Effects of Greenshell Mussel (Perna canaliculus) Intake on Pathological Markers of Multiple Phenotypes of Osteoarthritis in Rats

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenshell Mussel (GSM) Powder
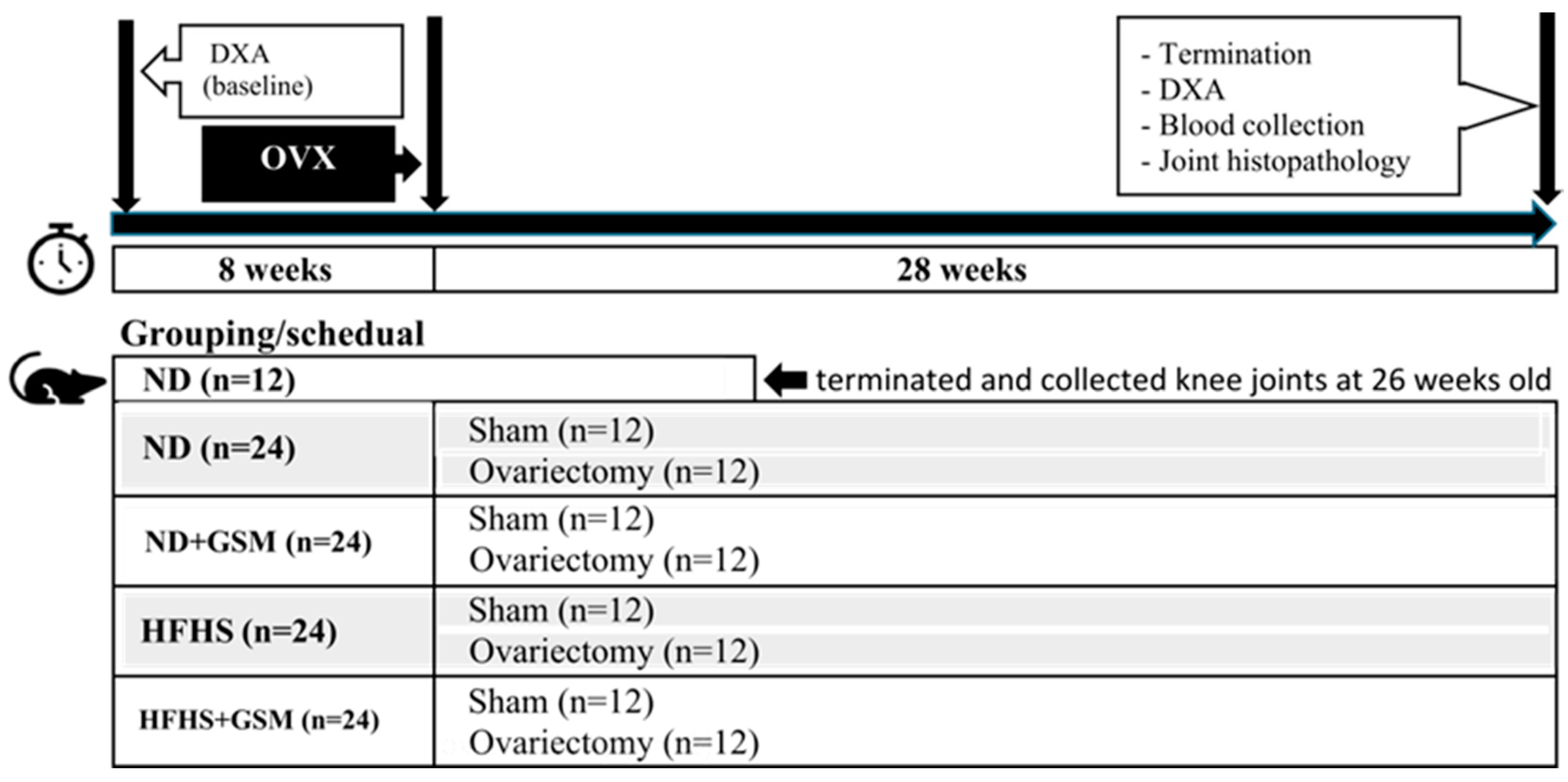
2.2. Animal Study
2.3. Dual-Energy X-ray Absorptiometry (DXA) Scans
2.4. Plasma Analysis
2.5. Leptin Gene Expression
2.6. Histopathological Examination of Knee Joints
2.7. Statistical Analysis
3. Results
3.1. Changes in Body Composition
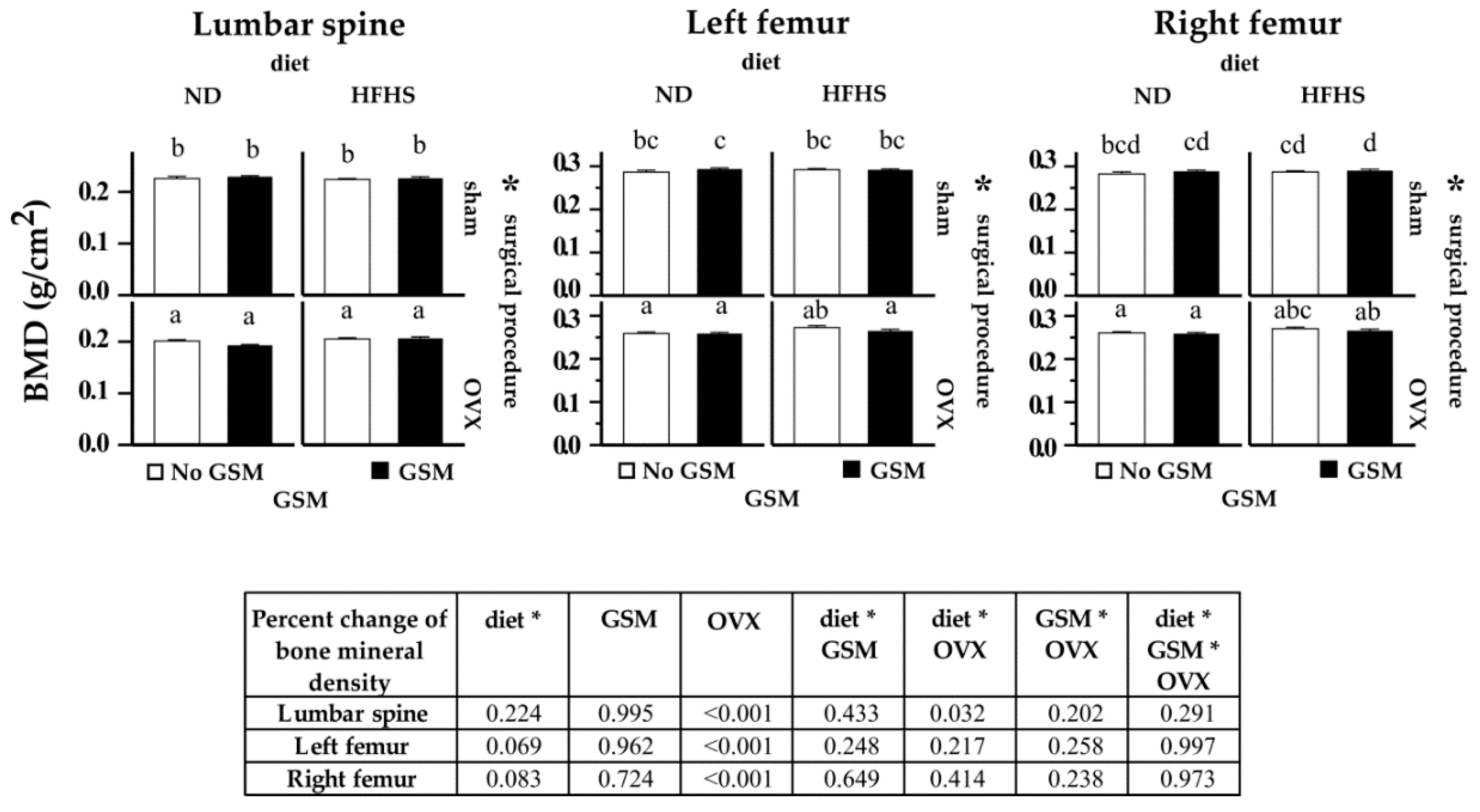
3.2. Bone Mineral Density (BMD) of Lumbar Spine and Femur
3.3. Analysis of Inflammatory Markers in Systemic Circulation
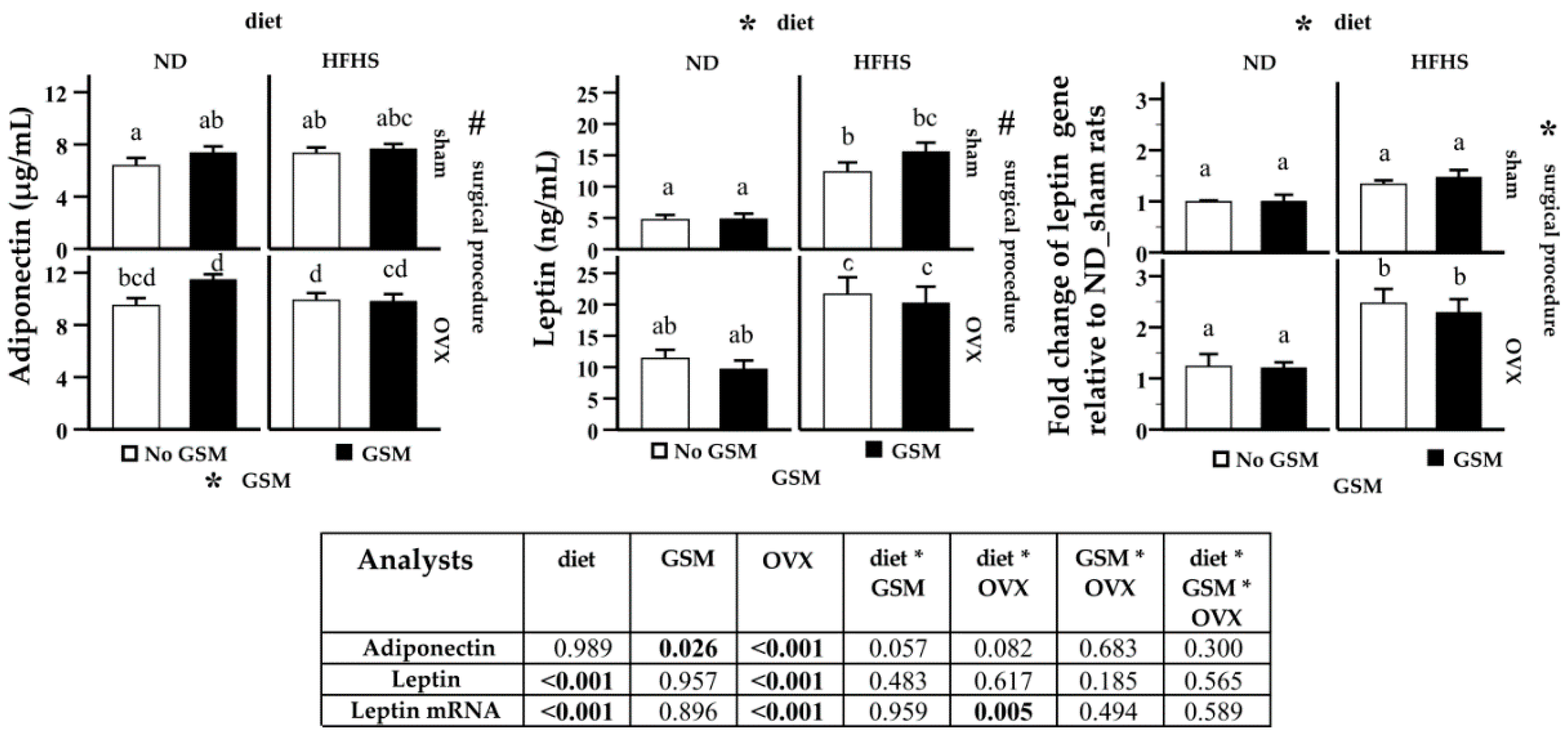
3.4. Analysis of Adipokines in Systemic Circulation and Fat Tissue
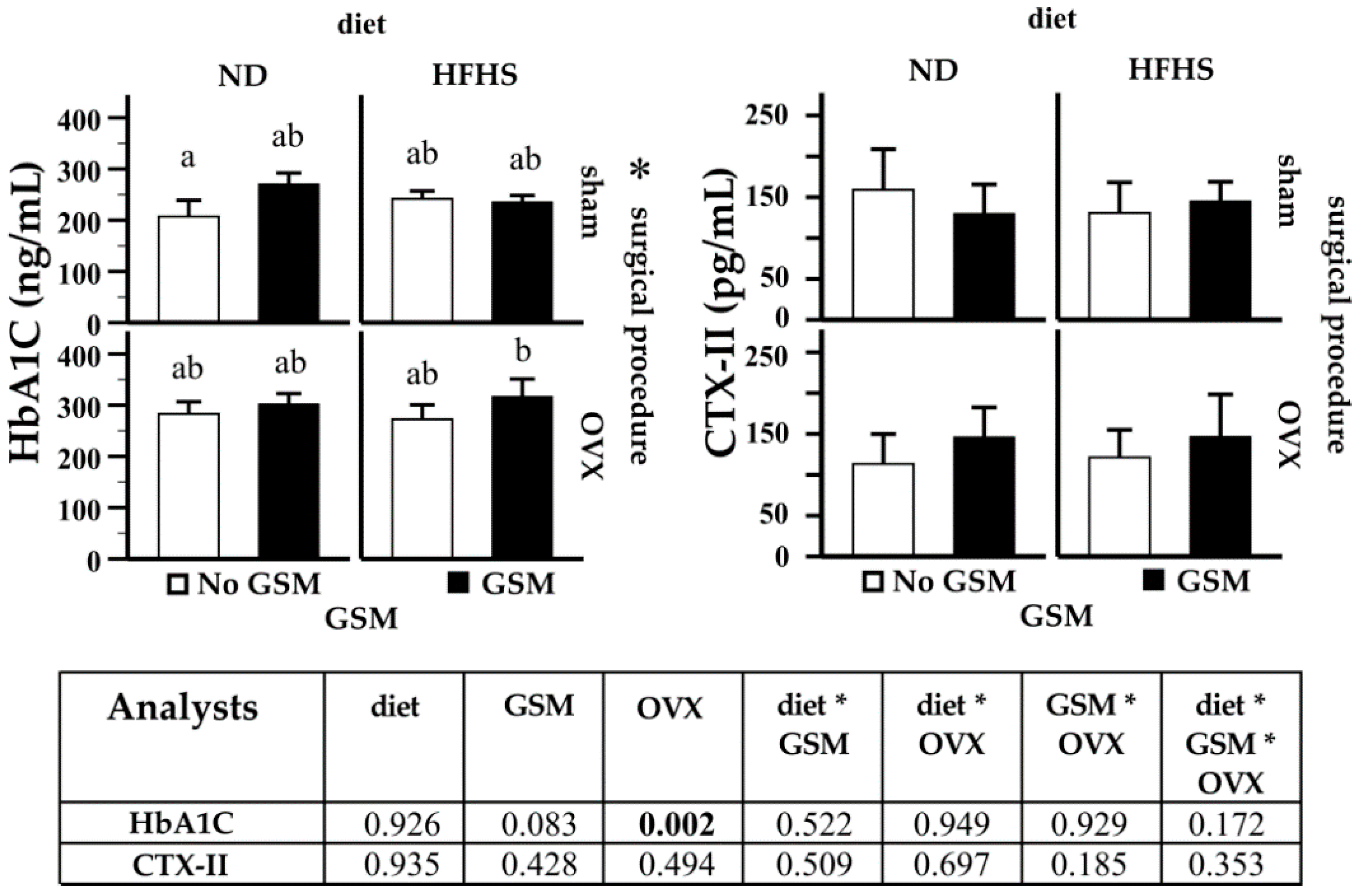
3.5. Analysis of Cartilage Degradation and Glucose Metabolism
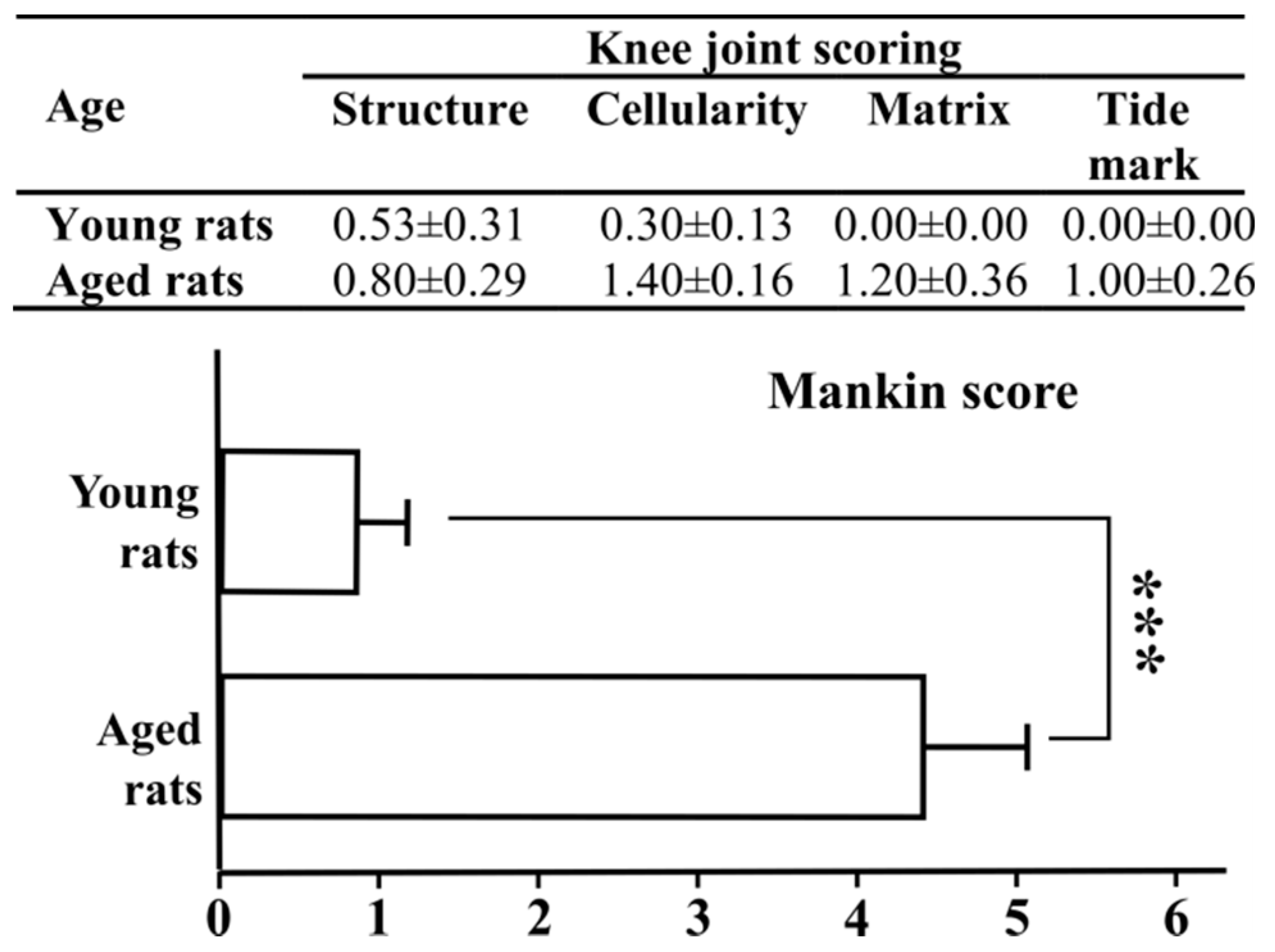
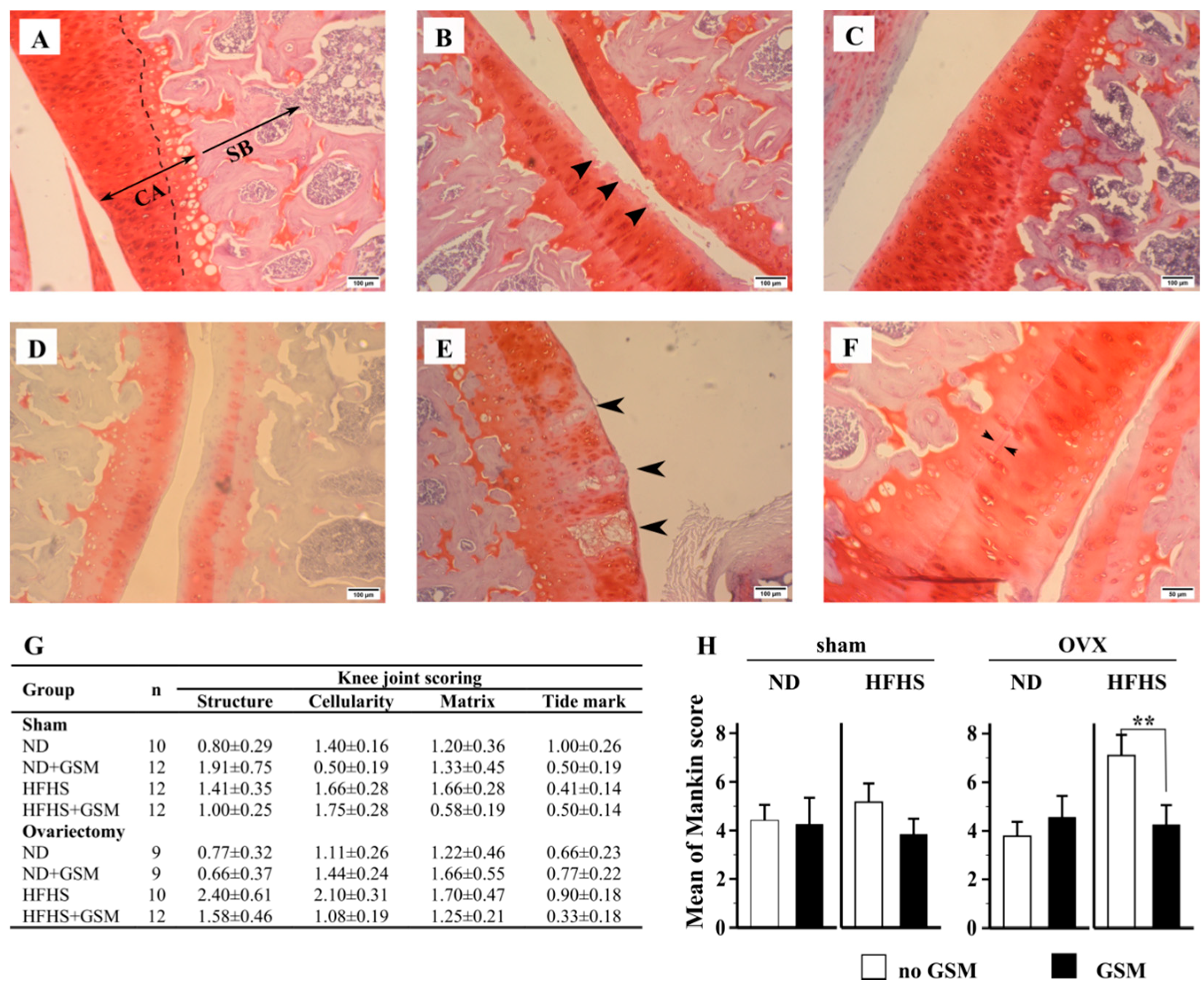
3.6. Pathological Changes in Knee Joints
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sham | OVX | ||||||||||||||
| ND | ND + GSM | HFHS | HFHS + GSM | ND | ND + GSM | HFHS | HFHS + GSM | p Values | |||||||
| BW 12th week | 273.73 ± 8.53 | 276.06 ± 6.20 | 280.84 ± 7.01 | 281.62 ± 7.54 | 275.64 ± 8.20 | 267.37 ± 8.96 | 276.51 ± 7.93 | 280.70 ± 6.41 | 0.911 | ||||||
| BW the end | 403.89 ± 17.30 a | 428.18 ± 18.05 a | 535.38 ± 27.16 bc | 578.15 ± 21.78 bc | 499.20 ± 18.56 ab | 488.27 ± 22.36 ab | 633.35 ± 34.01 c | 619.05 ± 29.34 c | <0.001 | ||||||
| Change of BW (%) | 49.01 ± 4.74 a | 54.55 ± 3.97 ab | 89.68 ± 6.97 cd | 105.51 ± 6.86 cde | 80.99 ± 3.34 bc | 83.41 ± 7.66 bc | 129.03 ± 10.51 e | 118.15 ± 7.44 de | <0.001 | ||||||
| Change of %body fat (%) | 185.55 ± 25.75 a | 221.94 ± 35.14 ab | 344.14 ± 33.06 bc | 377.30 ± 41.02 c | 328.88 ± 43.10 abc | 303.18 ± 18.48 abc | 357.20 ± 27.90 bc | 338.88 ± 37.36 bc | 0.001 | ||||||
| Statistical analysis by two-way ANOVA with three effects | |||||||||||||||
| diet | GSM | OVX | diet * GSM | diet * OVX | GSM * OVX | diet * GSM * OVX | |||||||||
| BW 12th week | 0.217 | 0.964 | 0.579 | 0.615 | 0.944 | 0.740 | 0.518 | ||||||||
| BW the end | <0.001 | 0.538 | <0.001 | 0.824 | 0.807 | 0.176 | 0.748 | ||||||||
| Change of BW (%) | <0.001 | 0.498 | <0.001 | 0.874 | 0.642 | 0.119 | 0.217 | ||||||||
| Change of %body fat (%) | <0.001 | 0.792 | 0.042 | 0.966 | 0.011 | 0.243 | 0.913 | ||||||||
| Sham | OVX | ||||||||
| ND | ND + GSM | HFHS | HFHS + GSM | ND | ND + GSM | HFHS | HFHS + GSM | p Values | |
| IL-1β (pg/mL) | 8.68 ± 3.63 | 7.03 ± 3.91 | 5.93 ± 1.77 | 4.48 ± 1.44 | 4.23 ± 1.61 | 7.50 ± 2.00 | 6.57 ± 1.68 | 3.73 ± 0.88 | 0.800 |
