Cigarette Smoke Extract Stimulates MMP-2 Production in Nasal Fibroblasts via ROS/PI3K, Akt, and NF-κB Signaling Pathways

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of CSE
2.2. Patients and Tissue Collection
2.3. Nasal Fibroblast Cultures
2.4. Real-Time PCR
2.5. Gelatin Zymography
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Western Blotting Analysis
2.8. Immunofluorescent Staining
2.9. Measurement of ROS Produced
2.10. Statistical Data Analysis
3. Results
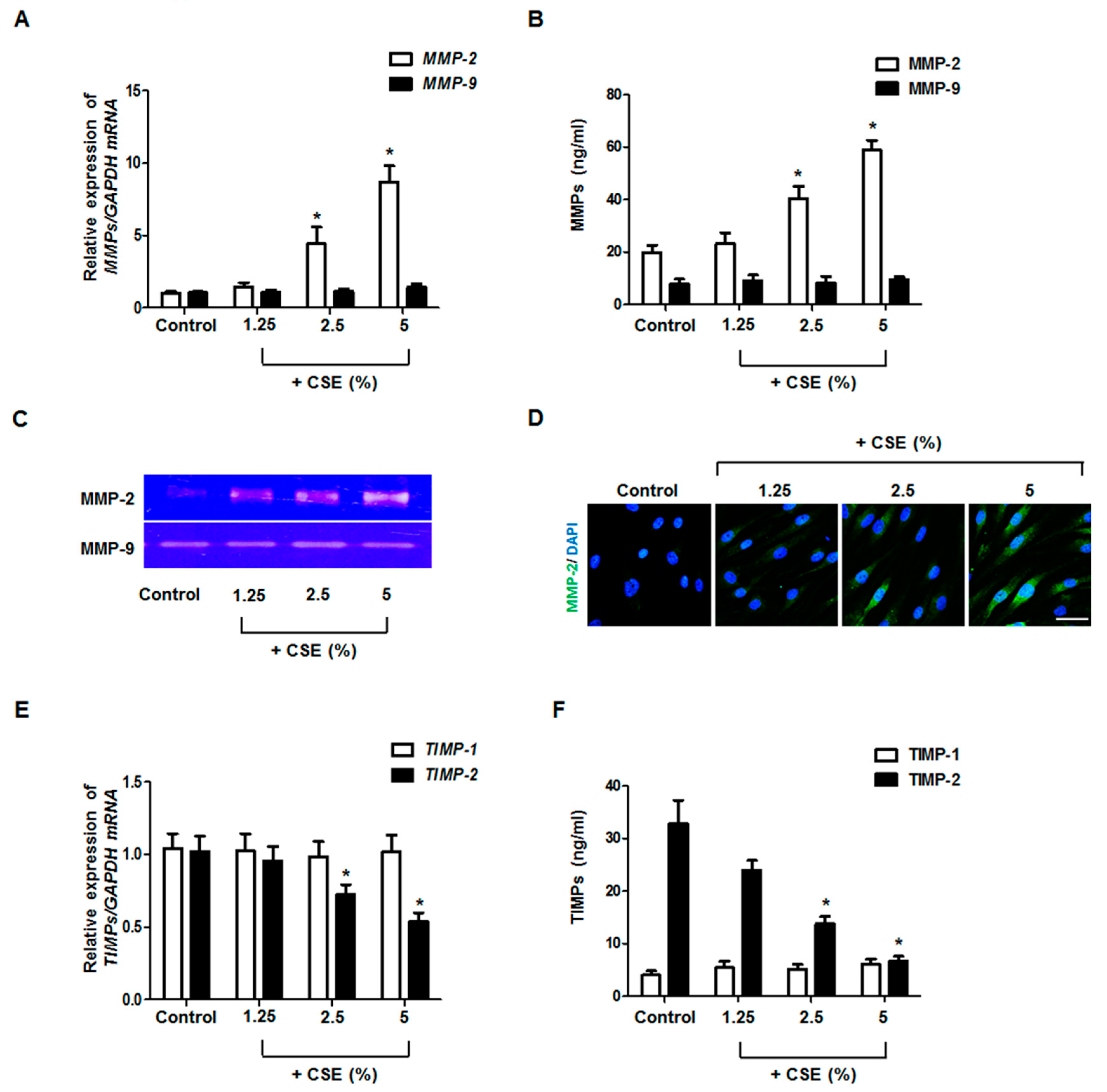
3.1. Effects of CSE on MMP and TIMP Production in Nasal Fibroblasts
3.2. Role of ROS in MMP and TIMP Production in CSE-Stimulated Nasal Fibroblasts
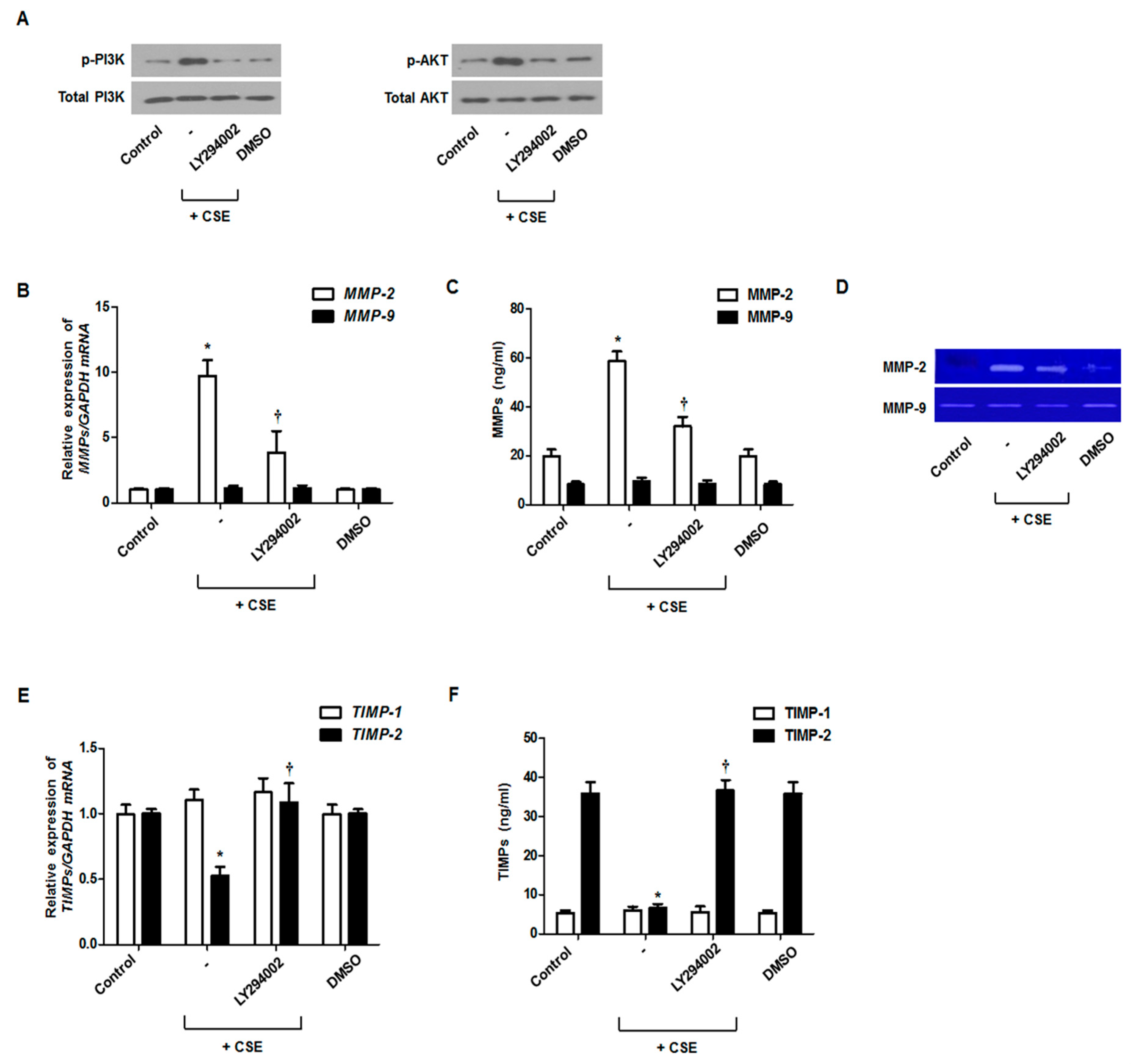
3.3. Involvement of the PI3K/Akt Cascade in MMP and TIMP Production in CSE-Stimulated Nasal Fibroblasts
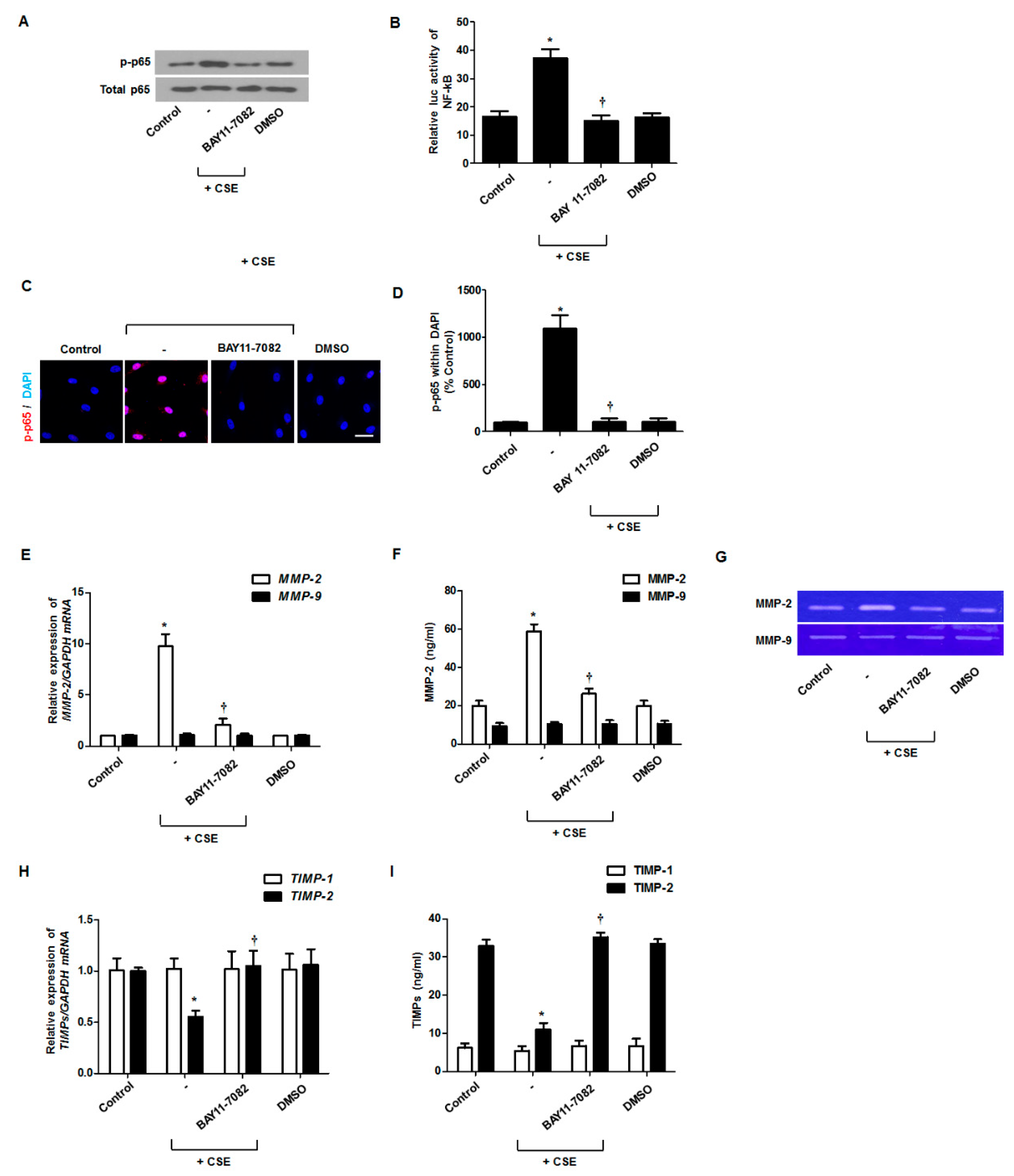
3.4. Effect of CSE on NF-κB Activation for MMP and TIMP Production
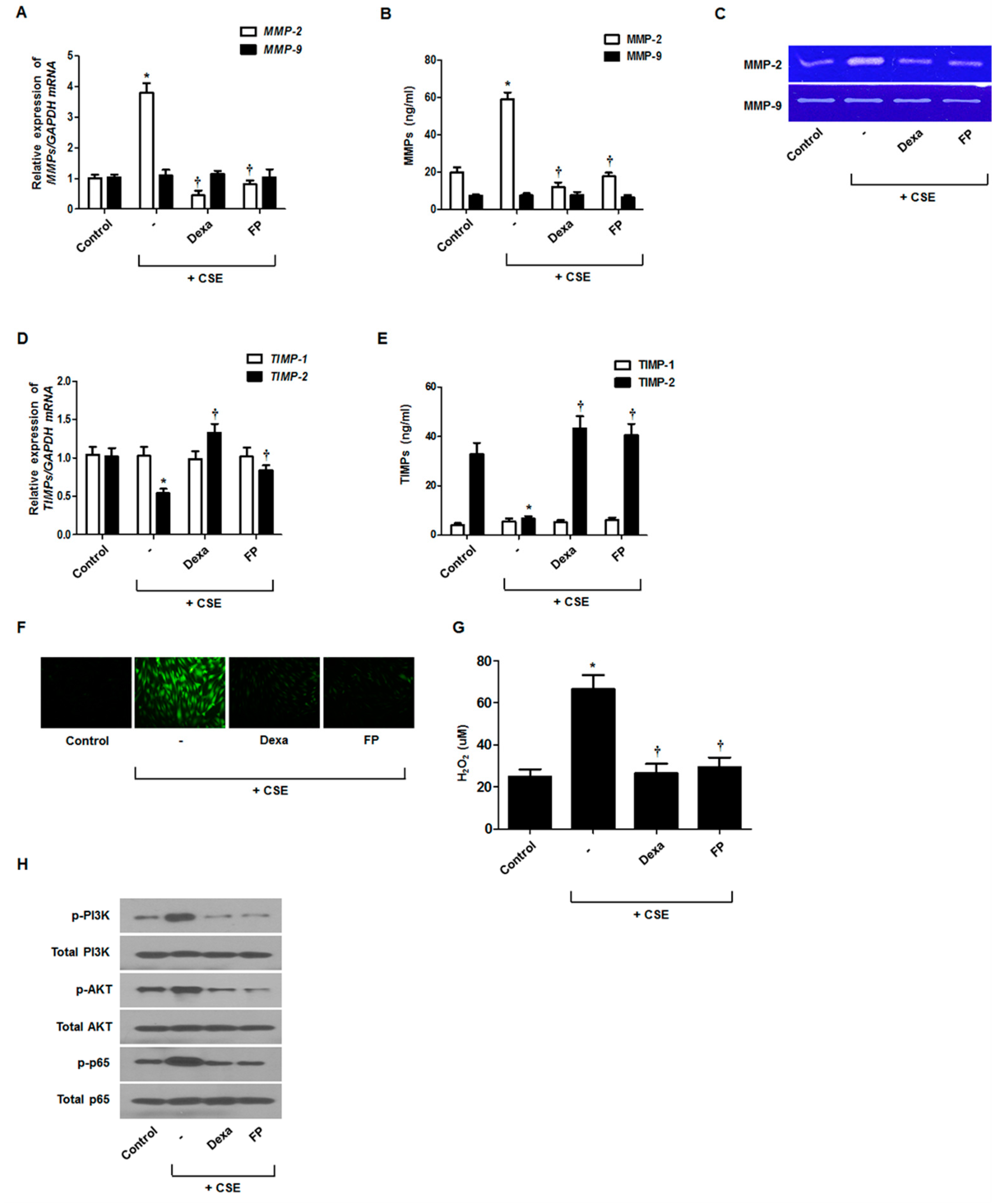
3.5. Effect of Steroids on CSE-Regulated MMP and TIMP Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lange, B.; Holst, R.; Thilsing, T.; Baelum, J.; Kjeldsen, A. Quality of life and associated factors in persons with chronic rhinosinusitis in the general population: A prospective questionnaire and clinical cross-sectional study. Clin. Otolaryngol. 2013, 38, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Reh, D.D.; Higgins, T.S.; Smith, T.L. Impact of tobacco smoke on chronic rhinosinusitis: A review of the literature. Int. Forum Allergy Rhinol. 2012, 2, 362–369. [Google Scholar] [CrossRef]
