Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds

 ,
,  ,
,  ,
,  ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
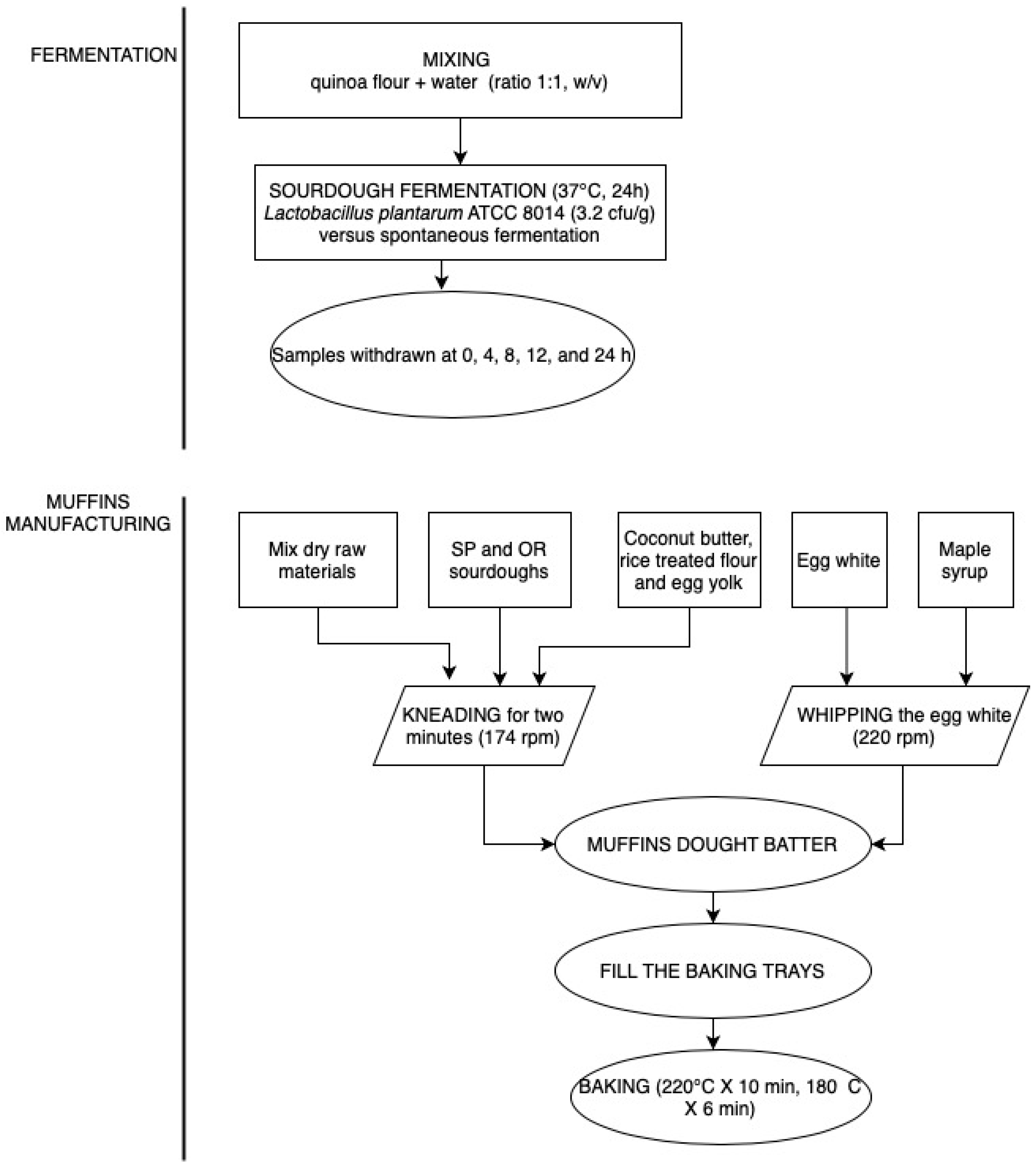
2.2. Microbial Starter Culture Preparation, Sourdough Preparation, and Muffins Formulation
2.3. Organic Acids and Glucose, Maltose, and Fructose Determination by HPLC-RID
2.4. Folic Acid Determination
2.5. Minerals Content
2.6. Total Flavonoids
2.7. Total Phenols Assay by Folin–Ciocalteau Reagent
2.8. Radical Scavenging Activity by DPPH Assay
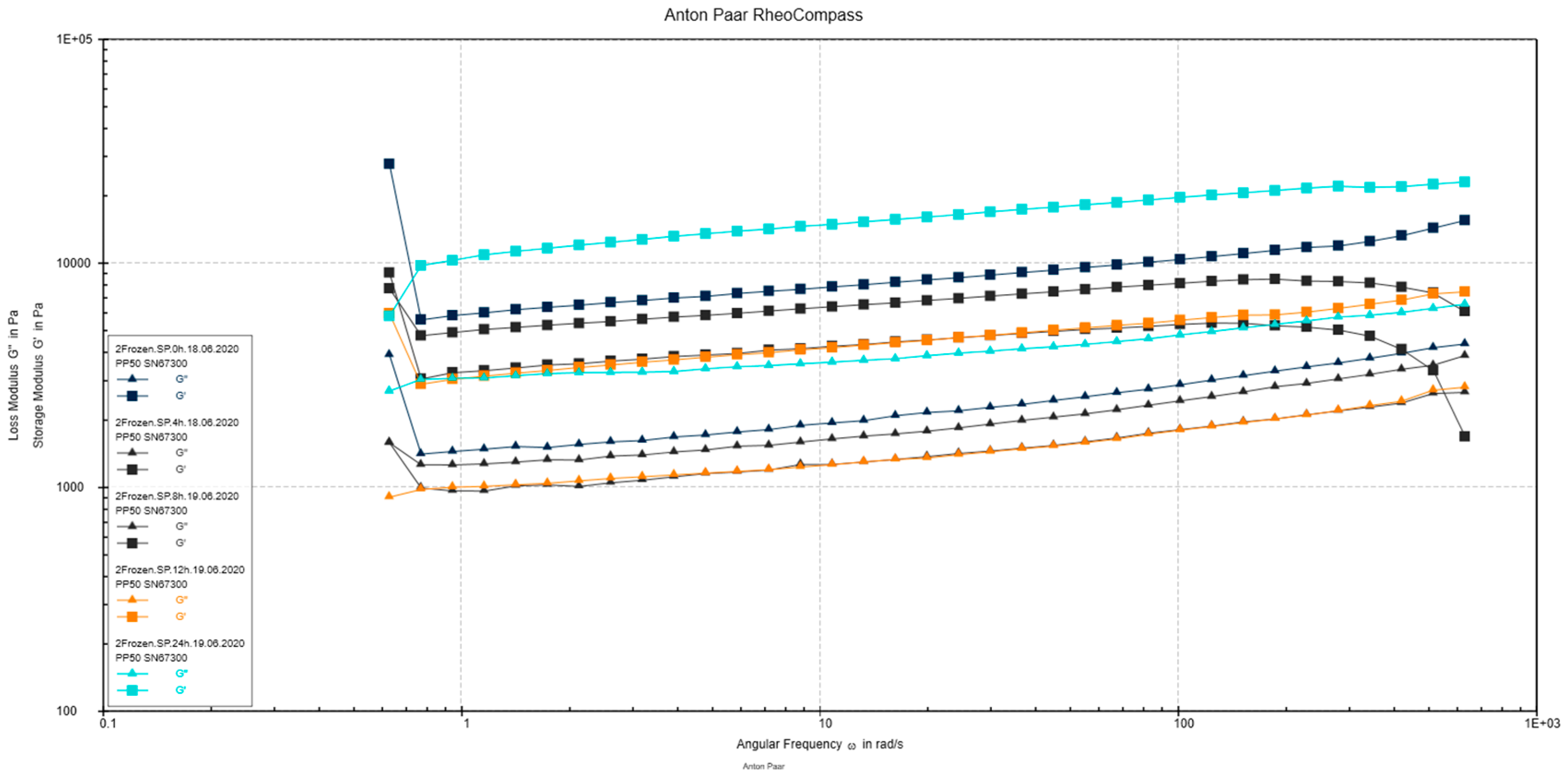
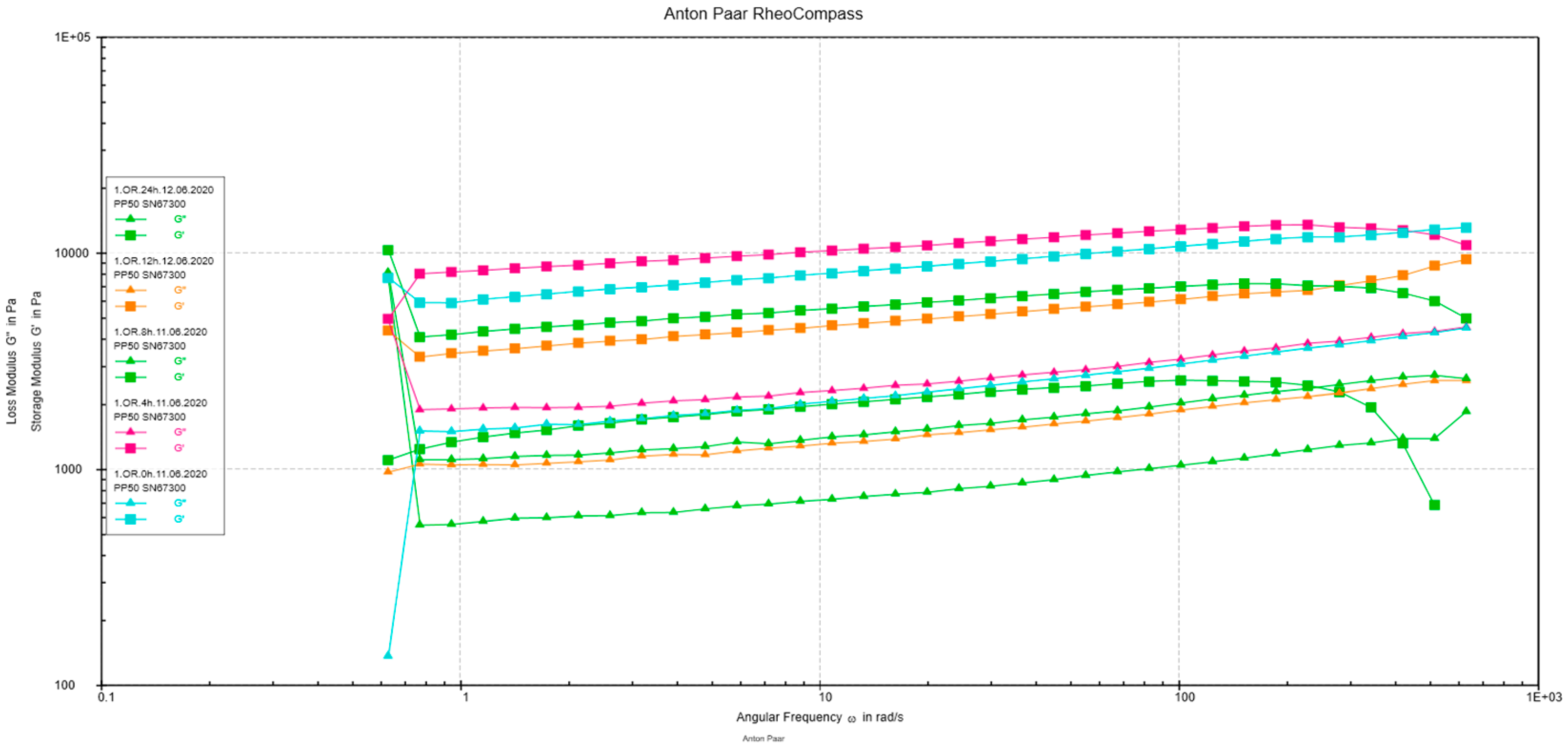
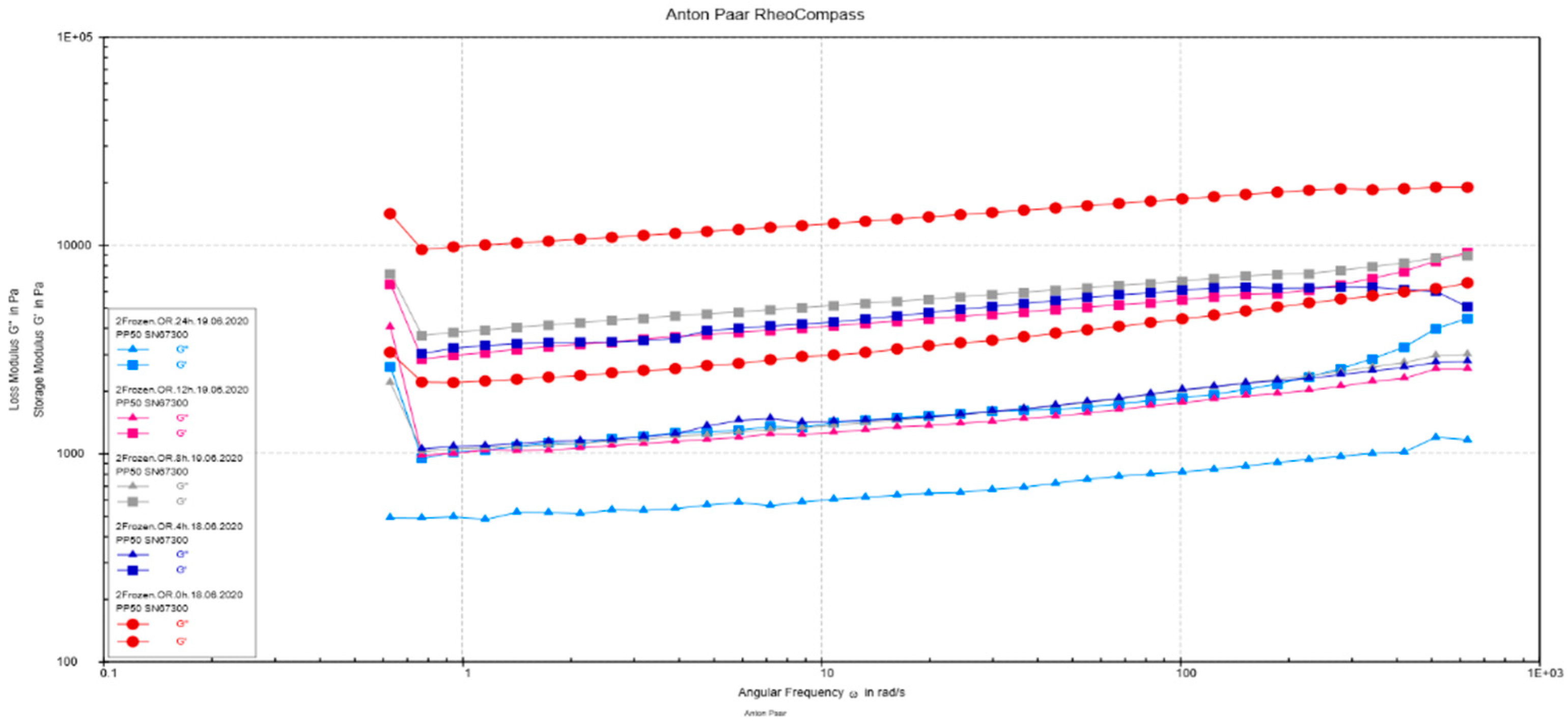
2.9. Rheological Measurements
2.10. Statistical Analysis
3. Results and Discussion
3.1. Carbohydrates, Organic Acids, Folic Acid, Minerals, Flavonoids, Total Phenols Content, and Radical Scavenging Activity of Quinoa Flour (QF)
3.1.1. Carbohydrates and Organic Acids from Quinoa Wholemeal Flour (QWF)
3.1.2. Quinoa Wholemeal Flour (QWF) Content in Folic Acid
3.1.3. QWF Minerals Content
3.1.4. QWF Flavonoids
3.2. Carbohydrates, Organic Acids, Folic Acid, Minerals, Flavonoids, Total Phenols Content, Radical Scavenging Activity, and Rheological Features from Quinoa Sourdoughs with Lactobacillus plantarum (Lp) ATCC 8014 (SP) and without Lactobacillus plantarum ATCC 8014 (OR)
3.2.1. OR, SP Carbohydrates, and Organic Acids Contents
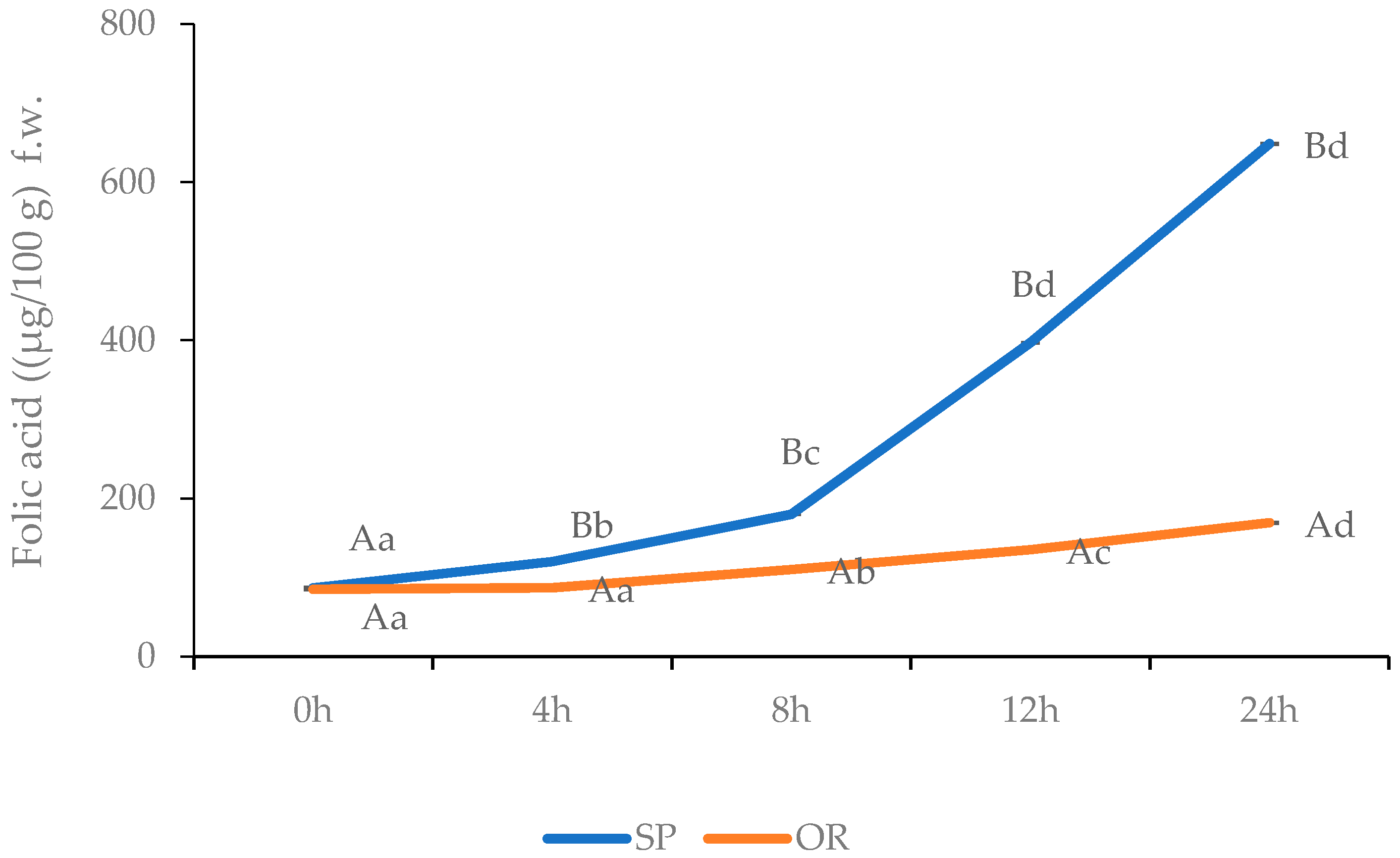
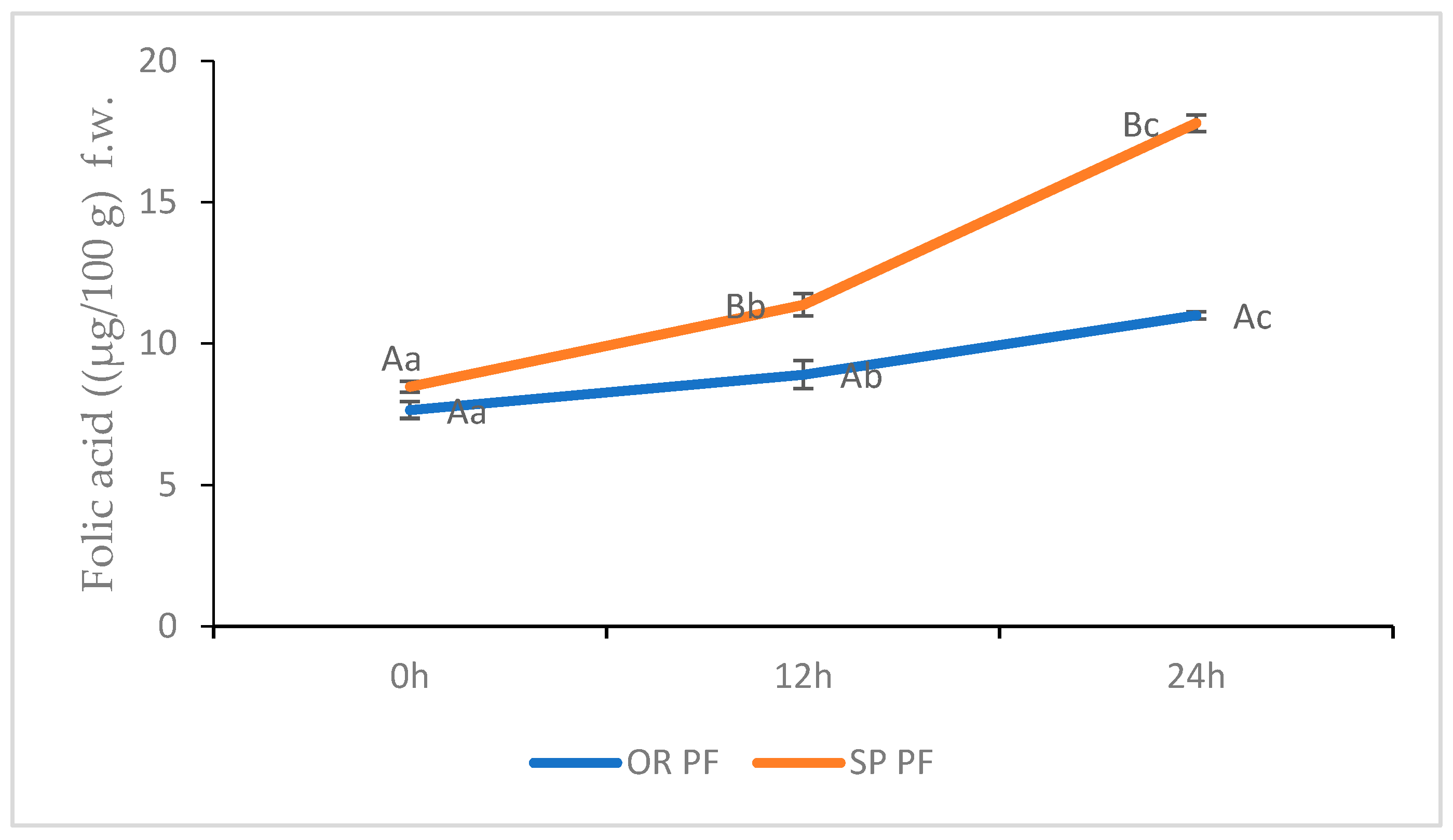
3.2.2. SP and OR folic Acid Content
3.2.3. QP and QQ Macro and Microelements Content
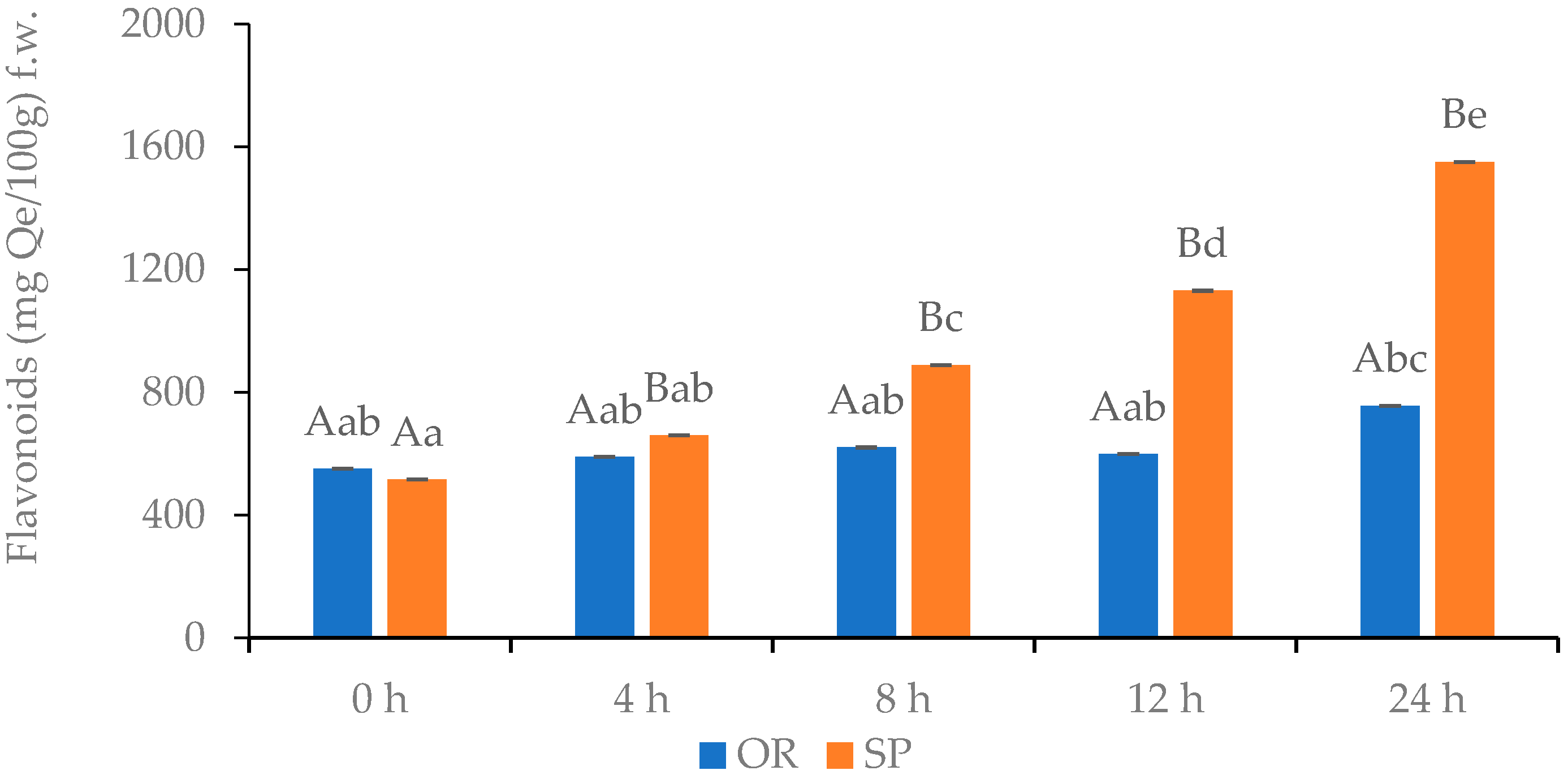
3.2.4. Total Flavonoids Content of OR and SP Sourdoughs
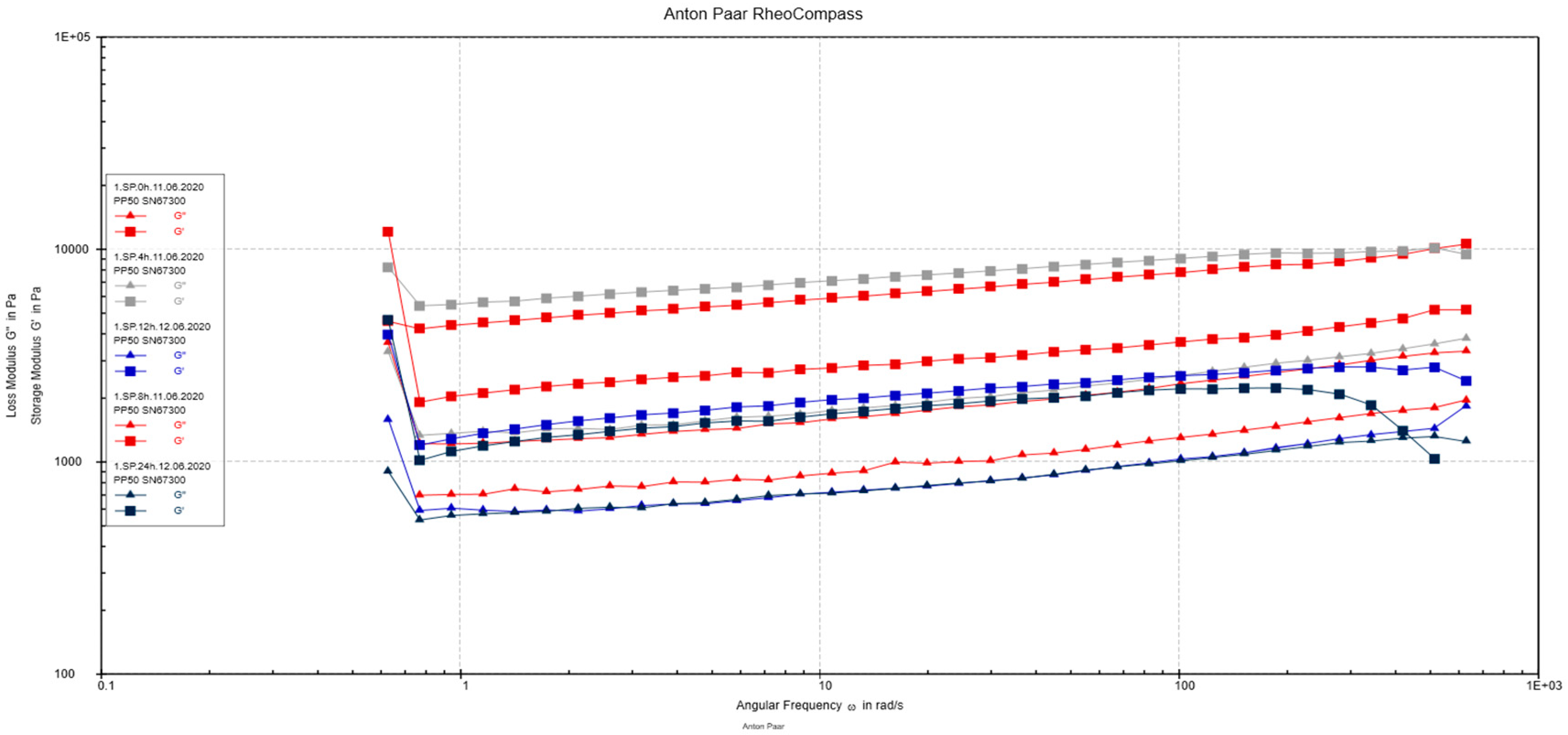
3.2.5. Rheological Measurements
3.3. Carbohydrates, Organic Acids, Folic Acid, Minerals, Flavonoids, Total Phenols Content, Radical Scavenging Activity, of OR PF and SP PF Muffins
3.3.1. Carbohydrates and organic acids content of gluten free muffins (GFM)
3.3.2. Folic Acid of Gluten Free Muffins (GFM)
3.3.3. Minerals Content of Gluten Free Muffins (GFM)
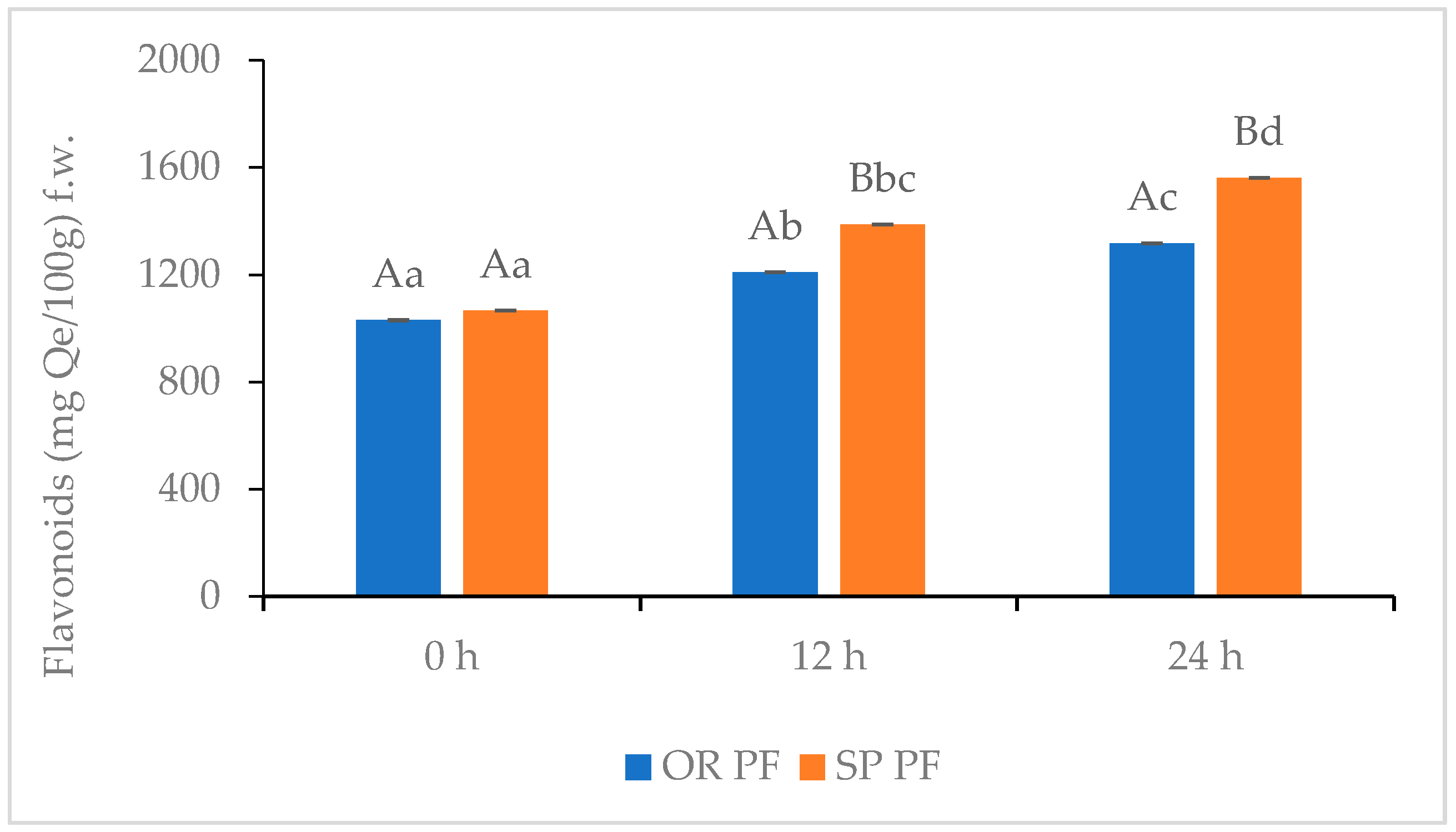
3.3.4. Flavonoids, Total Phenols Content, Radical Scavenging Activity of Gluten Free Muffins (GFM)
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kårlund, A.; Gómez-Gallego, C.; Korhonen, J.; Palo-Oja, O.M.; El-Nezami, H.; Kolehmainen, M. Harnessing microbes for sustainable development: Food fermentation as a tool for improving the nutritional quality of alternative protein sources. Nutrients 2020, 12, 1020. [Google Scholar] [CrossRef] [Green Version]
- Chiş, M.S.; Păucean, A.; Stan, L.; Suharoschi, R.; Socaci, S.A.; Man, S.M.; Pop, C.R.; Muste, S. Protein metabolic conversion of nutritional features during quinoa sourdough fermentation and its impact on baked goods. CyTA J. Food 2018, 280, 744–753. [Google Scholar]
- Cyrkot, M.; Anders, S.; Kamprath, C.; Liu, A.; Mileski, H.; Dowhaniuk, J.; Nasser, R.; Marcon, M.; Brill, H.; Turner, J.M.; et al. Folate content of gluten-free food purchases and dietary intake are low in children with coeliac disease. Int. J. Food Sci. Nutr. 2020, 7486, 1–13. [Google Scholar] [CrossRef] [PubMed]
- El-Sohaimy, S.A.; Shehata, M.G.; Mehany, T.; Zeitoun, M.A. Nutritional, physicochemical, and sensorial evaluation of flat bread supplemented with quinoa flour. Hindawi Int. J. Food Sci. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pappier, U.; Pinto, V.F.; Larumbe, G.; Vaamonde, G. Effect of processing for saponin removal on fungal contamination of quinoa seeds (Chenopodium quinoa Willd.). Int. J. Food Microbiol. 2008, 125, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Gonzalez, C.; Bolívar-Monsalve, J.; Ramírez-Toro, C.; Bolívar, G.A. Effect of lactic acid fermentation on quinoa dough to prepare gluten-free breads with high nutritional and sensory quality. J. Food Process. Preserv. 2017, 42, e13551. [Google Scholar] [CrossRef]
