Melatonin Ameliorates Inflammation and Oxidative Stress by Suppressing the p38MAPK Signaling Pathway in LPS-Induced Sheep Orchitis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Chemicals and Reagents
2.3. Isolation and Culture of Testicular Macrophages
2.4. Leydig Cells Co-Culture with Testicular Macrophages
2.5. Effects of Regionally Injecting LPS with or without Melatonin into Testes
2.6. Quantitative Reverse Transcription—(qRT-) PCR
2.7. Western Blots
2.8. ELISA
2.9. Detection of Metabolites and Proteins Associated with Oxidative Stress
2.10. Statistical Analysis
3. Results
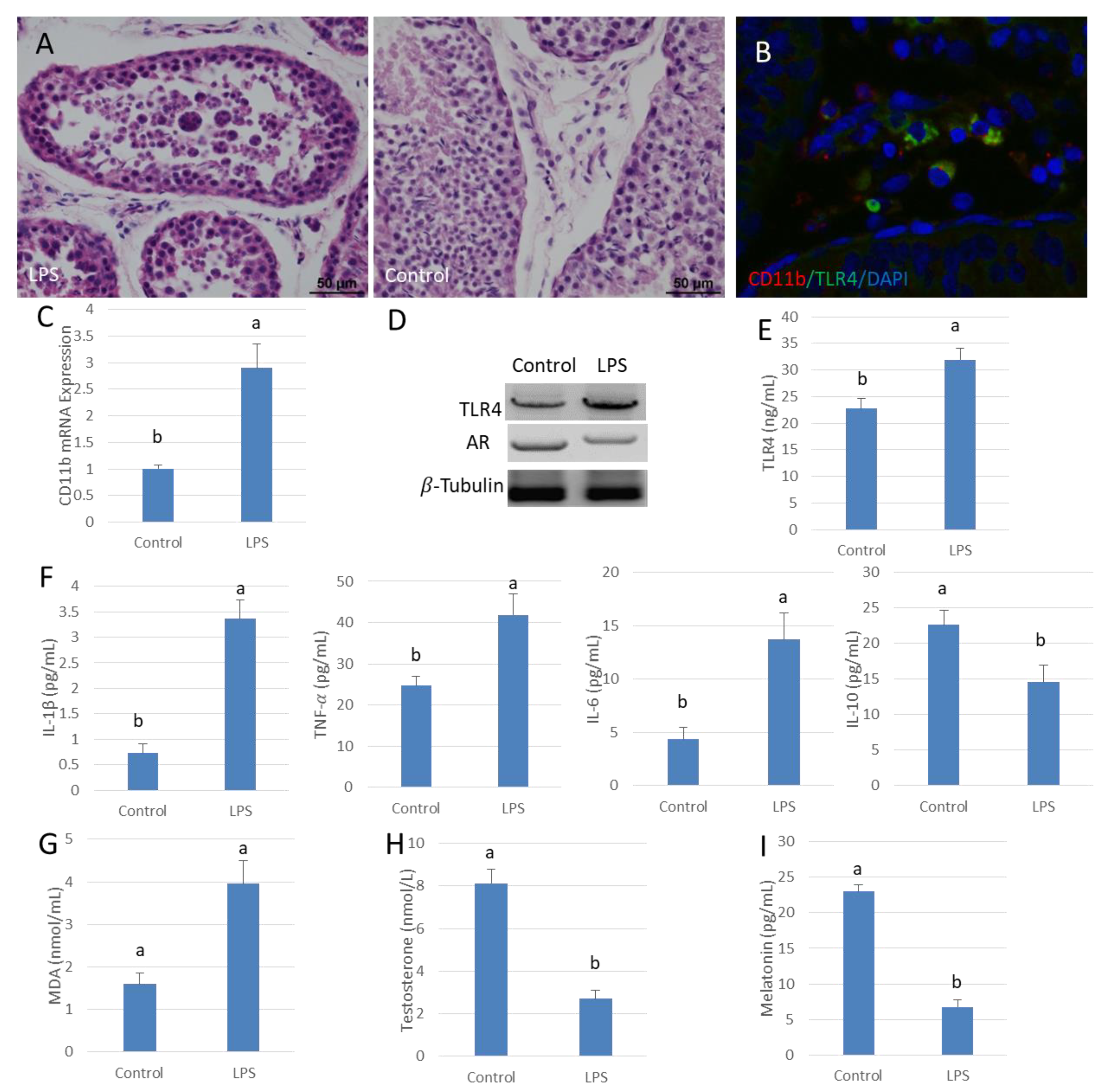
3.1. Low Testosterone Levels in LPS Contributes to Orchitis Tissues
3.2. Melatonin Enhances Phagocytosis of Testicular Macrophages
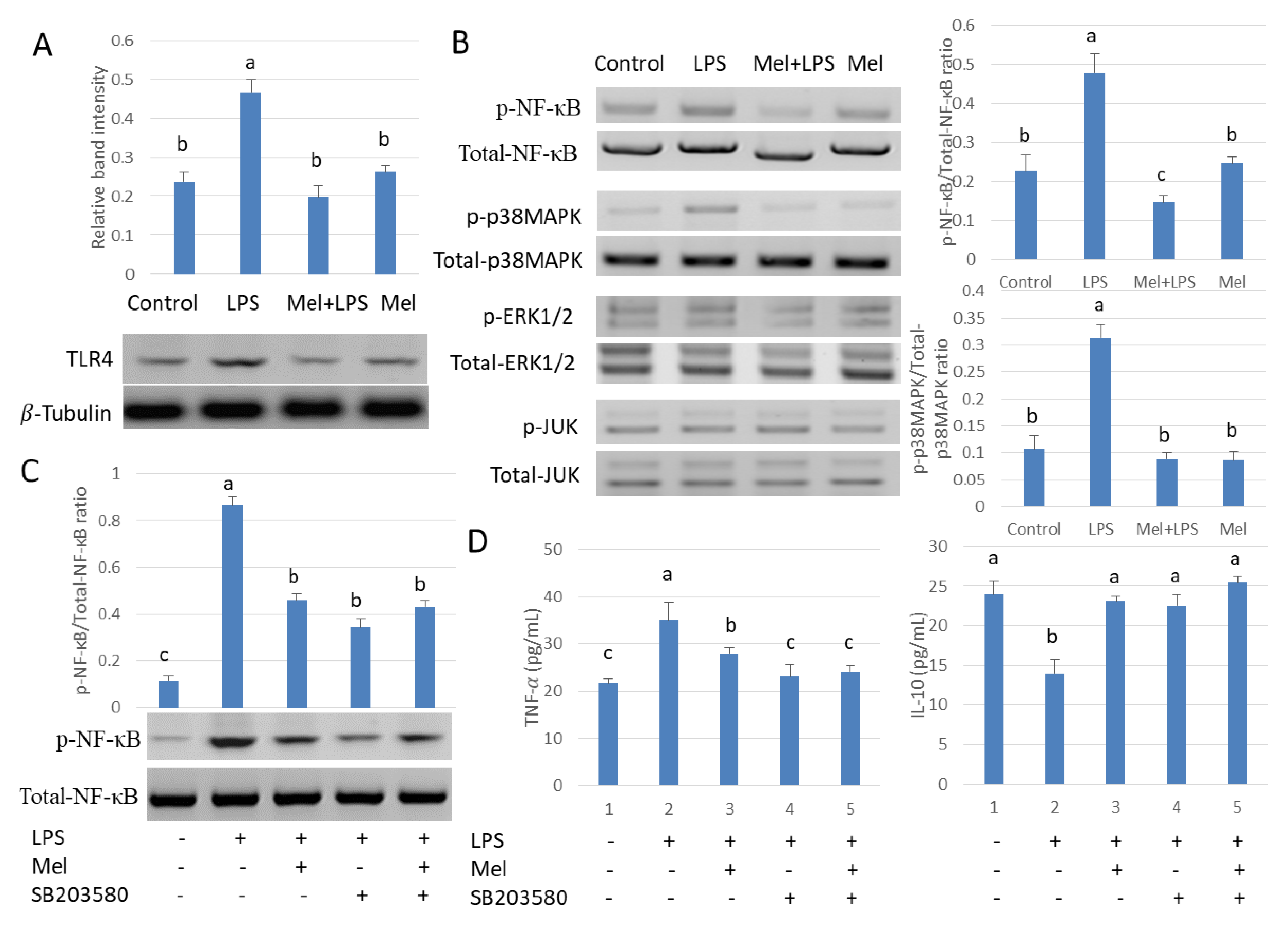
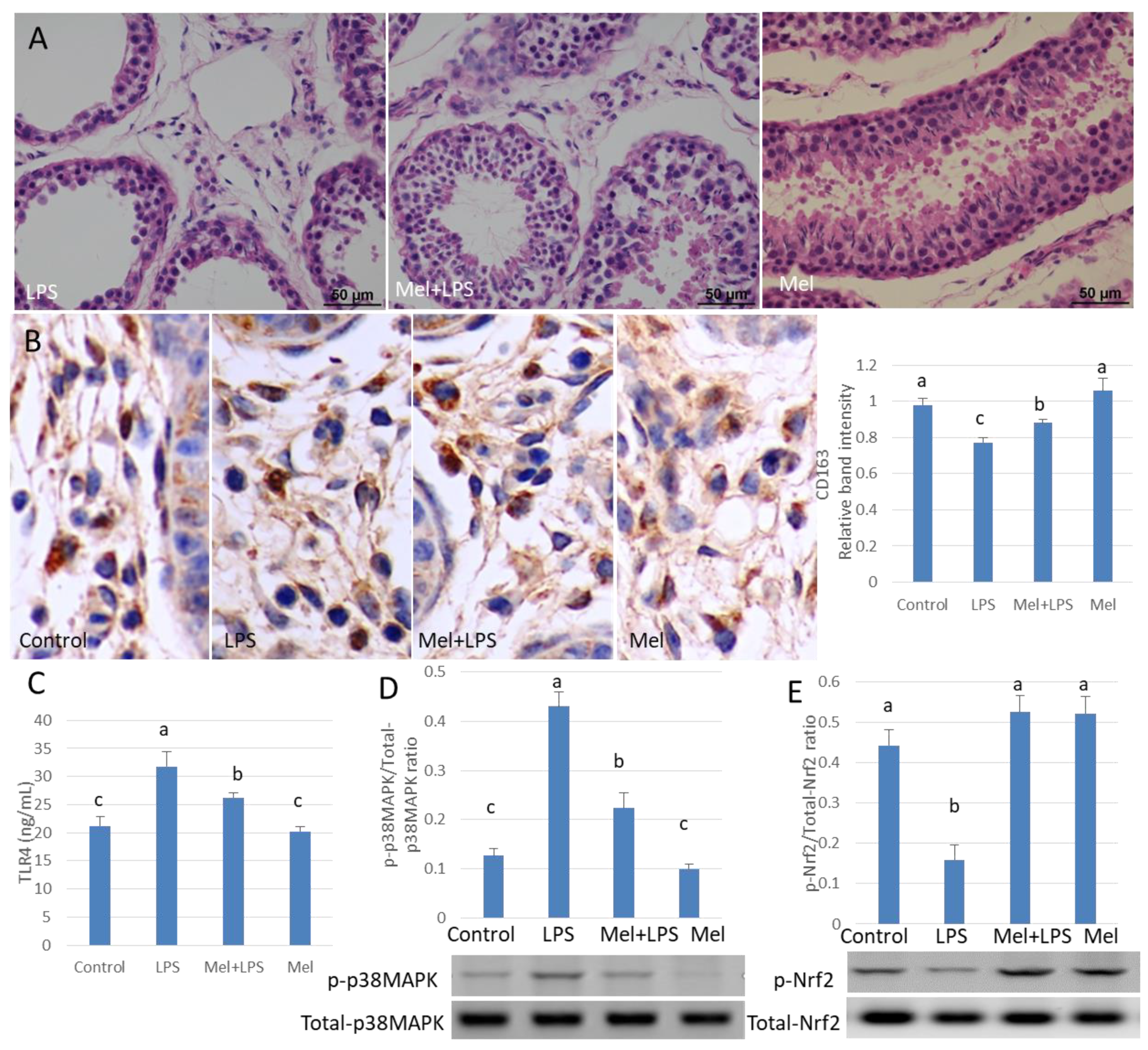
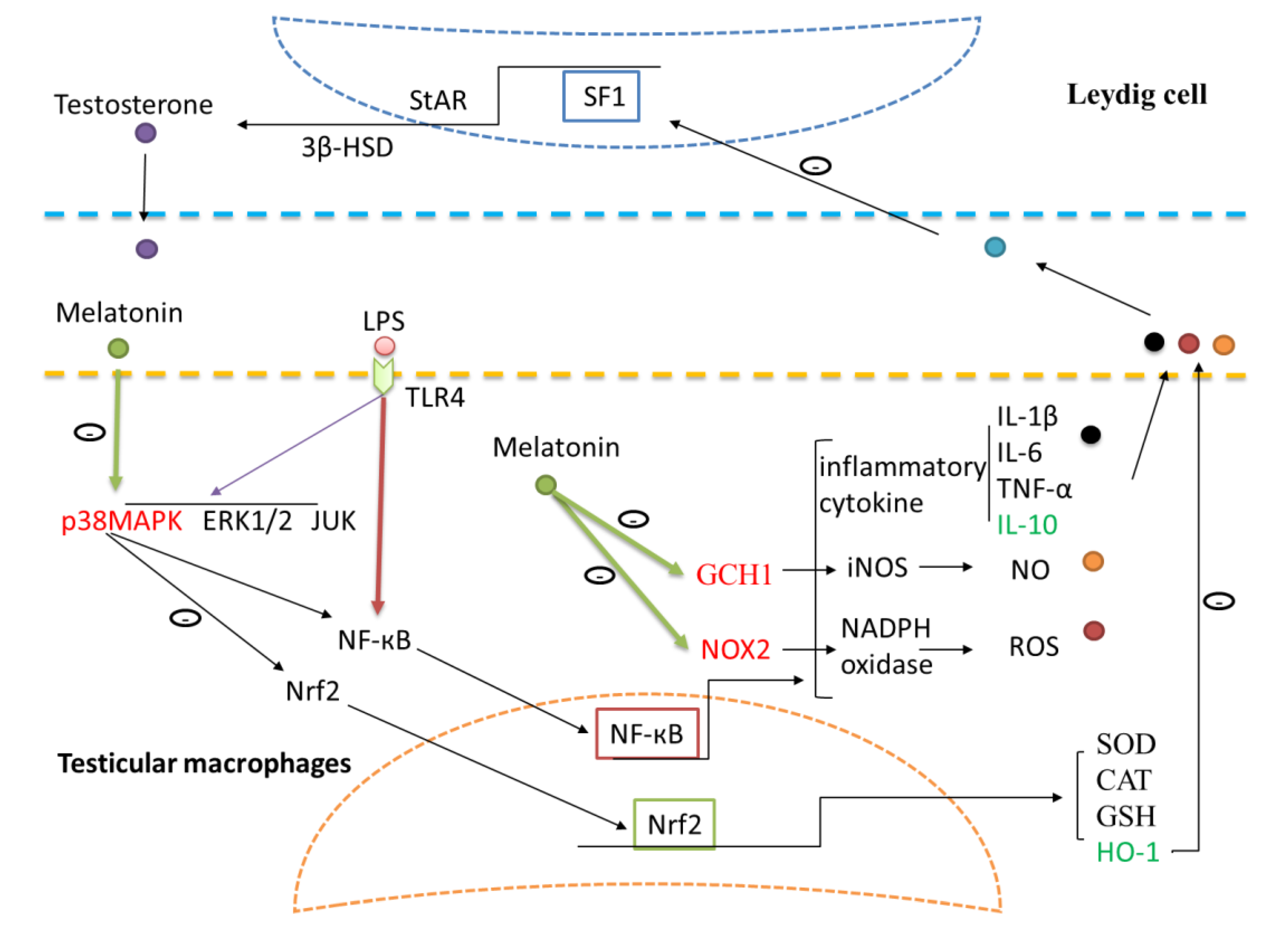
3.3. Melatonin Decreases TLR4-Mediated Inflammatory Genes via Inhibition of p38/MAPK Signaling Pathway
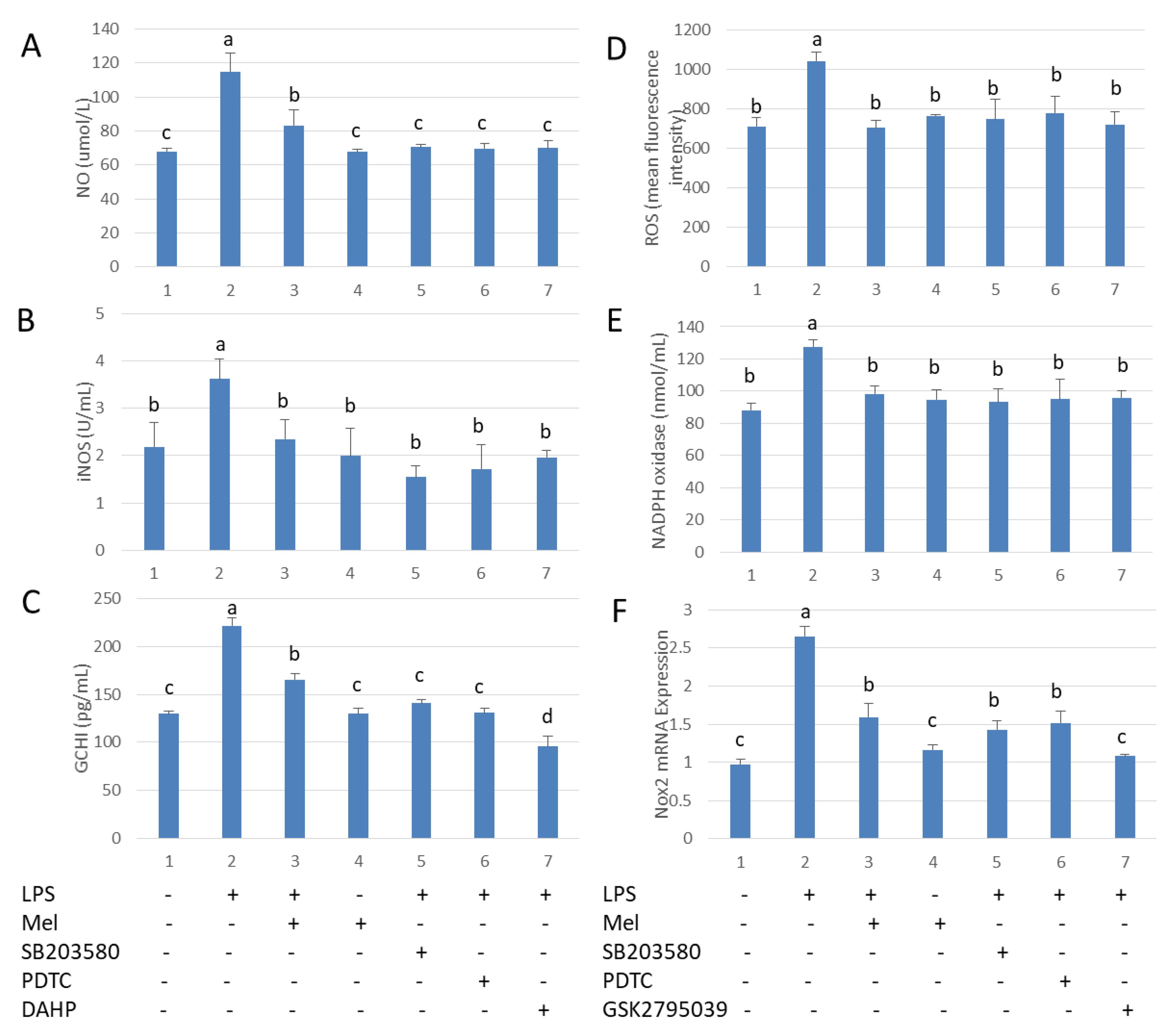
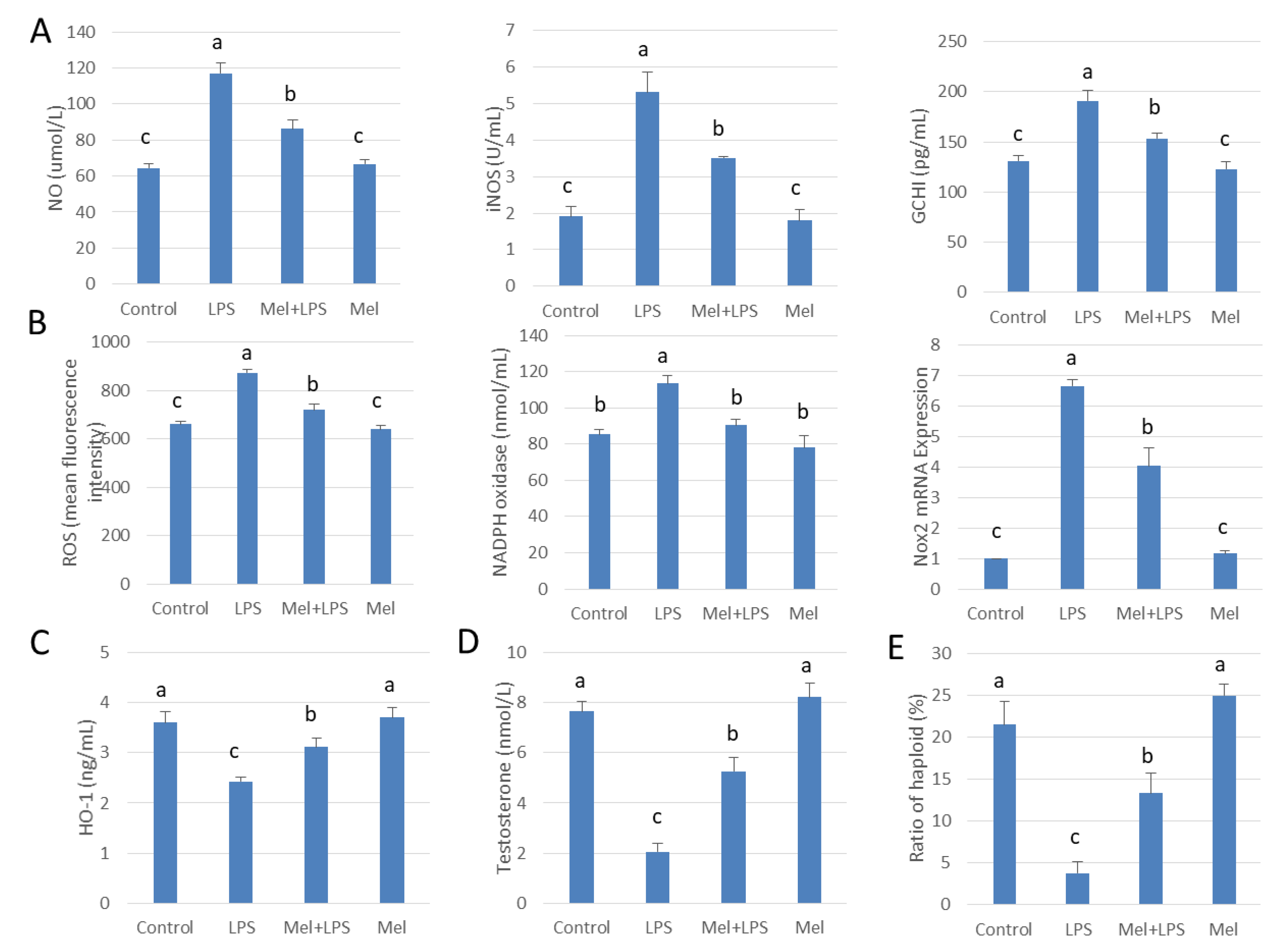
3.4. Melatonin Inhibits GCH1 and Nox2, and Decreased iNOS and NADPH Oxidase Activity in Testicular Macrophages
3.5. Melatonin Attenuates LPS Oxidative Damage of Testicular Macrophages through Nrf2/HO-1 Pathways
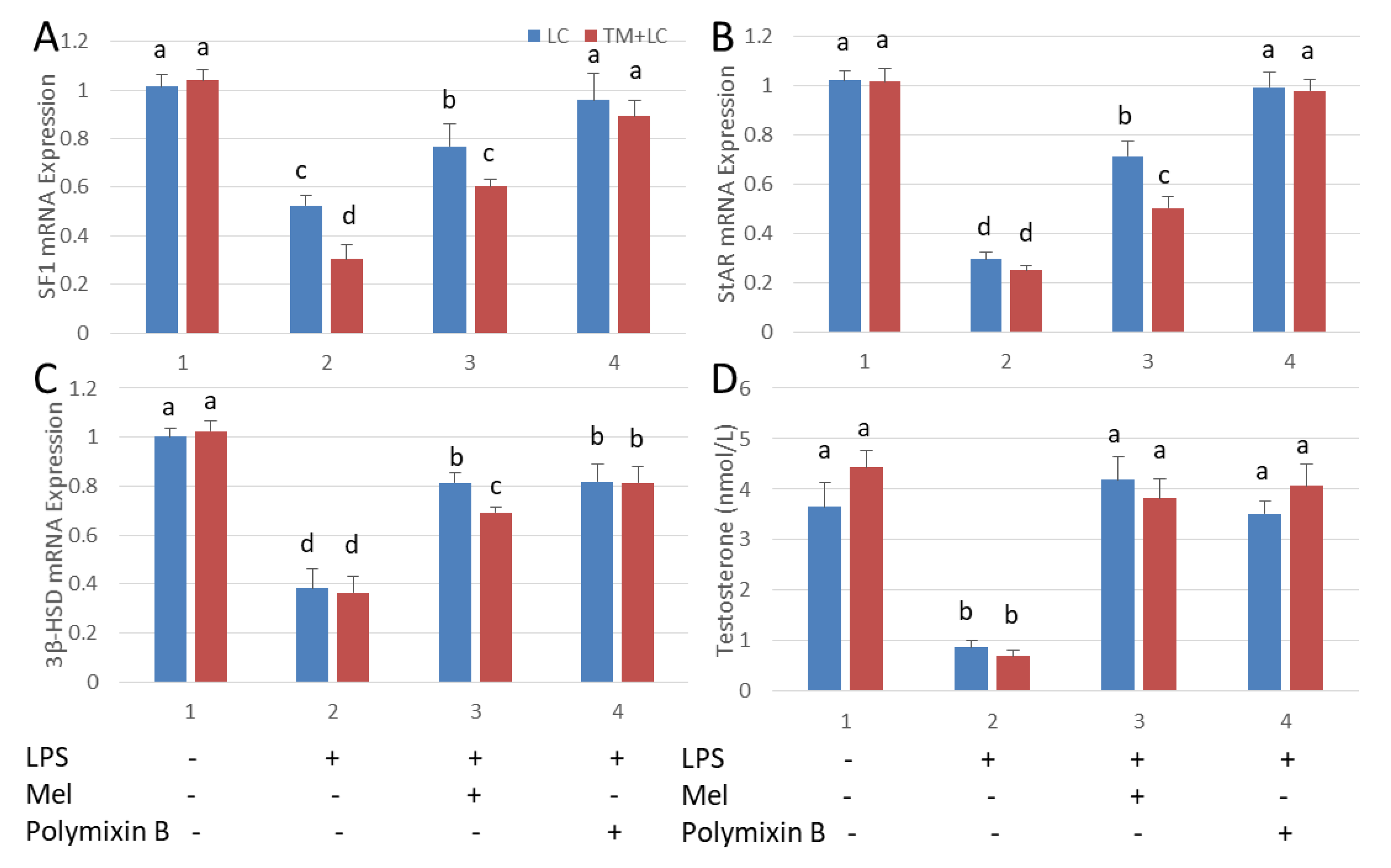
3.6. Melatonin Increases Testosterone Levels in Leydig Cells Co-Cultured with Testicular Macrophages
3.7. Melatonin Alleviates LPS-Induced Orchitis in Sheep
4. Discussions
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Choudhury, A.; Mukherjee, S. In silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J. Med Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Calandra, R.S.; Lustig, L.; Meineke, V.; Köhn, F.M.; Vogt, H.J.; Mayerhofer, A. Number, distribution pattern, and identification of macrophages in the testes of infertile men. Fertil. Steril. 2002, 78, 298–306. [Google Scholar] [CrossRef]
