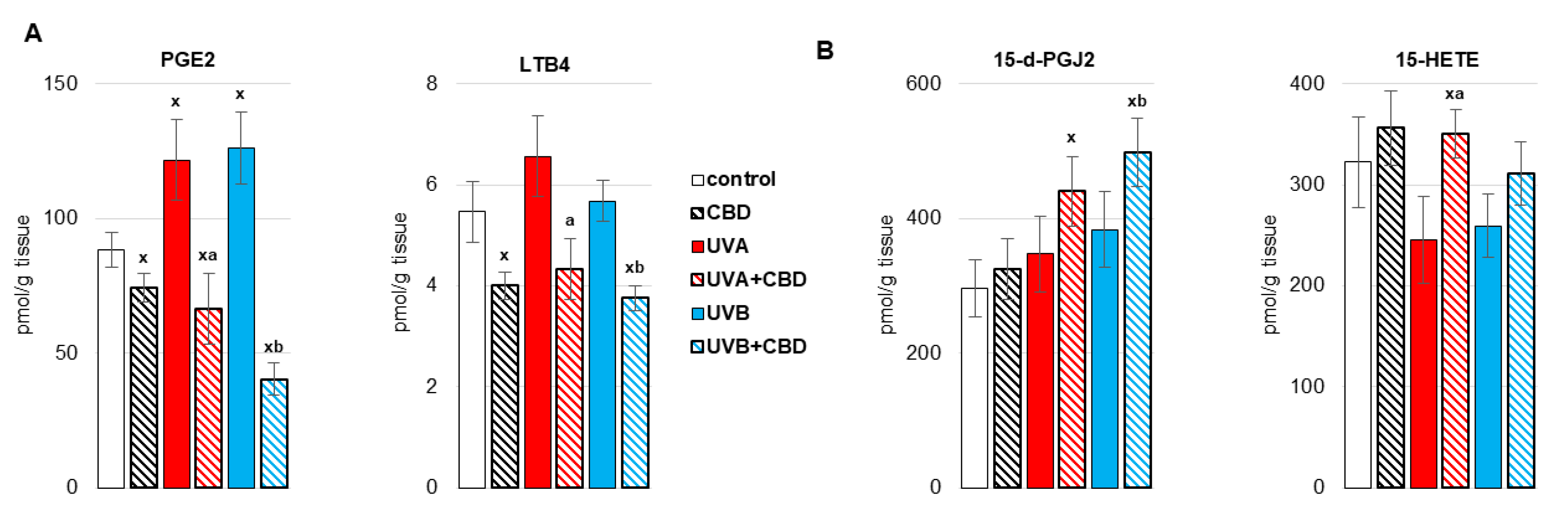
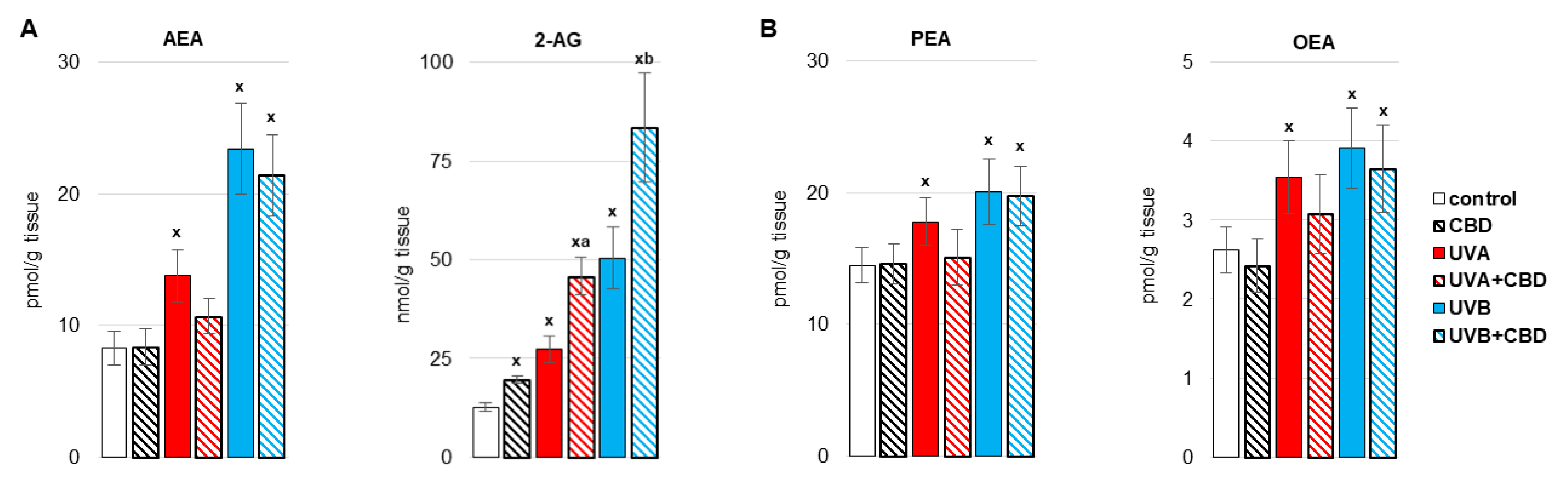
3. Results
4. Discussion
4.1. Effect of UV Radiation on the Metabolism of Liver Phospholipids
4.2. Effect of CBD on the Metabolism of Liver Phospholipids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, X.-F.; Chen, D.-D.; Zhu, H.-J.; Ge, W.-H. Prognostic Value of Endogenous and Exogenous Metabolites in Liver Transplantation. Biomark Med. 2020, 14, 1165–1181. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’H, J.; Siliart, B.; Dumon, H. Liver Lipid Metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Tontonoz, P. Phospholipid Remodeling in Physiology and Disease. Annu. Rev. Physiol. 2019, 81, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Chamulitrat, W.; Jansakun, C.; Li, H.; Liebisch, G. Rescue of Hepatic Phospholipid Remodeling Defect in IPLA2β-Null Mice Attenuates Obese but Not Non-Obese Fatty Liver. Biomolecules 2020, 10, 1332. [Google Scholar] [CrossRef] [PubMed]
- Galano, J.-M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C.-Y. Isoprostanes, Neuroprostanes and Phytoprostanes: An Overview of 25 years of Research in Chemistry and Biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef]
- Viedma-Poyatos, Á.; González-Jiménez, P.; Langlois, O.; Company-Marín, I.; Spickett, C.M.; Pérez-Sala, D. Protein Lipoxidation: Basic Concepts and Emerging Roles. Antioxidants 2021, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Spickett, C.M. Chemistry of Phospholipid Oxidation. Biochim. Biophys. Acta. Biomembr. 2012, 1818, 2374–2387. [Google Scholar] [CrossRef] [Green Version]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and Related N-Acylethanolamines: Biological Activities and Metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef]
- Alger, B.E.; Kim, J. Supply and Demand for Endocannabinoids. Trends Neurosci. 2011, 34, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Bazwinsky-Wutschke, I.; Zipprich, A.; Dehghani, F. Endocannabinoid System in Hepatic Glucose Metabolism, Fatty Liver Disease, and Cirrhosis. Int. J. Mol. Sci. 2019, 20, 2516. [Google Scholar] [CrossRef] [Green Version]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of Arachidonic Acid Metabolism: A Review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef]
- Gladine, C.; Ostermann, A.I.; Newman, J.W.; Schebb, N.H. MS-Based Targeted Metabolomics of Eicosanoids and Other Oxylipins: Analytical and Inter-Individual Variabilities. Free. Radic. Biol. Med. 2019, 144, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, D.; Drozd, A.; Skonieczna-Żydecka, K.; Skórka-Majewicz, M.; Dec, K.; Jakubczyk, K.; Pilutin, A.; Stachowska, E. Eicosanoids in Nonalcoholic Fatty Liver Disease (NAFLD) Progression. Do Serum Eicosanoids Profile Correspond with Liver Eicosanoids Content during NAFLD Development and Progression? Molecules 2020, 25, 2026. [Google Scholar] [CrossRef] [PubMed]