- Vega-Galvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martinez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Nascimento, A.C.; Mota, C.; Coelho, I.; Gueifao, S.; Santos, M.; Matos, A.S.; Gimenez, A.; Lobo, M.; Samman, N.; Castanheira, I. Characterisation of nutrient profile of quinoa (Chenopodium quinoa), amaranth (Amaranthus caudatus), and purple corn (Zea mays L.) consumed in the North of Argentina: Proximates, minerals and trace elements. Food Chem. 2014, 148, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco-Valencia, R.; Hellström, J.K.; Pihlava, J.M.; Mattila, P.H. Flavonoids and other phenolic compounds in Andean indigenous grains: Quinoa (Chenopodium quinoa), kañiwa (Chenopodium pallidicaule) and kiwicha (Amaranthus caudatus). Food Chem. 2010, 120, 128–133. [Google Scholar] [CrossRef]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Caboni, M.F. Simultaneous determination of phenolic compounds and saponins in quinoa (Chenopodium quinoa Willd.) by a liquid chromatography-diode array detection-electrospray ionization-time-of-flight mass spectrometry methodology. J. Agric. Food Chem. 2011, 59, 10815–10825. [Google Scholar] [CrossRef]
- Kumpun, S.; Maria, A.; Crouzet, S.; Evrard-Todeschi, N.; Girault, J.P.; Lafont, R. Ecdysteroids from Chenopodium quinoa Willd., an ancient Andean crop of high nutritional value. Food Chem. 2011, 125, 1226–1234. [Google Scholar] [CrossRef]
- Miranda, M.; Vega-Gálvez, A.; Uribe, E.; Lópeza, J.; Martínez, E.; Rodrígueza, M.J.; Quispea, I.; Di Scalac, K. Physicochemical analysis, antioxidant capacity and vitamins of six ecotypes of Chilean quinoa (Chenopodium quinoa Willd). Procedia Food Sci. 2011, 1, 1439–1446. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhu, F. Formulation and quality attributes of quinoa food products. Food Bioprocess Technol. 2015, 9, 1–20. [Google Scholar] [CrossRef]
- Chillo, S.; Laverse, J.; Falcone, P.M.; Del Nobile, M.A. Quality of spaghetti in base amaranthus wholemeal flour added with quinoa, broad bean and chick pea. J. Food Eng. 2008, 84, 101–107. [Google Scholar] [CrossRef]
- Lorusso, A.; Verni, M.; Montemurro, M.; Coda, R.; Gobbetti, M.; Rizzello, C.G. Use of fermented quinoa flour for pasta making and evaluation of the technological and nutritional features. LWT Food Sci. Technol. 2017, 78, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Stikic, R.; Glamoclija, D.; Demin, M.; Vucelic-Radovic, B.; Jovanovic, Z.; Milojkovic-Opsenica, D.; Jacobsen, S.E.; Milovanovic, M. Agronomical and nutritional evaluation of quinoa seeds (Chenopodium quinoa Willd.) as an ingredient in bread formulations. J. Cereal Sci. 2012, 55, 132–138. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Sefora Rutella, G.; Taneyo Saa, D.L.; Gianotti, A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.S.; Zannini, E.; Czerny, M.; Arendt, E.K. Impact of sourdough fermented with Lactobacillus plantarum FST 1.7 on baking and sensory properties of gluten-free breads. Eur. Food Res. Technol. 2014, 239, 1–12. [Google Scholar] [CrossRef]
- Gallagher, E.; Gormley, T.R.; Arendt, E.K. Crust and crumb characteristics of gluten-free breads. J. Food Eng. 2003, 56, 153–161. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Cagno, R.D.; Gobbetti, M.; Rizzello, C.G. Sourdough lactic acid bacteria: Exploration of non-wheat cereal-based fermentation. Food Microbiol. 2014, 37, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R.; Calasso, R.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Arendt, E.K.; Moroni, A.; Zannini, E. Medical nutrition therapy: Use of sourdough lactic acid bacteria as a cell factory for delivering functional biomolecules and food ingredients in gluten free bread. Microb. Cell Fact. 2011, 10 (Suppl. 1), 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zannin, E.; Pontonio, E.; Waters, D.M.; Arendt, E.K. Applications of microbial fermentations for production of gluten-free products and perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 473–485. [Google Scholar] [CrossRef]
- Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. From microbial ecology to innovative applications in food quality improvements: The case of sourdough as a model matrix. J. Multidiscip. Sci. J. 2020, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Axel, C.; Brosnan, B.; Zannini, E.; Peyer, L.C.; Furey, A.; Coffey, A.; Arendt, E.K. Antifungal activities of three different Lactobacillus species and their production of antifungal carboxylic acids in wheat sourdough. Appl. Microbiol. Biotechnol. 2016, 100, 1701–1711. [Google Scholar] [CrossRef]
- Păucean, A.; Vodnar, D.C.; Socaci, S.A.; Socaciu, C. Carbohydrate metabolic conversions to lactic acid and volatile derivatives, as influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 efficiency during in vitro and sourdough fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Salvetti, A.; Torriani, S.; Felis, G.E. The genus Lactobacillus: A taxonomic update. Probiotics Antimicrob. Proteins 2012, 4, 217–226. [Google Scholar] [CrossRef]
- Hammes, W.P.; Vogel, R.F. The Genus Lactobacillus. In The Genera of Lactic Acid Bacteria; Wood, B.J.B., Holzapfel, W.H., Eds.; Blackie Academic & Professional: London, UK, 1995; pp. 19–54. [Google Scholar]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Olojede, A.O.; Sanni, A.I.; Banwo, K. Rheological, textural and nutritional properties of gluten-free sourdough made with functionally important lactic acid bacteria and yeast from Nigerian sorghum. LWT 2020, 120, 1–8. [Google Scholar] [CrossRef]
- Di Cagno, R.; Rizzello, C.G.; De Angelis, M.; Cassone, A.; Giuliani, G.; Benedusi, A.; Limitone, A.; Surico, M.F.; Gobbetti, M. Use of selected sourdough strains of Lactobacillus for removing gluten and enhancing the nutritional properties of gluten-free bread. J. Food Prot. 2008, 71, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, S.A.; Seitter, M.; Singer, U.; Brandt, M.J.; Hertel, C. Adaptability of lactic acid bacteria and yeasts to sourdoughs prepared from cereals, pseudocereals and cassava and use of competitive strains as starters. Int. J. Food Microbiol. 2009, 130, 205–212. [Google Scholar] [CrossRef]
- Moroni, A.V.; Arendt, E.K.; Bello, F.D. Biodiversity of lactic acid bacteria and yeasts in spontaneously-fermented buckwheat and teff sourdoughs. Food Microbiol. 2011, 28, 497–502. [Google Scholar] [CrossRef]
- Chiş, M.S.; Păucean, A.; Stan, L.; Mureşan, V.; Vlaic, R.A.; Man, S.; Biriş-Dorhoi, S.E.; Muste, S. Lactobacillus plantarum ATCC 8014 in quinoa sourdough adaptability and antioxidant potential. Rom. Biotechnol. Lett. 2018, 23, 13581–13591. [Google Scholar]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Bonta, V.; Pop, A.M.; Stan, L.; Beldean, B.V.; Pop, C.R.; Mureșan, V.; Muste, S. Effect of rice flour fermentation with Lactobacillus spicheri DSM 1549 on the nutritional features of gluten free muffins. Foods 2020, 9, 822. [Google Scholar] [CrossRef]
- Păucean, A.; Moldovan, O.P.; Mureşan, V.; Socaci, S.A.; Dulf, F.; Man, M.S.; Mureşan, A.E.; Muste, S. Folic acid, minerals, amino-acids, fatty acids and volatile compounds of green and red lentils. Folic acid content optimization in wheat-lentils composite flours. Chem. Cent. J. 2018, 12, 1–9. [Google Scholar]
- Urcan, A.D.; Criste, A.D.; Dezmirean, D.S.; Mărgăoan, R.; Caeiro, A.; Campos, M.G. Similarity of data from bee bread with the same taxa collected in India and Romania. Molecules 2018, 23, 2491. [Google Scholar] [CrossRef] [Green Version]