- Mossadegh-Keller, N.; Gentek, R.; Gimenez, G.; Bigot, S.; Mailfert, S.; Sieweke, M.H. Developmental origin and maintenance of distinct testicular macrophage populations. J. Exp. Med. 2017, 214, 2829–2841. [Google Scholar] [CrossRef] [PubMed]
- Aslani, F.; Schuppe, H.-C.; Guazzone, V.A.; Bhushan, S.; Wahle, E.; Lochnit, G.; Lustig, L.; Meinhardt, A.; Fijak, M. Targeting high mobility group box protein 1 ameliorates testicular inflammation in experimental autoimmune orchitis. Hum. Reprod. 2015, 30, 417–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.A.; Diemer, T.; Janus, P.; Hales, K.H.; Hales, D.B. Bacterial Endotoxin Lipopolysaccharide and Reactive Oxygen Species Inhibit Leydig Cell Steroidogenesis via Perturbation of Mitochondria. Endocrine 2004, 25, 265–276. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Kim, M.H.; Han, J.M.; Hong, J.; Lee, T.H.; Kim, S.H.; Yang, W.M. The anti-inflammatory potential of Cortex Phellodendron in vivo and in vitro: Down-regulation of NO and iNOS through suppression of NF-κB and MAPK activation. Int. Immunopharmacol. 2014, 19, 214–220. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jeong, J.-M.; Kim, S.J.; Seo, W.; Kim, M.-H.; Choi, W.-M.; Yoo, W.; Lee, J.-H.; Shim, Y.-R.; Yi, H.-S.; et al. Pro-inflammatory hepatic macrophages generate ROS through NADPH oxidase 2 via endocytosis of monomeric TLR4–MD2 complex. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Deng, S.-L.; Sun, T.-C.; Yu, K.; Wang, Z.-P.; Zhang, B.-L.; Zhang, Y.; Wang, X.-X.; Lian, Z.; Liu, Y.-X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free. Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef]
- Deng, S.L.; Chen, S.R.; Wang, Z.P.; Zhang, Y.; Tang, J.X.; Li, J.; Wang, X.X.; Cheng, J.M.; Jin, C.; Li, X.Y.; et al. Melatonin promotes development of haploid germ cells from early developing spermatogenic cells of Suffolk sheep under in vitro condition. J. Pineal Res. 2016, 60, 435–447. [Google Scholar] [CrossRef]