- Gueders, M.M.; Foidart, J.M.; Noel, A.; Cataldo, D.D. Matrix metalloproteinases (MMPs) and tissue inhibitors of MMPs in the respiratory tract: Potential implications in asthma and other lung diseases. Eur. J. Pharmacol. 2006, 533, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Van Bruaene, N.; Bachert, C. Tissue remodeling in chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care (New Rochelle) 2015, 4, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Langhammer, T.; Westhofen, M.; Lorenzen, J. Relationship between matrix metalloproteinases MMP-2, MMP-9, tissue inhibitor of matrix metalloproteinases-1 and IL-5, IL-8 in nasal polyps. Allergy 2007, 62, 66–72. [Google Scholar] [CrossRef]
- Wang, R.D.; Wright, J.L.; Churg, A. Transforming growth factor-beta (1) drives airway remodeling in cigarette smoke-exposed tracheal explants. Am. J. Resp. Cell. Mol. 2005, 33, 387–393. [Google Scholar] [CrossRef]
- Jia, M.; Chen, X.; Liu, J.; Chen, J. PTEN promotes apoptosis of H2O2-injured rat nasal epithelial cells through PI3K/Akt and other pathways. Mol. Med. Rep. 2018, 17, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Dagli, M.; Eryilmaz, A.; Besler, T.; Akmansu, H.; Acar, A.; Korkmaz, H. Role of free radicals and antioxidants in nasal polyps. Laryngoscope 2004, 114, 1200–1203. [Google Scholar] [CrossRef]