| IL-6 in (pg/mL) | 32.96 ± 4.53 | 26.79 ± 5.04 | 28.91 ± 5.35 | 27.69 ± 4.60 | 36.96 ± 9.36 | 32.44 ± 3.87 | 27.88 ± 4.27 | 28.75 ± 4.05 | 0.900 |
| IL-10 in(pg/mL) | 0.28 ± 0.29 | 1.27 ± 1.04 | 0.34 ± 0.24 | 0.07 ± 0.05 | 0.44 ± 0.32 | 0.46 ± 0.22 | 0.87 ± 0.58 | 0.17 ± 0.17 | 0.470 |
| TNF-α(pg/mL) | 1.08 ± 0.18 | 1.88 ± 0.28 | 1.78 ± 0.28 | 2.09 ± 0.18 | 1.53 ± 0.31 | 2.04 ± 0.84 | 1.89 ± 0.17 | 1.88 ± 0.56 | 0.750 |
| MCP-1(pg/mL) | 340.65 ± 49.76 ab | 387.69 ± 53.60 ab | 320.93 ± 55.72 a | 376.20 ± 71.69 ab | 483.26 ± 81.78 ab | 559.44 ± 69.88 ab | 574.41 ± 122.81 ab | 732.36 ± 163.99 b | 0.020 |
| PGE2 (pg/mL) | 1254.75 ± 187.65 | 1314.51 ± 234.97 | 1263.81 ± 212.19 | 1155.18 ± 237.85 | 1446.57 ± 221.96 | 1017.02 ± 123.24 | 1323.19 ± 313.98 | 1683.71 ± 321.77 | 0.660 |
| IFN-γ (pg/mL) | 241.55 ± 35.38 | 279.23 ± 40.09 | 222.36 ± 27.98 | 241.99 ± 35.16 | 263.20 ± 30.81 | 226.20 ± 33.19 | 217.63 ± 33.88 | 249.71 ± 26.38 | 0.695 |
| CXCL1 (pg/mL) | 23.65 ± 6.42 | 19.74 ± 5.36 | 16.83 ± 3.54 | 19.31 ± 4.88 | 32.09 ± 7.27 | 17.44 ± 4.71 | 19.71 ± 6.31 | 18.55 ± 4.82 | 0.283 |
| IL-18 (pg/mL) | 136.10 ± 48.45 | 159.41 ± 55.21 | 163.36 ± 76.23 | 112.23 ± 29.89 | 111.33 ± 13.87 | 119.65 ± 41.65 | 99.90 ± 16.12 | 130.33 ± 27.99 | 0.974 |
| IL-12 (pg/mL) | 40.30 ± 13.65 | 48.85 ± 11.20 | 34.60 ± 4.77 | 48.33 ± 5.69 | 51.25 ± 5.78 | 35.09 ± 0.76 | 65.66 ± 3.36 | 53.98 ± 5.35 | 0.804 |
| IL-17 (pg/mL) | 43.72 ± 8.31 | 50.58 ± 10.12 | 33.37 ± 6.78 | 43.40 ± 7.56 | 53.26 ± 4.60 | 33.08 ± 5.47 | 41.89 ± 10.90 | 34.59 ± 4.41 | 0.428 |
| IL-33 (pg/mL) | 612.27 ± 68.81 | 598.94 ± 95.12 | 622.62 ± 111.93 | 516.23 ± 60.95 | 650.36 ± 42.59 | 577.06 ± 80.56 | 584.99 ± 46.44 | 617.76 ± 58.09 | 0.850 |
| Statistical Analysis by Two-Way ANOVA with Three Effects | |||||||||
| diet | GSM | OVX | diet * GSM | diet * OVX | GSM * OVX | diet * GSM * OVX | |||
| IL-1β (pg/mL) | 0.316 | 0.689 | 0.540 | 0.379 | 0.563 | 0.597 | 0.347 | ||
| IL-6 in (pg/mL) | 0.297 | 0.468 | 0.525 | 0.497 | 0.528 | 0.805 | 0.977 | ||
| IL-10 in (pg/mL) | 0.375 | 0.970 | 0.995 | 0.085 | 0.256 | 0.220 | 0.646 | ||
| TNF-α (pg/mL) | 0.338 | 0.171 | 0.661 | 0.390 | 0.542 | 0.607 | 0.982 | ||
| MCP-1 (pg/mL) | 0.367 | 0.193 | 0.001 | 0.727 | 0.253 | 0.609 | 0.775 | ||
| PGE2 (pg/mL) | 0.565 | 0.863 | 0.480 | 0.363 | 0.311 | 0.976 | 0.163 | ||
| IFN-γ (pg/mL) | 0.400 | 0.573 | 0.760 | 0.583 | 0.712 | 0.504 | 0.350 | ||
| CXCL1 (pg/mL) | 0.233 | 0.267 | 0.593 | 0.200 | 0.794 | 0.354 | 0.646 | ||
| IL-18 (pg/mL) | 0.858 | 0.925 | 0.343 | 0.651 | 0.868 | 0.565 | 0.405 | ||
| IL-12 (pg/mL) | 0.436 | 0.871 | 0.333 | 0.778 | 0.262 | 0.161 | 0.984 | ||
| IL-17 (pg/mL) | 0.189 | 0.609 | 0.690 | 0.438 | 0.711 | 0.036 | 0.639 | ||