- Markus, R.P.; Cecon, E.; Pires-Lapa, M.A. Immune-Pineal Axis: Nuclear Factor κB (NF-kB) Mediates the Shift in the Melatonin Source from Pinealocytes to Immune Competent Cells. Int. J. Mol. Sci. 2013, 14, 10979–10997. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Meng, F.; Francis, H.; Wu, N.; Chen, L.; Kennedy, L.; Zhou, T.; Franchitto, A.; Onori, P.; Gaudio, E.; et al. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J. Pineal Res. 2020, 68, e12639. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Deng, S.-L.; Sun, T.-C.; Jiang, W.; Liu, Y.-X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires-Lapa, M.A.; Carvalho-Sousa, C.E.; Cecon, E.; Fernandes, P.A.; Markus, R.P. β-Adrenoceptors Trigger Melatonin Synthesis in Phagocytes. Int. J. Mol. Sci. 2018, 19, 2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frungieri, M.B.; Calandra, R.S.; Rossi, S.P. Local Actions of Melatonin in Somatic Cells of the Testis. Int. J. Mol. Sci. 2017, 18, 1170. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Yu, J.P.; Yu, H.G.; Xu, X.M.; Yu, L.L.; Liu, J.; Luo, H.S. Melatonin reduces inflammatory injury through inhibiting NF-kappaB activation in rats with colitis. Mediat. Inflamm. 2005, 2005, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Soto, M.; Alarcón-De-La-Lastra, C.; Cárdeno, A.; Sánchez-Fidalgo, S.; Sánchez-Hidalgo, M. Melatonin modulates microsomal PGE synthase 1 and NF-E2-related factor-2-regulated antioxidant enzyme expression in LPS-induced murine peritoneal macrophages. Br. J. Pharmacol. 2014, 171, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Gang, S.; Sengupta, M. Functional Status of Testicular Macrophages in an Immunopriviledged Niche in Cadmium Intoxicated Murine Testes. Am. J. Reprod. Immunol. 2014, 72, 14–21. [Google Scholar] [CrossRef]