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, A.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism Disrupting Chemicals and Metabolic Disorders. Reprod. Toxicol. 2017, 68, 3–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erden Inal, M.; Kahraman, A. The Protective Effect of Flavonol Quercetin against Ultraviolet a Induced Oxidative Stress in Rats. Toxicology 2000, 154, 21–29. [Google Scholar] [CrossRef]
- Svobodová, A.R.; Galandáková, A.; Sianská, J.; Doležal, D.; Ulrichová, J.; Vostálová, J. Acute Exposure to Solar Simulated Ultraviolet Radiation Affects Oxidative Stress-Related Biomarkers in Skin, Liver and Blood of Hairless Mice. Biol. Pharm. Bull. 2011, 34, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, N.; Reeve, V.E.; Nishimura, H.; Satoh, M.; Tohyama, C. Cutaneous Metallothionein Induction by Ultraviolet B Irradiation in Interleukin-6 Null Mice. J. Investig. Dermatol. 2000, 114, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Vangipuram, R.; Feldman, S.R. Ultraviolet Phototherapy for Cutaneous Diseases: A Concise Review. Oral Dis. 2016, 22, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and Anti-Inflammatory Properties of Cannabidiol. Antioxidants 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Biernacki, M.; Brzóska, M.M.; Markowska, A.; Gałażyn-Sidorczuk, M.; Cylwik, B.; Gęgotek, A.; Skrzydlewska, E. Oxidative Stress and Its Consequences in the Blood of Rats Irradiated with UV: Protective Effect of Cannabidiol. Antioxidants 2021, 10, 821. [Google Scholar] [CrossRef]
- Kicman, A.; Toczek, M. The Effects of Cannabidiol, a Non-Intoxicating Compound of Cannabis, on the Cardiovascular System in Health and Disease. Int. J. Mol. Sci. 2020, 21, 6740. [Google Scholar] [CrossRef] [PubMed]
- Ujváry, I.; Hanuš, L. Human Metabolites of Cannabidiol: A Review on Their Formation, Biological Activity, and Relevance in Therapy. Cannabis Cannabinoid Res. 2016, 1, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Perucca, E.; Bialer, M. Critical Aspects Affecting Cannabidiol Oral Bioavailability and Metabolic Elimination, and Related Clinical Implications. CNS Drugs 2020, 34, 795–800. [Google Scholar] [CrossRef]
- Wang, M.-H.; Hsiao, G.; Al-Shabrawey, M. Eicosanoids and Oxidative Stress in Diabetic Retinopathy. Antioxidants 2020, 9, 520. [Google Scholar] [CrossRef] [PubMed]
- Gallelli, C.A.; Calcagnini, S.; Romano, A.; Koczwara, J.B.; de Ceglia, M.; Dante, D.; Villani, R.; Giudetti, A.M.; Cassano, T.; Gaetani, S. Modulation of the Oxidative Stress and Lipid Peroxidation by Endocannabinoids and Their Lipid Analogues. Antioxidants 2018, 7, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katica, M.; Gradascevic, N. Hematologic Profile Of Laboratory Rats Fed With Bakery Products. Int. J. Res. Granthaalayah 2017, 5, 221–231. [Google Scholar] [CrossRef]