| IL-33 (pg/mL) | 0.659 | 0.468 | 0.719 | 0.951 | 0.828 | 0.719 | 0.370 | ||
References
- Briggs, A.M.; Cross, M.J.; Hoy, D.G.; Sanchez-Riera, L.; Blyth, F.M.; Woolf, A.D.; March, L. Musculoskeletal health conditions represent a global threat to healthy aging: A report for the 2015 world health organization world report on ageing and health. Gerontologist 2016, 56, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Cicuttini, F.M.; Baker, J.R.; Spector, T.D. The association of obesity with osteoarthritis of the hand and knee in women: A twin study. J. Rheumatol. 1996, 23, 1221–1226. [Google Scholar]
- Woolf, A.D.; Pfleger, B. Burden of major musculoskeletal conditions. Bull. World Health Organ 2003, 81, 646–656. [Google Scholar] [PubMed]
- Bijlsma, J.W.; Berenbaum, F.; Lafeber, F.P. Osteoarthritis: An update with relevance for clinical practice. Lancet 2011, 377, 2115–2126. [Google Scholar] [CrossRef]
- Karuppal, R. Current concepts in the articular cartilage repair and regeneration. J. Orthop. 2017, 14, A1–A3. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yin, J.; Gao, J.; Cheng, T.S.; Pavlos, N.J.; Zhang, C.; Zheng, M.H. Subchondral bone in osteoarthritis: Insight into risk factors and microstructural changes. Arthrits Res. Ther. 2013, 15. [Google Scholar] [CrossRef] [Green Version]
- Suri, S.; Walsh, D.A. Original full length article: Osteochondral alterations in osteoarthritis. Bone 2012, 51, 204–211. [Google Scholar] [CrossRef]
- Jimenez, P.A.; Glasson, S.S.; Trubetskoy, O.V. Spontaneous osteoarthritis in Dunkin Hartley guinea pigs: Histologic, radiologic, and biochemical changes. Lab. Anim. Sci. 1997, 47, 598–601. [Google Scholar]
- Poulet, B.; Westerhof, T.A.; Hamilton, R.W. Spontaneous osteoarthritis in Str/ort mice is unlikely due to greater vulnerability to mechanical trauma. Osteoarthr. Cartil. 2013, 21, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Kuyinu, E.L.; Narayanan, G.; Nair, L.S.; Laurencin, C.T. Animal models of osteoarthritis: Classification, update, and measurement of outcomes. J. Ortho. Sur. Res. 2016, 11, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, A. Obesity, metabolic syndrome, and type 2 diabetes: Inflammatory basis of glucose metabolic disorders. Nutr. Rev. 2007, 65, S152–S156. [Google Scholar] [CrossRef] [PubMed]
- Teichtahl, A.J.; Wluka, A.E.; Proietto, J.; Cicuttini, F.M. Obesity and the female sex, risk factors for knee osteoarthritis that may be attributable to systemic or local leptin biosynthesis and its cellular effects. Med. Hypotheses 2005, 65, 312–315. [Google Scholar] [CrossRef] [PubMed]