- Matzkin, M.; Valchi, P.; Riviere, E.; Rossi, S.; Tavalieri, Y.; De Toro, M.M.; Mayerhofer, A.; Bartke, A.; Calandra, R.; Frungieri, M. Aging in the Syrian hamster testis: Inflammatory-oxidative status and the impact of photoperiod. Exp. Gerontol. 2019, 124, 110649. [Google Scholar] [CrossRef]
- Deng, S.-L.; Wu, Q.; Yu, K.; Zhang, Y.; Yao, Y.; Li, W.; Deng, Z.; Liu, G.; Li, W.; Lian, Z. Changes in the Relative Inflammatory Responses in Sheep Cells Overexpressing of Toll-Like Receptor 4 When Stimulated with LPS. PLoS ONE 2012, 7, e47118. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.L.; Wang, Z.P.; Jin, C.; Kang, X.L.; Batool, A.; Zhang, Y.; Li, X.Y.; Wang, X.X.; Chen, S.R.; Chang, C.S.; et al. Melatonin promotes sheep Leydig cell testosterone secretion in a co-culture with Sertoli cells. Theriogenology 2018, 106, 170–177. [Google Scholar] [CrossRef]
- Khan, U.W.; Rai, U. Paracrine role of testicular macrophages in control of Leydig cell activities in the wall lizard, Hemidactylus flaviviridis. Gen. Comp. Endocrinol. 2008, 156, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cao, K.; Liao, Z.; Chen, Y.; Lei, X.; Wei, Q.; Liu, C.; Sun, X.; Yang, Y.; Cai, J.; et al. Monophosphoryl lipid A alleviated radiation-induced testicular injury through TLR4-dependent exosomes. J. Cell. Mol. Med. 2020, 24, 3917–3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Haldar, C. Effect of intra-testicular melatonin injection on testicular functions, local and general immunity of a tropical rodent Funambulus pennanti. Endocrine 2010, 37, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Han, R.; Wu, H.; Han, D. Viral threat to male fertility. Andrologia 2018, 50, e13140. [Google Scholar] [CrossRef] [Green Version]
- Ok, F.; Kaplan, H.M.; Kizilgok, B.; Demir, E. Protective effect of Alpha-Linolenic acid on Lipopolysaccharide-Induced Orchitis in mice. Andrologia 2020, e13667. [Google Scholar] [CrossRef]
- Winnall, W.R.; Muir, J.A.; Hedger, M.P. Rat resident testicular macrophages have an alternatively activated phenotype and constitutively produce interleukin-10 in vitro. J. Leukoc. Biol. 2011, 90, 133–143. [Google Scholar] [CrossRef]