- Luque-Córdoba, D.; Calderón-Santiago, M.; Luque de Castro, M.D.; Priego-Capote, F. Study of Sample Preparation for Determination of Endocannabinoids and Analogous Compounds in Human Serum by LC-MS/MS in MRM Mode. Talanta 2018, 185, 602–610. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Mize, C.E.; Langdon, R.G. Hepatic Glutathione Reductase. I. Purification and General Kinetic Properties. J. Biol. Chem. 1962, 237, 1589–1595. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Holmgren, A.; Björnstedt, M. Thioredoxin and Thioredoxin Reductase. Methods Enzymol. 1995, 252, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A.; Xie, C.; Gabbita, S.P.; Markesbery, W.R. Decreased Thioredoxin and Increased Thioredoxin Reductase Levels in Alzheimer’s Disease Brain. Free. Radic. Biol. Med. 2000, 28, 418–427. [Google Scholar] [CrossRef]
- De Leenheer, A.P.; De Bevere, V.O.; De Ruyter, M.G.; Claeys, A.E. Simultaneous Determination of Retinol and Alpha-Tocopherol in Human Serum by High-Performance Liquid Chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1979, 162, 408–413. [Google Scholar] [CrossRef]
- Maeso, N.; García-Martínez, D.; Rupérez, F.J.; Cifuentes, A.; Barbas, C. Capillary Electrophoresis of Glutathione to Monitor Oxidative Stress and Response to Antioxidant Treatments in an Animal Model. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2005, 822, 61–69. [Google Scholar] [CrossRef]
- Coolen, S.A.J.; van Buuren, B.; Duchateau, G.; Upritchard, J.; Verhagen, H. Kinetics of Biomarkers: Biological and Technical Validity of Isoprostanes in Plasma. Amino Acids 2005, 29, 429–436. [Google Scholar] [CrossRef]
- Luo, X.P.; Yazdanpanah, M.; Bhooi, N.; Lehotay, D.C. Determination of Aldehydes and Other Lipid Peroxidation Products in Biological Samples by Gas Chromatography-Mass Spectrometry. Anal. Biochem. 1995, 228, 294–298. [Google Scholar] [CrossRef]
- Reynolds, L.J.; Hughes, L.L.; Yu, L.; Dennis, E.A. 1-Hexadecyl-2-Arachidonoylthio-2-Deoxy-Sn-Glycero-3-Phosphorylcholine as a Substrate for the Microtiterplate Assay of Human Cytosolic Phospholipase A2. Anal. Biochem. 1994, 217, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Kulmacz, R.J.; Wang, L.H. Comparison of Hydroperoxide Initiator Requirements for the Cyclooxygenase Activities of Prostaglandin H Synthase-1 and -2. J. Biol. Chem. 1995, 270, 24019–24023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, B.A.; Kim, J.; Kenny, A.; Pedersen, T.L.; Pappan, K.L.; Newman, J.W. Circulating Levels of Endocannabinoids and Oxylipins Altered by Dietary Lipids in Older Women Are Likely Associated with Previously Identified Gene Targets. Biochim. Biophys. Acta 2016, 1861, 1693–1704. [Google Scholar] [CrossRef] [Green Version]