- Bunea, A.; Ruginǎ, D.O.; Pintea, A.M.; Sconţa, Z.; Bunea, C.I.; Socaciu, C. Comparative polyphenolic content and antioxidant activities of some wild and cultivated blueberries from Romania. Not. Bot. Horti Agrobot. 2011, 39, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Păucean, A.; Man, S.M.; Chis, M.S.; Mureşan, V.; Pop, C.R.; Socaci, S.A.; Mureşan, C.C.; Muste, S. Use of pseudocereals preferment made with aromatic yeast strains for enhancing wheat bread quality. Foods 2019, 8, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dordević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Teleky, B.E.; Martău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C. Exploitation of lactic acid bacteria and Baker’s yeast as single or multiple starter cultures of wheat flour dough enriched with soy flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- Schoenlechner, R. Quinoa: Its Unique Nutritional and Health-Promoting Attributes. In Gluten-Free Ancient Grains. Cereal, Pseudocereals and Legumes: Sustainaible, Nutritious and Health-Promoting Foods for the 21st Century, 1st ed.; Taylor, J., Awika, J.M., Eds.; Woodhead Publishing: Duxford, UK, 2017; pp. 105–129. [Google Scholar]
- Pellegrini, M.; Lucas-Gonzales, R.; Ricci, A.; Fontecha, J.; Fernandez-Lopez, J.; Perez-Alvarez, J.A.; Viuda-Martos, M. Chemical, fatty acid, polyphenolic profile, techno-functional and antioxidant properties of flours obtained from quinoa (Chenopodium quinoa Willd) seeds. Ind. Crop. Prod. 2018, 111, 38–46. [Google Scholar] [CrossRef]
- Pasko, P.; Barton, H.; Zagrodzki, P.; Izewska, A.; Krosniak, M.; Gawlik, M.; Gawlik, M.; Gorinstein, S. Effect of diet supplemented with quinoa seeds on oxidative status in plasma and selected tissues of high fructose-fed rats. Plant Foods Hum. Nutr. 2010, 65, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.; Encina-Zelada, C.; Barros, L.; Gonzales-Barron, U.; Cadavez, V.; Ferreira, C.F.R. Chemical and nutritional characterization of Chenopodium quinoa Willd (quinoa) grains: A good alternative to nutritious food. Food Chem. 2019, 280, 110–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeli, M.; Silva, P.M.; Massuela, C.D.; Whan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Hönninge, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An overview of the potentials of the “Golden Grain” and socio-economic and environmental aspects of its cultivation and marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Luna, A.; Fuentes, F.; Rivadeneyra, E.; Hernández, C.; Díaz-Sobac, R. Nutrimental content and functional properties of Quinoa flour from Chile and Mexico. Cienc. E Investig. Agrar. 2019, 46, 144–153. [Google Scholar] [CrossRef]
- Romano, N.; Ureta, M.M.; Guerrero-Sánchez, M.; Gómez-Zavaglia, A. Nutritional and technological properties of a quinoa (Chenopodium quinoa Willd.) spray-dried powdered extract. Food Res. Int. 2020, 129, 1–11. [Google Scholar] [CrossRef]
- Franco, W.; Pérez-Díaz, I.M.; Connelly, L.; Diaz, J.T. Isolation of exopolysaccharide-producing yeast and lactic acid bacteria from quinoa (Chenopodium quinoa) sourdough fermentation. Foods 2020, 9, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saubade, F.; Hemery, Y.M.; Rochette, I.; Guyot, J.P.; Humblot, C. Influence of fermentation and other processing steps on the folate content of a traditional African cereal-based fermented food. Int. J. Food Microbiol. 2018, 266, 79–86. [Google Scholar] [CrossRef]
- Bilgiçl, N.; İbanoğlu, Ş. Effect of pseudo cereal flours on some physical, chemical and sensory properties of bread. J. Food Sci. Technol. 2015, 52, 7525–7529. [Google Scholar] [CrossRef]
- Schoenlechner, R.; Wendner, M.; Siebenhandl-Ehn, S.; Berghofer, E. Pseudocereals as alternative sources for high folate content in staple foods. J. Cereal Sci. 2010, 52, 475–479. [Google Scholar] [CrossRef]
- Li, G.; Zhu, F. Physicochemical properties of quinoa flour as affected by starch interactions. Food Chem. 2017, 221, 1560–1568. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Puig, E.; Monedero, V.; Haros, M. Bread with whole quinoa flour and bifidobacterial phytases increases dietary mineral intake and bioavailability. LWT Food Sci. Technol. 2015, 60, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Valcárcel-Yamani, B.; Caetano da Silva Lannes, S. Applications of Quinoa (Chenopodium quinoa Willd) and amaranth (Amaranthus spp.) and their influence in the nutritional value of cereal based foods. Food Public Health 2012, 2, 265–275. [Google Scholar]