- Zeng, H.; Wang, Y.; Gu, Y.; Wang, J.; Zhang, H.; Gao, H.; Jin, Q.; Zhao, L. Polydatin attenuates reactive oxygen species-induced airway remodeling by promoting Nrf2-mediated antioxidant signaling in asthma mouse model. Life Sci. 2019, 218, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Al-Azzam, N.; Teegala, L.R.; Pokhrel, S.; Ghebreigziabher, S.; Chachkovskyy, T.; Thodeti, S.; Gavilanes, I.; Covington, K.; Thodeti, C.K.; Paruchuri, S. Transient Receptor Potential Vanilloid channel regulates fibroblast differentiation and airway remodeling by modulating redox signals through NADPH Oxidase 4. Sci. Rep. 2020, 10, 9827. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.S.; Lv, Y.; Wang, Z.; Ma, J.; Mi, C.; Li, X.; Xu, G.H.; Piao, L.X.; Zheng, S.Z.; Jin, X. Imperatorin efficiently blocks TNF-α-mediated activation of ROS/PI3K/Akt/NF-κB pathway. Oncol. Rep. 2017, 37, 3397–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.F.; Yuan, Q.; Ye, J. Relationship and significance among IL-6,PI3K/Akt and GSK 3β in chronic rhinosinusitis. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi J. Clin. Otorhinolaryngol. Head Neck Surg. 2016, 30, 1859–1864. [Google Scholar] [CrossRef]
- Jung, C.H.; Kim, E.M.; Park, J.K.; Hwang, S.G.; Moon, S.K.; Kim, W.J.; Um, H.D. Bmal1 suppresses cancer cell invasion by blocking the phosphoinositide 3-kinase-Akt-MMP-2 signaling pathway. Oncol. Rep. 2013, 29, 2109–2113. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.R.; Chida, A.S.; Bauter, M.R.; Shafiq, N.; Seweryniak, K.; Maggirwar, S.B.; Kilty, I.; Rahman, I. Cigarette smoke induces proinflammatory cytokine release by activation of NF-kappa B and posttranslational modifications of histone deacetylase in macrophages. Am. J. Physiol. Lung C. 2006, 291, L46–L57. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Kang, J.H.; Um, J.Y.; Han, I.H.; Park, I.H.; Lee, H.M. Lipopolysaccharide Induces Pro-Inflammatory Cytokines and MMP Production via TLR4 in Nasal Polyp-Derived Fibroblast and Organ Culture. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Léguillette, R.; Tohver, T.; Bond, S.L.; Nicol, J.A.; McDonald, K.J. Effect of Dexamethasone and Fluticasone on Airway Hyperresponsiveness in Horses with Inflammatory Airway Disease. J. Vet. Intern. Med. 2017, 31, 1193–1201. [Google Scholar] [CrossRef]
- Hadar, T.; Yaniv, E.; Shvili, Y.; Koren, R.; Shvero, J. Histopathological changes of the nasal mucosa induced by smoking. Inhal. Toxicol. 2009, 21, 1119–1122. [Google Scholar] [CrossRef]
- Amin, K.A.M. Allergic Respiratory Inflammation and Remodeling. Turk. Thorac. J. 2015, 16, 133–140. [Google Scholar] [CrossRef]
- Christensen, D.N.; Franks, Z.G.; McCrary, H.C.; Saleh, A.A.; Chang, E.H. A Systematic Review of the Association between Cigarette Smoke Exposure and Chronic Rhinosinusitis. Otolaryng. Head Neck 2018, 158, 801–816. [Google Scholar] [CrossRef] [PubMed]
- Vent, J.; Robinson, A.M.; Gentry-Nielsen, M.J.; Conley, D.B.; Hallworth, R.; Leopold, D.A.; Kern, R.C. Pathology of the olfactory epithelium: Smoking and ethanol exposure. Laryngoscope 2004, 114, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; Deng, J.S.; Huang, S.S.; Li, P.Y.; Liang, Y.C.; Huang, G.J. 3, 4-dihydroxybenzalacetone attenuates lipopolysaccharide-induced inflammation in acute lung injury via down-regulation of MMP-2 and MMP-9 activities through suppressing ROS-mediated MAPK and PI3K/AKT signaling pathways. Int. Immunopharmacol. 