- Kremslehner, C.; Miller, A.; Nica, R.; Nagelreiter, I.-M.; Narzt, M.-S.; Golabi, B.; Vorstandlechner, V.; Mildner, M.; Lachner, J.; Tschachler, E.; et al. Imaging of Metabolic Activity Adaptations to UV Stress, Drugs and Differentiation at Cellular Resolution in Skin and Skin Equivalents—Implications for Oxidative UV Damage. Redox Biol. 2020, 37, 101583. [Google Scholar] [CrossRef]
- Jarocka-Karpowicz, I.; Biernacki, M.; Wroński, A.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Effects on Phospholipid Metabolism in Keratinocytes from Patients with Psoriasis Vulgaris. Biomolecules 2020, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Marzo, N.; Pérez-Sánchez, A.; Barrajón-Catalán, E.; Castillo, J.; Herranz-López, M.; Micol, V. Rosemary Diterpenes and Flavanone Aglycones Provide Improved Genoprotection against UV-Induced DNA Damage in a Human Skin Cell Model. Antioxidants 2020, 9, 255. [Google Scholar] [CrossRef] [Green Version]
- Atalay, S.; Dobrzyńska, I.; Gęgotek, A.; Skrzydlewska, E. Cannabidiol Protects Keratinocyte Cell Membranes Following Exposure to UVB and Hydrogen Peroxide. Redox Biol. 2020, 36, 101613. [Google Scholar] [CrossRef]
- Hasegawa, T.; Kaneko, F.; Niwa, Y. Changes in Lipid Peroxide Levels and Activity of Reactive Oxygen Scavenging Enzymes in Skin, Serum and Liver Following UVB Irradiation in Mice. Life Sci. 1992, 50, 1893–1903. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Schaur, R.J.; Siems, W.; Bresgen, N.; Eckl, P.M. 4-Hydroxy-Nonenal-A Bioactive Lipid Peroxidation Product. Biomolecules 2015, 5, 2247–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in Redox Homeostasis. Free. Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Karabowicz, P.; Wroński, A.; Ostrowska, H.; Waeg, G.; Zarkovic, N.; Skrzydlewska, E. Reduced Proteasome Activity and Enhanced Autophagy in Blood Cells of Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 7608. [Google Scholar] [CrossRef] [PubMed]
- Gresham, A.; Masferrer, J.; Chen, X.; Leal-Khouri, S.; Pentland, A.P. Increased Synthesis of High-Molecular-Weight CPLA2 Mediates Early UV-Induced PGE2 in Human Skin. Am. J. Physiol. 1996, 270, C1037–C1050. [Google Scholar] [CrossRef]
- Nicolaou, A.; Masoodi, M.; Gledhill, K.; Haylett, A.K.; Thody, A.J.; Tobin, D.J.; Rhodes, L.E. The Eicosanoid Response to High Dose UVR Exposure of Individuals Prone and Resistant to Sunburn. Photochem. Photobiol. Sci. 2012, 11, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive Lipids, Inflammation and Chronic Diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef]
- Law, S.-H.; Chan, M.-L.; Marathe, G.K.; Parveen, F.; Chen, C.-H.; Ke, L.-Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [Green Version]