- Prado, F.E.; Fernández-Turiel, J.L.; Tsarouchi, M.; Psaras, G.K.; González, J.A. Variation of seed mineral concentrations in seven quinoa cultivars grown in two agroecological sites. Cereal Chem. 2014, 91, 453–459. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.A.; Konishi, Y.; Bruno, M.; Valoy, M.; Prado, F.E. Interrelationships among seed yield, total protein and amino acid composition of ten quinoa (Chenopodium quinoa) cultivars from two different agroecological regions. J. Sci. Food Agric. 2012, 92, 1222–1229. [Google Scholar] [CrossRef]
- Carciochi, R.A.; Dimitrov, K. Optimization of antioxidant phenolic compounds extraction from quinoa (Chenopodium quinoa) seeds. J. Food Sci. Technol. 2014, 52, 4396–4404. [Google Scholar] [CrossRef]
- De Carvalho, F.G.; Ovídio, P.P.; Padovan, G.J.; Jordão Junior, A.A.; Marchini, J.S.; Navarro, A.M. Metabolic parameters of postmenopausal women after quinoa or corn flakes intake-a prospective and double-blind study. Int. J. Food Sci. Nutr. 2014, 65, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Arneja, I.; Tanwar, B.; Chauhan, A. Nutritional composition and health benefits of golden grain of 21st century, quinoa (Chenopodium quinoa willd.): A review. Pak. J. Nutr. 2015, 14, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Vrancken, G.; Ravyts, F.; Rimaux, T.; Weckx, S. Biodiversity, ecological determinants, and metabolic exploitation of sourdough microbiota. Food Microbiol. 2009, 26, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flour. Int. J. Food Microbiol. 2018, 302, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Von Wright, A.; Axerlsson, L. Lactic Acid Bacteria: An Introduction. In Lactic Acid Bacteria: Microbiological and Functional Aspects, 4th ed.; Lahtinen, S., Ouwehand, A.C., Salminen, S., Von Wright, A., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA; London, UK, 2011; pp. 1–17. [Google Scholar]
- Petrova, P.; Petrov, K. Lactic acid fermentation of cereals and pseudocereals: Ancient nutritional biotechnologies with modern applications. Nutrients 2020, 12, 1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrizo, S.L.; de Moreno de LeBlanc, A.; LeBlanc, J.G.; Rollán, G.C. Quinoa pasta fermented with lactic acid bacteria prevents nutritional deficiencies in mice. Food Res. Int. 2020, 127, 1–12. [Google Scholar] [CrossRef]
- Florou-paneri, P.; Christaki, E.; Bonos, E. Lactic Acid Bacteria as Source of Functional Ingredients. In Lactic Acid Bacteria, 1st ed.; Kongo, M., Ed.; Intech Open Limited: Rijeka, Croatia, 2013; pp. 1–26. [Google Scholar]
- Leblanc, J.G.; Savoy de Giori, G.; Smid, E.J.; Hugenholtz, J.; Sesma, F. Folate Production by Lactic Acid Bacteria and Other Food-Grade Microorganisms. In Bioactive Foods as Dietary Interventions for Liver and Gastrointestinal Disease, 1st ed.; Watson, R.R., Preedy, V., Eds.; Academis Press: Cambridge, MA, USA, 2007; pp. 329–339. [Google Scholar]
- Carrizo, S.L.; Montes de Oca, C.E.; Laiño, J.E.; Suarez, N.E.; Vignolo, G.; LeBlanc, J.G.; Rollán, G. Ancestral Andean grain quinoa as source of lactic acid bacteria capable to degrade phytate and produce B-group vitamins. Food Res. Int. 2016, 89, 488–494. [Google Scholar] [CrossRef]
- Motta, C.; Delgado, I.; Matos, A.S.; Gonzales, G.B.; Torres, D.; Santos, M.; Chandra-Hioe, M.V.; Arcot, J.; Castanheira, I. Folates in quinoa (Chenopodium quinoa), amaranth (Amaranthus sp.) and buckwheat (Fagopyrum esculentum): Influence of cooking and malting. J. Food Compos. Anal. 2017, 64, 181–187. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvucci, E.; Leblanc, J.G.; Perez, G. Technological properties of Lactic Acid Bacteria isolated from raw cereal material. LWT Food Sci. Technol. 2016, 70, 1–26. [Google Scholar] [CrossRef]
- Taylor, J.R.N.; Belton, P.S.; Beta, T.; Duodu, K.G. Increasing the utilisation of sorghum, millets and pseudocereals: Developments in the science of their phenolic phytochemicals, biofortification and protein functionality. J. Cereal Sci. 2014, 59, 257–275. [Google Scholar] [CrossRef] [Green Version]