- de Visser, H.M.; Kozijn, A.E.; Pouran, B.; van Rijen, M.H.; Weinans, H.; Mastbergen, S.C.; Coeleveld, K.; Lafeber, F.P.J.G. Metabolic dysregulation accelerates injury-induced joint degeneration, driven by local inflammation; an in vivo rat study. J. Orthop. Res. 2018, 36, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmi, G.; Faust, R. Suitability of the C57 black mouse as an experimental animal for the study of skeletal changes due to ageing, with special reference to osteo-arthrosis and its response to tribenoside. Pharmacology 1976, 14, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.H.; Reimer, R.A.; Seerattan, R.A.; Leonard, T.R.; Herzog, W. Using diet-induced obesity to understand a metabolic subtype of osteoarthritis in rats. Osteoarthr. Cartil. 2015, 23, 957–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.H.; Paul, H.A.; Reimer, R.A.; Seerattan, R.A.; Hart, D.A.; Herzog, W. Relationship between inflammation, the gut microbiota, and metabolic osteoarthritis development: Studies in a rat model. Osteoarthr. Cartil. 2015, 23, 1989–1998. [Google Scholar] [CrossRef] [Green Version]
- Sun, A.R.; Panchal, S.K.; Friis, T.; Sekar, S.; Crawford, R.; Brown, L.; Xiao, Y.; Prasadam, I. Obesity-associated metabolic syndrome spontaneously induces infiltration of pro-inflammatory macrophage in synovium and promotes osteoarthritis. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.L.; Blanchet, T.J.; Peluso, D.; Hopkins, B.; Morris, E.A.; Glasson, S.S. Osteoarthritis severity is sex dependent in a surgical mouse model. Osteoarthr. Cartil. 2007, 15, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Sekar, S.; Shafie, S.R.; Prasadam, I.; Crawford, R.; Panchal, S.K.; Brown, L.; Xiao, Y. Saturated fatty acids induce development of both metabolic syndrome and osteoarthritis in rats. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Treschow, A.P.; Hodges, L.D.; Kalafatis, N.; Macrides, T.A.; Wright, P.F.A.; Wynne, P.M. Novel anti-inflammatory ω-3 PUFAs from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. Part B 2007, 147, 645–656. [Google Scholar] [CrossRef]
- Miller, M.R.; Pearce, L.; Bettjeman, B.I. Detailed distribution of lipids in greenshell™ mussel (Perna canaliculus). Nutrients 2014, 6, 1454–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (Lyprinol) from the NZ green-lipped mussel. Inflammopharmacology 1997, 5, 237–246. [Google Scholar] [CrossRef] [PubMed]
- McPhee, S.; Hodges, L.D.; Wright, P.F.A.; Wynne, P.M.; Kalafatis, N.; Harney, D.W.; Macrides, T.A. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. Part B 2007, 146, 346–356. [Google Scholar] [CrossRef]