- Winnall, W.R.; Hedger, M.P. Phenotypic and functional heterogeneity of the testicular macrophage population: A new regulatory model. J. Reprod. Immunol. 2013, 97, 147–158. [Google Scholar] [CrossRef]
- Bhushan, S.; Hossain, H.; Lü, Y.; Geißler, A.; Tchatalbachev, S.; Mikulski, Z.; Schuler, G.; Klug, J.; Pilatz, A.; Wagenlehner, F.; et al. Uropathogenic E. coli Induce Different Immune Response in Testicular and Peritoneal Macrophages: Implications for Testicular Immune Privilege. PLoS ONE 2011, 6, e28452. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Abreu-Gonzalez, P.; Marik, P.E.; Dominguez-Rodriguez, A. Therapeutic Algorithm for Use of Melatonin in Patients with COVID-19. Front. Med. 2020, 7, 226. [Google Scholar] [CrossRef]
- Farez, M.F.; Mascanfroni, I.D.; Méndez-Huergo, S.P.; Yeste, A.; Murugaiyan, G.; Garo, L.P.; Aguirre, M.E.B.; Patel, B.; Ysrraelit, M.C.; Zhu, C.; et al. Melatonin Contributes to the Seasonality of Multiple Sclerosis Relapses. Cell 2015, 162, 1338–1352. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, J.; Singh, J.; Lea, R.W.; Skwarlo-Sonta, K. Effect of melatonin on phagocytic activity and intracellular free calcium concentration in testicular macrophages from normal and streptozotocin-induced diabetic rats. Mol. Cell. Biochem. 2005, 275, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.P.; Windschuettl, S.; Matzkin, M.E.; Terradas, C.; Ponzio, R.; Puigdomenech, E.; Levalle, O.; Calandra, R.S.; Mayerhofer, A.; Frungieri, M.B. Melatonin in testes of infertile men: Evidence for anti-proliferative and anti-oxidant effects on local macrophage and mast cell populations. Andrologia 2014, 2, 436–449. [Google Scholar] [CrossRef]