- Magala, M.; Kohajdová, Z.; Karovičová, J. Degradation of phytic acid during fermentation of cereal substrates. J. Cereal Sci. 2015, 61, 94–96. [Google Scholar] [CrossRef]
- Lopez, H.W.; Duclos, V.; Coudray, C.; Krespine, V.; Feillet-Coudray, C.; Messager, A.; Demigne’, C.; Remesy, C. Making bread with sourdough improves mineral bioavailability from reconstituted whole wheat flour in rats. Nutrition 2003, 19, 524–530. [Google Scholar] [CrossRef]
- Taylor, J.R.N.; Parker, M.L. Quinoa. In Pseudocereals and Less Common Cereals, Grain Properties and Utilization Potential, 1st ed.; Belton, S., Taylor, J.R.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 93–122. [Google Scholar]
- Saa, D.T.; Di Silvestro, R.; Dinelli, G.; Gianotti, A. Effect of sourdough fermentation and baking process severity on dietary fibre and phenolic compounds of immature wheat flour bread. LWT Food Sci. Technol. 2017, 83, 26–32. [Google Scholar] [CrossRef]
- Montemurro, M.; Coda, R.; Rizzello, C.G. Recent advances in the use of Sourdough. Foods 2019, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolívar-Monsalve, J.; Ceballos-González, C.; Ramírez-Toro, C.; Bolívar, G.A. Reduction in saponin content and production of gluten-free cream soup base using quinoa fermented with Lactobacillus plantarum. J. Food Process. Preserv. 2018, 42, 1–10. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Zhang, Y.; Yang, W.; Ma, G.; Ma, N.; Hu, Q.; Pei, F. A novel lactic acid bacterium for improving the quality and shelf life of whole wheat bread. Food Control 2020, 109, 1–9. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.S.; Zannini, E.; Czerny, M.; Arendt, E.K. Influence of dextran-producing Weissella cibaria on baking properties and sensory profile of gluten-free and wheat breads. Int. J. Food Microbiol. 2014, 172, 83–91. [Google Scholar] [CrossRef]
- Turkut, G.M.; Cakmak, H.; Kumcuoglu, S.; Tavman, S. Effect of quinoa flour on gluten-free bread batter rheology and bread quality. J. Cereal Sci. 2016, 69, 174–181. [Google Scholar] [CrossRef]
- Mellado-Mojica, E.; Seeram, N.P.; López, M.G. Comparative analysis of maple syrups and natural sweeteners: Carbohydrates composition and classification (differentiation) by HPAEC-PAD and FTIR spectroscopy-chemometrics. J. Food Compos. Anal. 2016, 52, 1–8. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Blackburn, J.; Lada, R.R. A comparative physicochemical analysis of maple (Acer saccharum Marsh.) syrup produced in North America with special emphasis on seasonal changes in Nova Scotia maple syrup composition. J. Food Compos. Anal. 2020, 92, 1–31. [Google Scholar] [CrossRef]
- Debonne, E.; Vermeulen, A.; Bouboutiefskia, N.; Ruyssen, T.; Van Bockstaeled, F.; Eeckhout, M.; Devlieghere, F. Modelling and validation of the antifungal activity of DL-3-phenyllactic acid and acetic acid on bread spoilage moulds. Food Microbiol. 2020, 88, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Zoumpopoulou, G.; Georgalaki, M.; Alexandraki, V.; Kazou, M.; Anastasiou, R.; Tsakalidou, E. Sourdough Bread, Innovations in Traditional Foods; Galanakis, C.M., Ed.; Woodhead Publishing: Cambridge, UK, 2019; pp. 127–158. [Google Scholar]
- Reddy, G.; Altaf, M.; Naveena, B.J.; Venkateshwar, M.; Kumar, E.V. Amylolytic bacterial lactic acid fermentation—A review. Biotechnol. Adv. 2008, 26, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Kefalas, P.; Kotzamanidis, S.; Sabanis, D.; Yupsani, A.; Kefala, L.A.; Kokkalis, A.; Yupsanis, T. Bread making of durum wheat with chickpea sourdough or compressed baker’s yeast. J. Food. Qual. 2009, 32, 644–668. [Google Scholar] [CrossRef]
- Takata, Y.; Shu, X.O.; Buchowski, M.S.; Munro, H.M.; Wen, W.; Steinwandel, M.D.; Hargreaves, M.K.; Blot, W.J.; Cai, Q. Food intake of folate, folic acid and other B vitamins with lung cancer risk in a low-income population in the Southeastern United States. Eur. J. Nutr. 2020, 59, 671–683. [Google Scholar] [CrossRef]