- Singh, M.; Hodges, L.D.; Wright, P.F.A.; Cheah, D.M.Y.; Wynne, P.M.; Kalafatis, N.; Macrides, T.A. The CO2-SFE crude lipid extract and the free fatty acid extract from Perna canaliculus have anti-inflammatory effects on adjuvant-induced arthritis in rats. Comp. Biochem. Physiol. Part B 2008, 149, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Lu, W.W.; Chiu, K.Y. Importance of subchondral bone in the pathogenesis and management of osteoarthritis from bench to bed. J. Orthop. Trans. 2014, 2, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Shen, J.; Zhao, W.; Wang, T.; Han, L.; Hamilton, J.L.; Im, H.J. Osteoarthritis: Toward a comprehensive understanding of pathological mechanism. Bone Res. 2017, 5, 16044. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Q.; Yang, W.; Chen, J.; Wang, Y. Metabolic syndrome meets osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 729–737. [Google Scholar] [CrossRef]
- Kruger, M.C.; Morel, P.C.H. Experimental control for the ovariectomized rat model: Use of sham versus nonmanipulated animal. J. Appl. Anim. Welf. Sci. 2016, 19, 73–80. [Google Scholar] [CrossRef]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J. Bone Jt. Surg. Am. 1971, 53, 523–537. [Google Scholar] [CrossRef]
- Heidemann, C.; Schulze, M.B.; Franco, O.H. Dietary patterns and risk of mortality from cardiovascular disease, cancer, and all causes in a prospective cohort of women. Circulation 2008, 118, 230–237. [Google Scholar] [CrossRef]
- Siriarchavatana, P.; Kruger, M.C.; Miller, M.R.; Tian, H.S.; Wolber, F.M. The preventive effects of greenshell mussel (Perna canaliculus) on early-stage metabolic osteoarthritis in rats with diet-induced obesity. Nutrients 2019, 11, 1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, R.; Ross, R.P.; Stanton, C.; Fitzgerald, G.F. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Knott, L.; Avery, N.C.; Hollander, A.P.; Tarlton, J.F. Regulation of osteoarthritis by omega-3 (n-3) polyunsaturated fatty acids in a naturally occurring model of disease. Osteoarthr. Cartil. 2011, 19, 1150–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, A.; Hutchison, E.; Hudson, J.; Kawashima, Y.; Komori, N.; Singh, A.; Brush, R.S.; Anderson, R.E.; Sonntag, W.E.; Matsumoto, H.; et al. Metabolic enrichment of omega-3 polyunsaturated fatty acids does not reduce the onset of idiopathic knee osteoarthritis in mice. Osteoarthr. Cartil. 2014, 22, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaei, E.; Mowla, K.; Ghorbani, A.; Bahadoram, S.; Bahadoram, M.; Dargahi-Malamir, M. The effect of Omega-3 fatty acids in patients with active rheumatoid arthritis receiving DMARDs therapy: Double-blind randomized controlled trial. Glob. J. Health Sci. 2015, 8, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layé, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-inflammatory effects of omega-3 fatty acids in the brain: Physiological mechanisms and relevance to pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef]