- Tugal, D.; Liao, X.; Jain, M.K. Transcriptional Control of Macrophage Polarization. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1135–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.P.; Fang, X.L.; Fang, N.; Wang, X.B.; Qian, H.Y.; Cao, Z.; Cheng, Y.; Wang, B.N.; Wang, Y. Melatonin ameliorates vascular endothelial dysfunction, inflammation, and atherosclerosis by suppressing the TLR4/NF-κB system in high-fat-fed rabbits. J. Pineal Res. 2013, 55, 388–398. [Google Scholar] [CrossRef]
- Xia, M.-Z.; Liang, Y.-L.; Wang, H.; Chen, X.; Huang, Y.-Y.; Zhang, Z.-H.; Chen, Y.-H.; Zhang, C.; Zhao, M.; Xu, D.-X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88- and TRIF-dependent signaling pathways in lipopolysaccharide-stimulated RAW264.7 cells. J. Pineal Res. 2012, 53, 325–334. [Google Scholar] [CrossRef]
- Chang, M.; Li, Y.; Liu, D.; Zhang, L.; Zhang, H.; Tang, H.; Zhang, H. Melatonin prevents secondary intra-abdominal hypertension in rats possibly through inhibition of the p38 MAPK pathway. Free. Radic. Biol. Med. 2016, 97, 192–203. [Google Scholar] [CrossRef]
- Reiter, R.J.; Sharma, R.; Ma, Q.; Liu, C.; Manucha, W.; Abreu-Gonzalez, P.; Dominguez-Rodriguez, A. Plasticity of glucose metabolism in activated immune cells: Advantages for melatonin inhibition of COVID-19 disease. Melatonin Res. 2020, 3, 362–379. [Google Scholar] [CrossRef]
- Shi, D.; Xiao, X.; Wang, J.; Liu, L.; Chen, W.; Fu, L.; Xie, F.; Huang, W.; Deng, W. Melatonin suppresses proinflammatory mediators in lipopolysaccharide-stimulated CRL1999 cells via targeting MAPK, NF-κB, c/EBPβ, and p300 signaling. J. Pineal Res. 2012, 53, 154–165. [Google Scholar] [CrossRef]
- Liu, J.; Li, G.; Xie, W.J.; Wang, L.; Zhang, R.; Huang, K.S.; Zhou, Q.S.; Chen, D.C. Lipopolysaccharide Stimulates Surfactant Protein-A in Human Renal Epithelial HK-2 Cells through Upregulating Toll-like Receptor 4 Dependent MEK1/2-ERK1/2-NF-κB Pathway. Chin. Med. J. 2017, 130, 1236–1243. [Google Scholar] [CrossRef]