2017, 50, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Meng, J.; Qiao, X.M.; Liu, Y.F.; Liu, F.; Zhang, N.; Zhang, J.; Holtappels, G.; Luo, B.; Zhou, P.; et al. Expression of TGF, matrix metalloproteinases, and tissue inhibitors in Chinese chronic rhinosinusitis. J. Allergy Clin. Immun. 2010, 125, 1061–1068. [Google Scholar] [CrossRef]
- Shin, J.M.; Park, J.H.; Kang, B.; Lee, S.A.; Park, I.H.; Lee, H.M. Effect of doxycycline on transforming growth factor-beta-1-induced matrix metalloproteinase 2 expression, migration, and collagen contraction in nasal polyp-derived fibroblasts. Am. J. Rhinol. Allergy 2016, 30, 385–390. [Google Scholar] [CrossRef]
- Fordham, M.T.; Mulligan, J.K.; Casey, S.E.; Mulligan, R.M.; Wang, E.W.; Sansoni, E.R.; Schlosser, R.J. Reactive Oxygen Species in Chronic Rhinosinusitis and Secondhand Smoke Exposure. Otolaryng. Head Neck 2013, 149, 633–638. [Google Scholar] [CrossRef]
- Platt, M.P.; Soler, Z.; Metson, R.; Stankovic, K.M. Pathways analysis of molecular markers in chronic sinusitis with polyps. Otolaryngol. Head Neck Surg. 2011, 144, 802–808. [Google Scholar] [CrossRef]
- Liu, H.J.; Dai, X.N.; Cheng, Y.S.; Fang, S.C.; Zhang, Y.M.; Wang, X.G.; Zhang, W.; Liao, H.; Yao, H.H.; Chao, J. MCPIP1 mediates silica-induced cell migration in human pulmonary fibroblasts. Am. J. Physiol. Lung C. 2016, 310, L121–L132. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Ryter, S.W.; Kyung, S.Y.; Lee, S.P.; Jeong, S.H. The phosphodiesterase 4 inhibitor rolipram protects against cigarette smoke extract-induced apoptosis in human lung fibroblasts. Eur. J. Pharm. 2013, 706, 76–83. [Google Scholar] [CrossRef]