- Saltzman, E.T.; Thomsen, M.; Hall, S.; Vitetta, L. Perna canaliculus and the Intestinal Microbiome. Mar. Drugs 2017, 15, 207. [Google Scholar] [CrossRef]
- Miller, M.; Perry, N.; Burgess, E.; Marshall, S. Regiospecific analyses of triacylglycerols of hoki (Macruronus novaezelandiae) and Greenshell™ Mussel (Perna canaliculus). J. Am. Oil. Chem. Soc. 2011, 88, 509–516. [Google Scholar] [CrossRef]
- Haag, M.; Magada, O.N.; Claassen, N.; Böhmer, L.H.; Kruger, M.C. Omega-3 fatty acids modulate ATPases involved in duodenal Ca absorption. Prostaglandins Leukot Essent Fat. Acids 2003, 68, 423–429. [Google Scholar] [CrossRef]
- Pranprawit, A.; Wolber, F.M.; Heyes, J.A.; Molan, A.L.; Kruger, M.C. Short-term and long-term effects of excessive consumption of saturated fats and/or sucrose on metabolic variables in Sprague Dawley rats: A pilot study. J. Sci. Food Agric. 2013, 93, 3191–3197. [Google Scholar] [CrossRef]
- Ràfols, M.E. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Azamar-Llamas, D.; Hernandez-Molina, G.; Ramos-Avalos, B.; Furuzawa-Carballeda, J. Adipokine contribution to the pathogenesis of osteoarthritis. Mediat. Inflamm. 2017, 2017, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Rui, C.; Hua, W.; Fengxia, L. Mechanisms linking inflammation to insulin resistance. Int. J. Endocrinol. 2015. [Google Scholar] [CrossRef]
- Lizcano, F.; Guzmán, G. Estrogen deficiency and the origin of obesity during menopause. BioMed. Res. Int. 2014, 2014, 757461. [Google Scholar] [CrossRef] [PubMed]
- Villafuerte, B.C.; Fine, J.B.; Bai, Y.; Zhao, W.; Fleming, S.; DiGirolamo, M. Expressions of leptin and insulin-like growth factor-i are highly correlated and region-specific in adipose tissue of growing rats. Obes. Res. 2000, 8, 646–655. [Google Scholar] [CrossRef]
- Mulet, T.; Picó, C.; Oliver, P.; Palou, A. Blood leptin homeostasis: Sex-associated differences in circulating leptin levels in rats are independent of tissue leptin expression. Int. J. Biochem. Cell Biol. 2003, 35, 104–110. [Google Scholar] [CrossRef]
- Inadera, H. The usefulness of circulating adipokine levels for the assessment of obesity-related health problems. Int. J. Med. Sci. 2008, 5, 248–262. [Google Scholar] [CrossRef] [Green Version]
- Stofkova, A. Leptin and adiponectin: From energy and metabolic dysbalance to inflammation and autoimmunity. Endocr. Regul. 2009, 43, 157–168. [Google Scholar]