- Chis, M.S.; Păucean, A.; Man, S.M.; Muresan, V.; Socaci, S.A.; Pop, A.; Stan, L.; Rusu, B.; Muste, S. Textural and sensory features changes of gluten free muffins based on rice sourdough fermented with Lactobacillus spicheri DSM 15429. Foods 2020, 9, 363. [Google Scholar] [CrossRef] [Green Version]
- Naqash, F.; Gani, A.; Gani, A.; Masoodi, F.A. Gluten-Free baking: Combating the challenges—A review. Trends Food Sci. Technol. 2017, 66, 98–107. [Google Scholar] [CrossRef]
- Capriles, V.D.; dos Santos, F.G.; Arêas, J.A.G. Gluten-Free breadmaking: Improving nutritional and bioactive compounds. J. Cereal Sci. 2016, 67, 83–91. [Google Scholar] [CrossRef]
- Li, L.; Shakhawat Hossain, M.D.; Ma, H.; Yang, Q.; Gong, X.; Yang, P.; Feng, B. Comparative metabolomics reveals diFferences in flavonoid metabolites among different coloured buckwheat flowers. J. Food Compos. Anal. 2019, 85, 1–8. [Google Scholar] [CrossRef]
- Chlopicka, J.; Pasko, P.; Gorinstein, S.; Jedryas, A.; Zagrodzki, P. Total phenolic and total flavonoid content, antioxidant activity and sensory evaluation of pseudocereal breads. LWT Food Sci. Technol. 2012, 46, 548–555. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Lorusso, A.; Russo, V.; Pinto, D.; Marzani, B.; Gobbetti, M. Improving the antioxidant properties of quinoa flour through fermentation with selected autochthonous lactic acid bacteria. Int. J. Food Microbiol. 2017, 241, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Cannas, M.; Pulina, S.; Conte, P.; del Caro, A.; Urgeghe, P.P.; Piga, A.; Fadda, C. Efect of substitution of rice flour with quinoa flour on the chemical-physical, nutritional, volatile and sensory parameters of gluten-free ladyfinger biscuits. Foods 2020, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- Banu, I.; Vasilean, I.; Aprodu, I. Effect of lactic fermentation on antioxidant capacity of rye sourdough and bread. Food Sci. Technol. Res. 2010, 16, 571–576. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
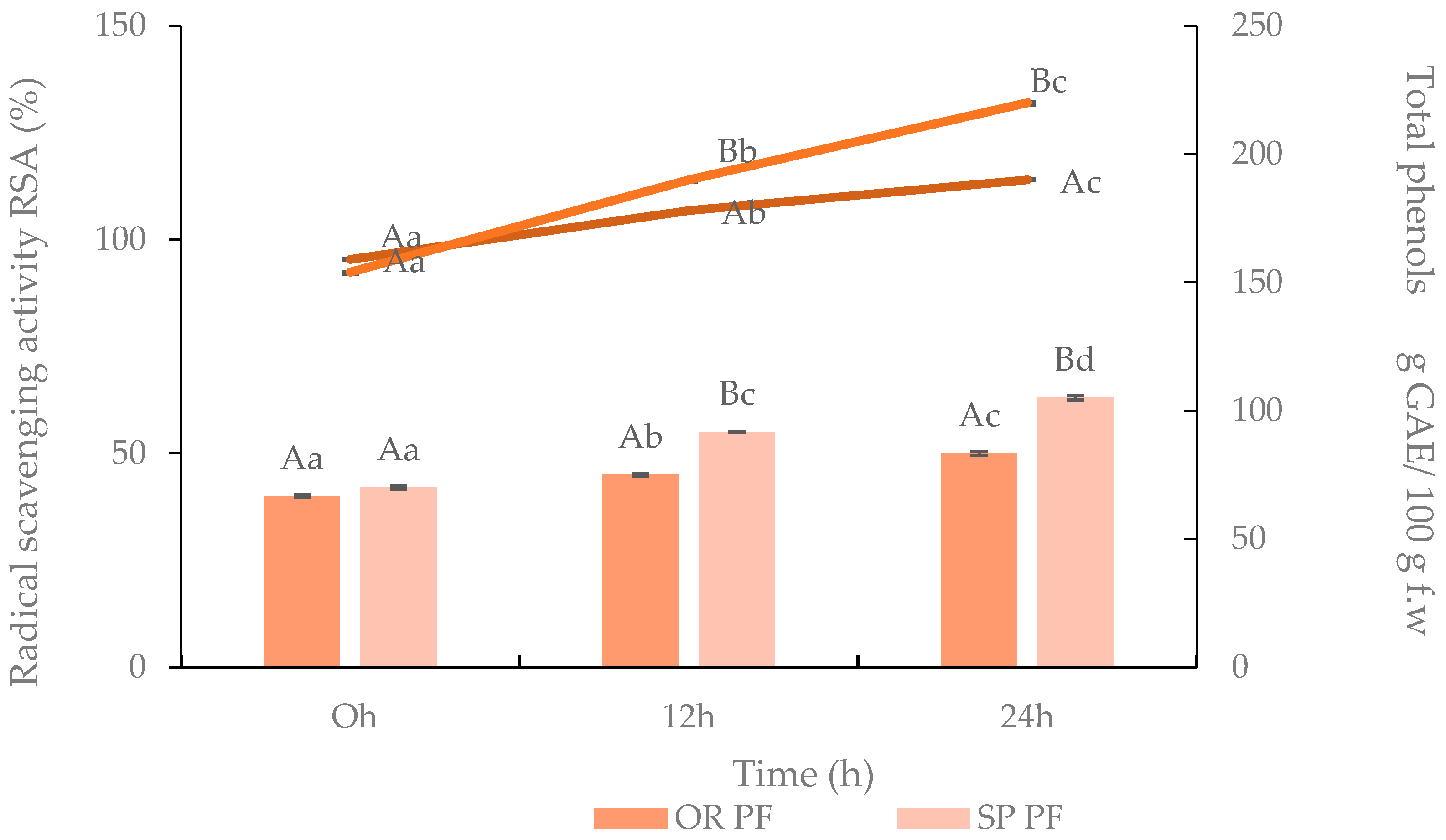{kind=link}
| Parameters | QWF | Retention Times (min) |
|---|---|---|
| Carbohydrates (mg/g f.w.) | ||
| Maltose | 6.64 ± 0.20 | 8.87 |
| Glucose | 89.45 ± 0.13 | 10.24 |
| Fructose | 12.12 ± 0.15 | 10.88 |
| Organic acids (mg/g f.w.) | ||
| Citric acid | 8.59 ± 0.30 | 9.39 |
| Lactic acid | n.d. | 13.76 |
| Acetic acid | n.d. | 15.92 |
| Parameters | QWF |
|---|---|
| Minerals, mg/100 g f.w. | |
| Calcium (Ca) | 18.09 ± 0.30 |
| Magnesium (Mg) | 303.43 ± 0.17 |
| Potassium (K) | 813.92 ± 0.11 |
| Iron (Fe) | 3.02 ± 0.20 |
| Copper (Cu) | 0.96 ± 0.03 |
| Zinc (Zn) | 1.82 ± 0.02 |
| Manganese (Mn) | 2.50 ± 0.01 |
| Chromium (Cr) | n.d. |
| Samples | Maltose mg/g f.w. | Glucose | Fructose | Citric Acid | Lactic Acid | Acetic Acid |
|---|---|---|---|---|---|---|
| mg/g f.w. | mg/g f.w. | mg/g f.w. | mg/g f.w. | mg/g f.w. | ||
| OR 0 h | 3.012 ± 0.02 Aa | 42.662 ± 0.34 Ac | 4.19 ± 0.19 Aabc | 5.25 ± 0.33 Abc | n.d. | n.d. |
| SP 0 h | 3.074 ± 0.21 Aa | 42.034 ± 0.54 Ac | 4.369 ± 0.28 Babc | 5.06 ± 0.22 Acd | n.d. | n.d. |
| OR 4 h | 2.979 ± 0.31 Aa | 46.576 ± 0.32 Abc | 4.21 ± 0.29 Babc | 4.26 ± 0.11 Ae | n.d. | n.d. |
| SP 4 h | 3.053 ± 0.11 Aa | 46.085 ± 0.53 Abc | 3.15 ± 0.22 Ad | 4.92 ± 0.45 Bcd | n.d. | n.d. |
| OR 8 h | 2.848 ± 0.22 Aa | 51.126 ± 0.23 Abc | 4.76 ± 0.39 Ba | 4.74 ± 0.34 Ad | n.d. | n.d. |
| SP 8 h | 2.916 ± 0.15 Aa | 67.672 ± 0.61 Ba | 3.75 ± 0.2 Abcd | 5.9 ± 0.36 Ba | n.d. | n.d. |
| OR 12 h | 2.85 ± 0.25 Aa | 55.01 ±0.22 Bb | 4.48 ± 0.5 Bab | 5.06 ± 0.33 Acd | 2.42 ± 0.02 Aa | 0.50 ± 0.01 Aa |
| SP 12 h | 1.814 ± 0.14 Bb | 53.00 ± 0.45 Abc | 3.08 ± 0.4 Ad | 5.54 ± 0.12 Bab | 4.60 ± 0.05 Bb | 0.84 ± 0.02 Bab |
| OR 24 h | 2.09 ± 0.03 Ab | 46.02 ± 0.39 Bbc | 3.66 ± 0.6 Bcd | 5.15 ± 0.44 Bbcd | 3.81 ± 0.31 Ab | 2.98 ± 0.21 Bc |
| SP 24 h | 1.05 ± 0.30 Bc | 22.00 ± 0.27 Ad | 1.69 ± 0.4 Ae | 4.89 ± 0.33 Acd | 8.50 ± 0.5 Bc | 1.40 ± 0.21 Ab |
| Samples | Ca | Mg | K | Fe | Cu | Zn | Mn |
|---|---|---|---|---|---|---|---|