- Speed, N.; Blair, I.A. Cyclooxygenase- and Lipoxygenase-Mediated DNA Damage. Cancer Metastasis Rev. 2011, 30, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Nagamachi, M.; Honda, T.; Nishigori, C.; Miyachi, Y.; Tokura, Y.; Narumiya, S. Prostaglandin E2 Is Required for Ultraviolet B-Induced Skin Inflammation via EP2 and EP4 Receptors. Lab. Investig. 2007, 87, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ji, X.; Wang, M.; Liao, X.; Liang, C.; Tang, J.; Wen, Z.; Dominique, F.; Li, Z. Involvement of TLR4 Signaling Regulated-COX2/PGE2 Axis in Liver Fibrosis Induced by Schistosoma Japonicum Infection. Parasit Vectors 2021, 14, 279. [Google Scholar] [CrossRef]
- Silvestri, C.; Ligresti, A.; Di Marzo, V. Peripheral Effects of the Endocannabinoid System in Energy Homeostasis: Adipose Tissue, Liver and Skeletal Muscle. Rev. Endocr. Metab. Disord. 2011, 12, 153–162. [Google Scholar] [CrossRef]
- Hammell, D.C.; Zhang, L.P.; Ma, F.; Abshire, S.M.; McIlwrath, S.L.; Stinchcomb, A.L.; Westlund, K.N. Transdermal Cannabidiol Reduces Inflammation and Pain-Related Behaviours in a Rat Model of Arthritis. Eur. J. Pain. 2016, 20, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, J.; Rosado, T.; Soares, S.; Simão, A.Y.; Caramelo, D.; Luís, Â.; Fernández, N.; Barroso, M.; Gallardo, E.; Duarte, A.P. Cannabis and Its Secondary Metabolites: Their Use as Therapeutic Drugs, Toxicological Aspects, and Analytical Determination. Medicines 2019, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guard, S.E.; Chapnick, D.A.; Poss, Z.; Ebmeier, C.C.; Jacobsen, J.; Nemkov, T.; Ball, K.A.; Webb, K.J.; Simpson, H.L.; Coleman, S.; et al. Multi-Omic Analysis Reveals Cannabidiol Disruption of Cholesterol Homeostasis in Human Cell Lines. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kraemer, M.; Broecker, S.; Madea, B.; Hess, C. Decarbonylation: A Metabolic Pathway of Cannabidiol in Humans. Drug Test. Anal. 2019, 11, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Usami, N.; Osada, S.; Narimatsu, S.; Yamamoto, I.; Yoshimura, H. Cannabidiol Metabolism Revisited: Tentative Identification of Novel Decarbonylated Metabolites of Cannabidiol Formed by Human Liver Microsomes and Recombinant Cytochrome P450 3A4. Forensic Toxicol. 2019, 37, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Mechoulam, R.; Tchilibon, S.; Fride, E.; Hanus, L.; Breuer, A.; Gallily, R. Pharmaceutical Compositions Comprising Cannabidiol Derivatives. U.S. Patent 7,759,526, 20 July 2010. [Google Scholar]