- Lago, R.; Gomez, R.; Otero, M.; Lago, F.; Gallego, R.; Dieguez, C.; Gomez-Reino, J.J.; Gualillo, O. A new player in cartilage homeostasis: Adiponectin induces nitric oxide synthase type II and pro-inflammatory cytokines in chondrocytes. Osteoarthr. Cartil. 2008, 16, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- BrahmaNaidu, P.; Nemani, H.; Meriga, B.; Mehar, S.K.; Potana, S.; Ramgopalrao, S. Mitigating efficacy of piperine in the physiological derangements of high fat diet induced obesity in Sprague Dawley rats. Chem. Biol. Interact. 2014, 221, 42–51. [Google Scholar] [CrossRef]
- Ilich, J.Z.; Kelly, O.J.; Kim, Y.; Spicer, M.T. Low-grade chronic inflammation perpetuated by modern diet as a promoter of obesity and osteoporosis. Arch. Ind. Hyg. Toxicol. Arh. Hig. Rada I Toksikol. 2014, 65, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagiolo, U.; Cossarizza, A.; Scala, E.; Fanales-Belasio, E.; Ortolani, C.; Cozzi, E.; Monti, D.; Franceschi, C.; Paganelli, R. Increased cytokine production in mononuclear cells of healthy elderly people. Euro. J. Immunol. 1993, 23, 2375–2378. [Google Scholar] [CrossRef] [PubMed]
- Culemann, S.; Grüneboom, A.; Nicolás-Ávila, J.Á.; Weidner, D.; Lämmle, K.F.; Rothe, T.; Quintana, J.A.; Kirchner, P.; Krljanac, B.; Eberhardt, M.; et al. Locally renewing resident synovial macrophages provide a protective barrier for the joint. Nature 2019, 572, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Nio, Y.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Funata, M.; Yamaguchi, M.; Ueki, K.; Kadowaki, T. Monocyte chemoattractant protein-1 (MCP-1) deficiency enhances alternatively activated M2 macrophages and ameliorates insulin resistance and fatty liver in lipoatrophic diabetic A-ZIP transgenic mice. Diabetol. Croat. 2012, 55, 3350–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, H.; Tateya, S.; Tamori, Y. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Kouyama, K.; Miyake, K.; Zenibayashi, M.; Hirota, Y. Association of serum MCP-1 concentration and MCP-1 polymorphism with insulin resistance in Japanese individuals with obese type 2 diabetes. Kobe J. Med. Sci. 2007, 53, 345–354. [Google Scholar]
- Zenobia, C.; Hajishengallis, G. Basic biology and role of interleukin-17 in immunity and inflammation. Periodontol 2000 2015, 69, 142–159. [Google Scholar] [CrossRef]
- Goldring, S.R.; Goldring, M.B. Changes in the osteochondral unit during osteoarthritis: Structure, function and cartilage-bone crosstalk. Nat. Rev. Rheumatol. 2016, 12, 632–644. [Google Scholar] [CrossRef]
- Cook, J.L.; Hung, C.T.; Kuroki, K.; Stoker, A.M.; Cook, C.R.; Pfeiffer, F.M.; Sherman, S.L.; Stannard, J.P. Animal models of cartilage repair. Bone Joint Res. 2014, 3, 89–94. [Google Scholar] [CrossRef]