- Bhushan, S.; Tchatalbachev, S.; Lu, Y.; Fröhlich, S.; Fijak, M.; Vijayan, V.; Chakraborty, T.; Meinhardt, A. Differential Activation of Inflammatory Pathways in Testicular Macrophages Provides a Rationale for Their Subdued Inflammatory Capacity. J. Immunol. 2015, 194, 5455–5464. [Google Scholar] [CrossRef] [Green Version]
- Calvo, J.R.; González-Yanes, C.; Maldonado, M.D. The role of melatonin in the cells of the innate immunity: A review. J. Pineal Res. 2013, 55, 103–120. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Feng, D.; Liu, Y.; Xu, M.; Gao, A.; Tian, F.; Zhang, L.; Cui, Y.; Wang, Z.; et al. Alterations in the time course of expression of the Nox family in the brain in a rat experimental cerebral ischemia and reperfusion model: Effects of melatonin. J. Pineal Res. 2014, 57, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Sidarala, V.; Veluthakal, R.; Syeda, K.; Vlaar, C.; Newsholme, P.; Kowluru, A. Phagocyte-like NADPH oxidase (Nox2) promotes activation of p38MAPK in pancreatic β-cells under glucotoxic conditions: Evidence for a requisite role of Ras-related C3 botulinum toxin substrate 1 (Rac1). Biochem. Pharmacol. 2015, 95, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; He, X.; Wang, H.; Wang, Z.; Kelly, G.T.; Wang, X.; Chen, Y.; Wang, T.; Qian, Z. TLR4-NOX2 axis regulates the phagocytosis and killing of Mycobacterium tuberculosis by macrophages. BMC Pulm. Med. 2017, 17, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.-L.; Yu, K.; Zhang, B.; Yao, Y.; Wang, Z.; Zhang, J.; Zhang, X.; Liu, G.; Li, N.; Liu, Y.; et al. Toll-Like Receptor 4 Promotes NO Synthesis by Upregulating GCHI Expression under Oxidative Stress Conditions in Sheep Monocytes/Macrophages. Oxidative Med. Cell. Longev. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vilar, A.; De Lemos, L.; Patraca, I.; Martínez, N.; Folch, J.; Junyent, F.; Verdaguer, E.; Pallàs, M.; Auladell, C.; Camins, A. Melatonin suppresses nitric oxide production in glial cultures by pro-inflammatory cytokines through p38 MAPK inhibition. Free. Radic. Res. 2014, 48, 119–128. [Google Scholar] [CrossRef]