- Kozela, E.; Haj, C.; Hanuš, L.; Chourasia, M.; Shurki, A.; Juknat, A.; Kaushansky, N.; Mechoulam, R.; Vogel, Z. HU-446 and HU-465, Derivatives of the Non-Psychoactive Cannabinoid Cannabidiol, Decrease the Activation of Encephalitogenic T Cells. Chem. Biol. Drug Des. 2016, 87, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Hampson, A.J.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (-)Delta9-Tetrahydrocannabinol Are Neuroprotective Antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhukova, N.V.; Karaman, Y.K.; Zhukova, N.V. The Effect of High Fat Food on Erythrocyte Phospholipids, Fatty Acids Composition and Glutathione Redox-System of Rats with Alimentary Dyslipidemia. Health 2010, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.P.; Maiorino, M.; Ursini, F.; Girotti, A.W. Protective Action of Phospholipid Hydroperoxide Glutathione Peroxidase against Membrane-Damaging Lipid Peroxidation. In Situ Reduction of Phospholipid and Cholesterol Hydroperoxides. J. Biol. Chem. 1990, 265, 454–461. [Google Scholar] [CrossRef]
- Wang, Y.; Mukhopadhyay, P.; Cao, Z.; Wang, H.; Feng, D.; Haskó, G.; Mechoulam, R.; Gao, B.; Pacher, P. Cannabidiol Attenuates Alcohol-Induced Liver Steatosis, Metabolic Dysregulation, Inflammation and Neutrophil-Mediated Injury. Sci. Rep. 2017, 7, 12064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bih, C.I.; Chen, T.; Nunn, A.V.W.; Bazelot, M.; Dallas, M.; Whalley, B.J. Molecular Targets of Cannabidiol in Neurological Disorders. Neurotherapeutics 2015, 12, 699–730. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.; Starosta, V.; Maier, K.; Beck-Speier, I.; Rebhan, C.; Becker, B.F.; Latzin, P.; Fischer, R.; Ratjen, F.; Huber, R.M.; et al. Inhaled Glutathione Decreases PGE2 and Increases Lymphocytes in Cystic Fibrosis Lungs. Free. Radic. Biol. Med. 2005, 39, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-A.; Kim, E.-Y.; Song, H.; Kim, C.; Kim, J.-H. Reactive Oxygen Species Are Generated through a BLT2-Linked Cascade in Ras-Transformed Cells. Free. Radic. Biol. Med. 2008, 44, 624–634. [Google Scholar] [CrossRef]
- Kim, E.-Y.; Seo, J.-M.; Cho, K.-J.; Kim, J.-H. Ras-Induced Invasion and Metastasis Are Regulated by a Leukotriene B4 Receptor BLT2-Linked Pathway. Oncogene 2010, 29, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Poeckel, D.; Funk, C.D. The 5-Lipoxygenase/Leukotriene Pathway in Preclinical Models of Cardiovascular Disease. Cardiovasc. Res. 2010, 86, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Massey, K.A.; Nicolaou, A. Lipidomics of Polyunsaturated-Fatty-Acid-Derived Oxygenated Metabolites. Biochem. Soc. Trans. 2011, 39, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ternowitz, T.; Fogh, K.; Kragballe, K. 15-Hydroxyeicosatetraenoic Acid (15-HETE) Specifically Inhibits LTB4-Induced Chemotaxis of Human Neutrophils. Ski. Pharmacol. Physiol. 1988, 1, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Blunder, S.; Rühl, R.; Moosbrugger-Martinz, V.; Krimmel, C.; Geisler, A.; Zhu, H.; Crumrine, D.; Elias, P.M.; Gruber, R.; Schmuth, M.; et al. Alterations in Epidermal Eicosanoid Metabolism Contribute to Inflammation and Impaired Late Differentiation in FLG-Mutated Atopic Dermatitis. J. Investig. Dermatol. 2017, 137, 706–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Chang, N.; Tian, L.; Yang, J.; Ji, X.; Xie, J.; Yang, L.; Li, L. MiR-27b-3p, MiR-181a-1-3p, and MiR-326-5p Are Involved in the Inhibition of Macrophage Activation in Chronic Liver Injury. J. Mol. Med. 2017, 95, 1091–1105. [Google Scholar] [CrossRef]