- McCoy, A. Animal models of osteoarthritis: Comparisons and key considerations. Vet. Pathol. 2015, 52, 803–818. [Google Scholar] [CrossRef]
- Vogel, H.G.; Vogel, W.H. Drug Discovery and Evaluation: Pharmacological Assays; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar]
- Griffin, T.M.; Guilak, F. Why is obesity associated with osteoarthritis? Insights from mouse models of obesity. Biorheology 2008, 45, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Brunner, A.M.; Henn, C.M.; Drewniak, E.I.; Lesieur-Brooks, A.; Machan, J.; Crisco, J.J.; Ehrlich, M.G. High dietary fat and the development of osteoarthritis in a rabbit model. Osteoarthr. Cartil. 2012, 20, 584–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrandiz, M.L.; Terencio, M.C.; Ruhi, R.; Verges, J.; Montell, E.; Torrent, A.; Alcaraz, M.J. Influence of age on osteoarthritis progression after anterior cruciate ligament transection in rats. Exp. Gerontol. 2014, 55, 44–48. [Google Scholar] [CrossRef]
- Oestergaard, S.; Sondergaard, B.C.; Hoegh-Andersen, P.; Henriksen, K.; Qvist, P.; Christiansen, C.; Tanko, L.B.; Karsdal, M.A. Effects of ovariectomy and estrogen therapy on type II collagen degradation and structural integrity of articular cartilage in rats—Implications of the time of initiation. Arthritis Rheum. 2006, 54, 2441–2451. [Google Scholar] [CrossRef] [PubMed]
- Høegh-Andersen, P.; Tankó, L.B.; Andersen, T.L.; Lundberg, C.V.; Mo, J.A.; Heegaard, A.-M.; Delaissé, J.-M.; Christgau, S. Ovariectomized rats as a model of postmenopausal osteoarthritis: Validation and application. Arthritis Res. Ther. 2004, 6, R169–R180. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siriarchavatana, P.; Kruger, M.C.; Miller, M.R.; Tian, H.; Wolber, F.M. Effects of Greenshell Mussel (Perna canaliculus) Intake on Pathological Markers of Multiple Phenotypes of Osteoarthritis in Rats. Appl. Sci. 2020, 10, 6131. https://0-doi-org.brum.beds.ac.uk/10.3390/app10176131
Siriarchavatana P, Kruger MC, Miller MR, Tian H, Wolber FM. Effects of Greenshell Mussel (Perna canaliculus) Intake on Pathological Markers of Multiple Phenotypes of Osteoarthritis in Rats. Applied Sciences. 2020; 10(17):6131. https://0-doi-org.brum.beds.ac.uk/10.3390/app10176131
Chicago/Turabian StyleSiriarchavatana, Parkpoom, Marlena C. Kruger, Matthew R. Miller, Hong (Sabrina) Tian, and Frances M. Wolber. 2020. "Effects of Greenshell Mussel (Perna canaliculus) Intake on Pathological Markers of Multiple Phenotypes of Osteoarthritis in Rats" Applied Sciences 10, no. 17: 6131. https://0-doi-org.brum.beds.ac.uk/10.3390/app10176131