- Kang, J.-W.; Lee, S.-M. Melatonin inhibits type 1 interferon signaling of toll-like receptor 4 via heme oxygenase-1 induction in hepatic ischemia/reperfusion. J. Pineal Res. 2012, 53, 67–76. [Google Scholar] [CrossRef]
- Jang, H.J.; Kim, Y.M.; Tsoyi, K.; Park, E.J.; Lee, Y.S.; Kim, H.J.; Lee, J.H.; Joe, Y.; Chung, H.T.; Chang, K.C. Ethyl Pyruvate Induces Heme Oxygenase-1 Through p38 Mitogen-Activated Protein Kinase Activation by Depletion of Glutathione in RAW 264.7 Cells and Improves Survival in Septic Animals. Antioxid. Redox Signal. 2012, 17, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.C.; Ko, Y.S.; Kim, H.J.; Nam, D.-Y.; Lee, D.-U. 13-Methylberberine reduces HMGB1 release in LPS-activated RAW264.7 cells and increases the survival of septic mice through AMPK/P38 MAPK activation. Int. Immunopharmacol. 2016, 40, 269–276. [Google Scholar] [CrossRef]
- Potter, S.J.; DeFalco, T. Role of the testis interstitial compartment in spermatogonial stem cell function. Reproduction 2017, 153, R151–R162. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.-L.; Zhang, B.-L.; Reiter, R.J.; Liu, Y.-X. Melatonin Ameliorates Inflammation and Oxidative Stress by Suppressing the p38MAPK Signaling Pathway in LPS-Induced Sheep Orchitis. Antioxidants 2020, 9, 1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9121277
Deng S-L, Zhang B-L, Reiter RJ, Liu Y-X. Melatonin Ameliorates Inflammation and Oxidative Stress by Suppressing the p38MAPK Signaling Pathway in LPS-Induced Sheep Orchitis. Antioxidants. 2020; 9(12):1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9121277
Chicago/Turabian StyleDeng, Shou-Long, Bao-Lu Zhang, Russel J. Reiter, and Yi-Xun Liu. 2020. "Melatonin Ameliorates Inflammation and Oxidative Stress by Suppressing the p38MAPK Signaling Pathway in LPS-Induced Sheep Orchitis" Antioxidants 9, no. 12: 1277. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9121277