- Krimmer, D.I.; Burgess, J.K.; Wooi, T.K.; Black, J.L.; Oliver, B.G.G. Matrix Proteins from Smoke-Exposed Fibroblasts Are Pro-proliferative. Am. J. Resp. Cell Mol. 2012, 46, 34–39. [Google Scholar] [CrossRef]
- Hou, G.; Yin, Y.; Han, D.; Wang, Q.Y.; Kang, J. Rosiglitazone attenuates the metalloprotease/anti-metalloprotease imbalance in emphysema induced by cigarette smoke: Involvement of extracellular signal-regulated kinase and NF kappa B signaling. Int. J. Chronic Obs. 2015, 10, 715–724. [Google Scholar] [CrossRef] [Green Version]
- Todorova, L.; Bjermer, L.; Westergren-Thorsson, G.; Miller-Larsson, A. TGFbeta-induced matrix production by bronchial fibroblasts in asthma: Budesonide and formoterol effects. Respir. Med. 2011, 105, 1296–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gero, D.; Szabo, C. Glucocorticoids Suppress Mitochondrial Oxidant Production via Upregulation of Uncoupling Protein 2 in Hyperglycemic Endothelial Cells. PLoS ONE 2016, 11, e154813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoh, V.O.; Felty, Q.; Parkash, J.; Poppiti, R.; Roy, D. Reactive oxygen species via redox signaling to PI3K/AKT pathway contribute to the malignant growth of 4-hydroxy estradiol-transformed mammary epithelial cells. PLoS ONE 2013, 8, e54206. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | |
|---|---|---|
| MMP-2 | Forward | 5’- AGA TCT TCT TCT TCA AGG AAC CGT T -3’ |
| Reverse | 5’- GGC TGG TCA GTG GCT TGG GGT A -3’ | |
| MMP-9 | Forward | 5’- GCG GAG ATT GGG AAC CAG CTG TA -3’ |
| Reverse | 5’- GAC GCG CCT GTG TAC ACC CAC A -3’ | |
| TIMP-1 | Forward | 5’- ACC ACC TTA TAC CAG CGT TAT GA -3’ |
| Reverse | 5’- GGT GTA GAC GAA CCG GAT GTC -3’ | |
| TIMP-2 | Forward | 5’- GCT GCG AGT GCA AGA TCA C -3’ |
| Reverse | 5’- TGG TGC CCG TTG ATG TTC TTC | |
| TLR4 | Forward | 5’- TGA GCA GTC GTG CTG GTA TC -3’ |
| Reverse | 5’- CAG GGC TTT TCT GAG TCG TC -3’ | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-H.; Shin, J.-M.; Yang, H.-W.; Kim, T.H.; Lee, S.H.; Lee, H.-M.; Cho, J.-G.; Park, I.-H. Cigarette Smoke Extract Stimulates MMP-2 Production in Nasal Fibroblasts via ROS/PI3K, Akt, and NF-κB Signaling Pathways. Antioxidants 2020, 9, 739. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9080739
Park J-H, Shin J-M, Yang H-W, Kim TH, Lee SH, Lee H-M, Cho J-G, Park I-H. Cigarette Smoke Extract Stimulates MMP-2 Production in Nasal Fibroblasts via ROS/PI3K, Akt, and NF-κB Signaling Pathways. Antioxidants. 2020; 9(8):739. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9080739
Chicago/Turabian StylePark, Joo-Hoo, Jae-Min Shin, Hyun-Woo Yang, Tae Hoon Kim, Seung Hoon Lee, Heung-Man Lee, Jae-Gu Cho, and Il-Ho Park. 2020. "Cigarette Smoke Extract Stimulates MMP-2 Production in Nasal Fibroblasts via ROS/PI3K, Akt, and NF-κB Signaling Pathways" Antioxidants 9, no. 8: 739. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9080739