- Abdo, H.; Mahé, M.M.; Derkinderen, P.; Bach-Ngohou, K.; Neunlist, M.; Lardeux, B. The Omega-6 Fatty Acid Derivative 15-Deoxy-Δ12,14-Prostaglandin J2 Is Involved in Neuroprotection by Enteric Glial Cells against Oxidative Stress. J. Physiol. 2012, 590, 2739–2750. [Google Scholar] [CrossRef] [Green Version]
- Bie, Q.; Dong, H.; Jin, C.; Zhang, H.; Zhang, B. 15d-PGJ2 Is a New Hope for Controlling Tumor Growth. Am. J. Transl. Res. 2018, 10, 648–658. [Google Scholar]
- Berman, P.; Sulimani, L.; Gelfand, A.; Amsalem, K.; Lewitus, G.M.; Meiri, D. Cannabinoidomics—An Analytical Approach to Understand the Effect of Medical Cannabis Treatment on the Endocannabinoid Metabolome. Talanta 2020, 219, 121336. [Google Scholar] [CrossRef]
- Bisogno, T.; Howell, F.; Williams, G.; Minassi, A.; Cascio, M.G.; Ligresti, A.; Matias, I.; Schiano-Moriello, A.; Paul, P.; Williams, E.-J.; et al. Cloning of the First Sn1-DAG Lipases Points to the Spatial and Temporal Regulation of Endocannabinoid Signaling in the Brain. J. Cell Biol. 2003, 163, 463–468. [Google Scholar] [CrossRef]
- Artmann, A.; Petersen, G.; Hellgren, L.I.; Boberg, J.; Skonberg, C.; Nellemann, C.; Hansen, S.H.; Hansen, H.S. Influence of Dietary Fatty Acids on Endocannabinoid and N-Acylethanolamine Levels in Rat Brain, Liver and Small Intestine. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2008, 1781, 200–212. [Google Scholar] [CrossRef]
- Cao, Z.; Mulvihill, M.M.; Mukhopadhyay, P.; Xu, H.; Erdélyi, K.; Hao, E.; Holovac, E.; Haskó, G.; Cravatt, B.F.; Nomura, D.K.; et al. Monoacylglycerol Lipase Controls Endocannabinoid and Eicosanoid Signaling and Hepatic Injury in Mice. Gastroenterology 2013, 144, 808–817,e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biernacki, M.; Jastrząb, A.; Skrzydlewska, E. Changes in Hepatic Phospholipid Metabolism in Rats under UV Irradiation and Topically Treated with Cannabidiol. Antioxidants 2021, 10, 1157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081157
Biernacki M, Jastrząb A, Skrzydlewska E. Changes in Hepatic Phospholipid Metabolism in Rats under UV Irradiation and Topically Treated with Cannabidiol. Antioxidants. 2021; 10(8):1157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081157
Chicago/Turabian StyleBiernacki, Michał, Anna Jastrząb, and Elżbieta Skrzydlewska. 2021. "Changes in Hepatic Phospholipid Metabolism in Rats under UV Irradiation and Topically Treated with Cannabidiol" Antioxidants 10, no. 8: 1157. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081157