| OR 0 h | 8.06 ± 0.13 Aa | 156.09 ± 0.34 Aa | 330.9 ± 0.89 Aa | 0.61 ± 0.02 Aa | 0.11 ± 0.01 Aa | 0.46 ± 0.02 Aa | 0.55 ± 0.02 Aa |
| SP 0 h | 8.12 ± 0.11 Ba | 151.00 ± 0.23 Aa | 325.01 ± 0.99 Aa | 0.63 ± 0.03 Aab | 0.12 ± 0.02 Aa | 0.49 ± 0.01 Aa | 0.59 ± 0.06 Aab |
| OR 4 h | 8.23± 0.17 Aa | 159.03 ± 0.03 Aa | 341.08 ± 0.77 Aa | 0.69 ± 0.05 Aab | 0.13 ± 0.04 Aa | 0.50 ± 0.03 Aa | 0.57 ± 0.05 Aa |
| SP 4 h | 8.55 ± 0.33 Ba | 163.09 ± 0.03 Bab | 378.04 ± 0.88 Bbc | 0.75 ± 0.34 Aabc | 0.17 ± 0.01 Aab | 0.53 ± 0.04 Aa | 0.65 ± 0.07 Aab |
| OR 8 h | 8.9 ± 0.22 Aa | 172.04 ± 0.23 Aabc | 353.56 ± 0.89 Aab | 0.71 ± 0.11 Aab | 0.23 ± 0.01 Aab | 0.60 ± 0.01 Aa | 0.60 ± 0.06 Aab |
| SP 8 h | 9.6 ± 0.34 Bab | 176.89 ± 0.56 Babc | 456.67 ± 0.67 Bef | 0.89 ± 0.33 Bd | 0.29 ± 0.02 Ab | 0.72 ± 0.03 Aa | 0.72 ± 0.08 Abc |
| OR 12 h | 9.53 ± 0.11 Aab | 171.23 ± 0.45 Aabc | 386.4 ± 0.69 Abc | 0.77 ± 0.11 Abcd | 0.16 ± 0.03 Aab | 0.63 ± 0.02 Aa | 0.58 ± 0.06 Aa |
| SP 12 h | 11.67 ± 0.22 Bc | 198.98 ± 0.64 Bc | 407.61 ± 0.88 Bcd | 1.11 ± 0.02 Be | 0.51 ± 0.02 Bc | 0.99 ± 0.01 Bb | 0.89 ± 0.05 Bd |
| OR 24 h | 10.86 ± 0.45 Abc | 189.21 ± 0.59 Abc | 427.53 ± 0.79 Ade | 0.87 ± 0.02 Acd | 0.21 ± 0.04 Aab | 0.62 ± 0.02 Aa | 0.78 ± 0.03 Acd |
| SP 24 h | 17.06 ± 0.32 Bd | 294.59 ± 0.89 Bd | 486.44 ± 0.98 Bf | 1.42 ± 0.04 Bf | 0.79 ± 0.05 Bd | 1.68 ± 0.00 Bc | 1.23 ± 0.07 Be |
| Sample | Maltose | Glucose | Fructose | Citric Acid | Lactic Acid | Acetic Acid |
|---|---|---|---|---|---|---|
| mg/g f.w. | mg/g f.w. | mg/g f.w. | mg/g f.w. | mg/g f.w. | mg/g f.w. | |
| OR PF 0 h | 4.43 ± 0.03 Ade | 20.06 ± 0.31 Ad | 14.72 ± 0.56 Ade | 1.72 ± 0.05 Aa | n.d. | n.d. |
| SP PF 0 h | 4.15 ± 0.23 Acd | 20.18 ± 0.22 Ad | 14.17 ± 0.45 Acd | 1.73 ± 0.03 Aab | n.d. | n.d. |
| OR PF 12 h | 4.84 ± 0.23 Be | 19.58 ± 0.34 Bd | 15.39 ± 0.39 Be | 1.72 ± 0.11 Aa | 0.44 ± 0.02 Aa | 0.80 ± 0.02 Ba |
| SP PF 12 h | 3.14 ± 0.11 Ab | 17.67± 0.67 Ab | 12.92 ± 0.56 Ab | 1.86 ± 0.43 Aac | 0.93 ± 0.05 Bc | 0.62 ± 0.04 Aa |
| OR PF 24 h | 3.82 ± 0.03 Bc | 18.65 ± 0.55 Bc | 13.55 ± 0.88 Bbc | 2.27 ± 0.02 Ad | 0.82 ± 0.07 Ab | 1.69 ± 0.09 Bb |
| SP PF 24 h | 2.58 ± 0.34 Aa | 16.37 ± 0.77 Aa | 10.57 ± 0.65 Aa | 2.30 ± 0.27 Ad | 1.52 ± 0.08 Bd | 1.02 ± 0.09 Aa |
| Samples | Ca | Mg | K | Fe | Cu | Zn | Mn |
|---|---|---|---|---|---|---|---|
| OR PF 0 h | 8.21 ± 0.03 Aa | 147.16 ± 0.07 Aa | 411.67 ± 0.03 Aa | 0.99 ± 0.02 Aa | 0.40 ± 0.05 Aa | 0.90 ± 0.23 Aa | 0.89 ± 0.02 Aa |
| SP PF 0 h | 8.34 ± 0.05 Aa | 148.32 ± 0.09 Aa | 410.36 ± 0.29 Aab | 1.00 ± 0.03 Aa | 0.41 ± 0.04 Aa | 0.93 ± 0.02 Aa | 0.90 ± 0.05 Aa |
| OR PF 12 h | 10.30 ± 0.02 Aab | 151.00 ± 0.07 Aa | 420.69 ± 0.34 Aab | 1.03 ± 0.05 Aa | 0.41 ± 0.01 Aa | 1.09 ± 0.05 Ab | 0.99 ± 0.04 Ab |
| SP PF 12 h | 12.06 ± 0.01 Bd | 172.12 ± 0.04 Bb | 465.24 ± 0.69 Bc | 1.30 ± 0.06 Ab | 0.40 ± 0.34 Ab | 1.34 ± 0.07 Ac | 1.22 ± 0.45 Ac |
| OR PF 24 h | 11.20 ± 0.30 Ac | 159.81 ± 0.02 Aab | 430.95 ± 0.72 Ab | 1.09 ± 0.03 Aa | 0.45 ± 0.21 Aab | 1.20 ± 0.11 Ab | 1.12 ± 0.34 Ad |
| SP PF 24 h | 14.25 ± 0.65 Be | 195.99 ± 0.02 Bc | 490.20 ± 0.89 Bd | 1.55 ± 0.01 Bc | 0.89 ± 0.11 Bc | 1.75 ± 0.10 Bd | 1.51 ± 0.22 Be |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.-E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds. Appl. Sci. 2020, 10, 7140. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207140
Chiş MS, Păucean A, Man SM, Vodnar DC, Teleky B-E, Pop CR, Stan L, Borsai O, Kadar CB, Urcan AC, et al. Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds. Applied Sciences. 2020; 10(20):7140. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207140
Chicago/Turabian StyleChiş, Maria Simona, Adriana Păucean, Simona Maria Man, Dan Cristian Vodnar, Bernadette-Emoke Teleky, Carmen Rodica Pop, Laura Stan, Orsolya Borsai, Csaba Balasz Kadar, Adriana Cristina Urcan, and et al. 2020. "Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds" Applied Sciences 10, no. 20: 7140. https://0-doi-org.brum.beds.ac.uk/10.3390/app10207140