
Reuse of Pruning Waste from Subtropical Fruit Trees and Urban Gardens as a Source of Nutrients: Changes in the Physical, Chemical, and Biological Properties of the Soil

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
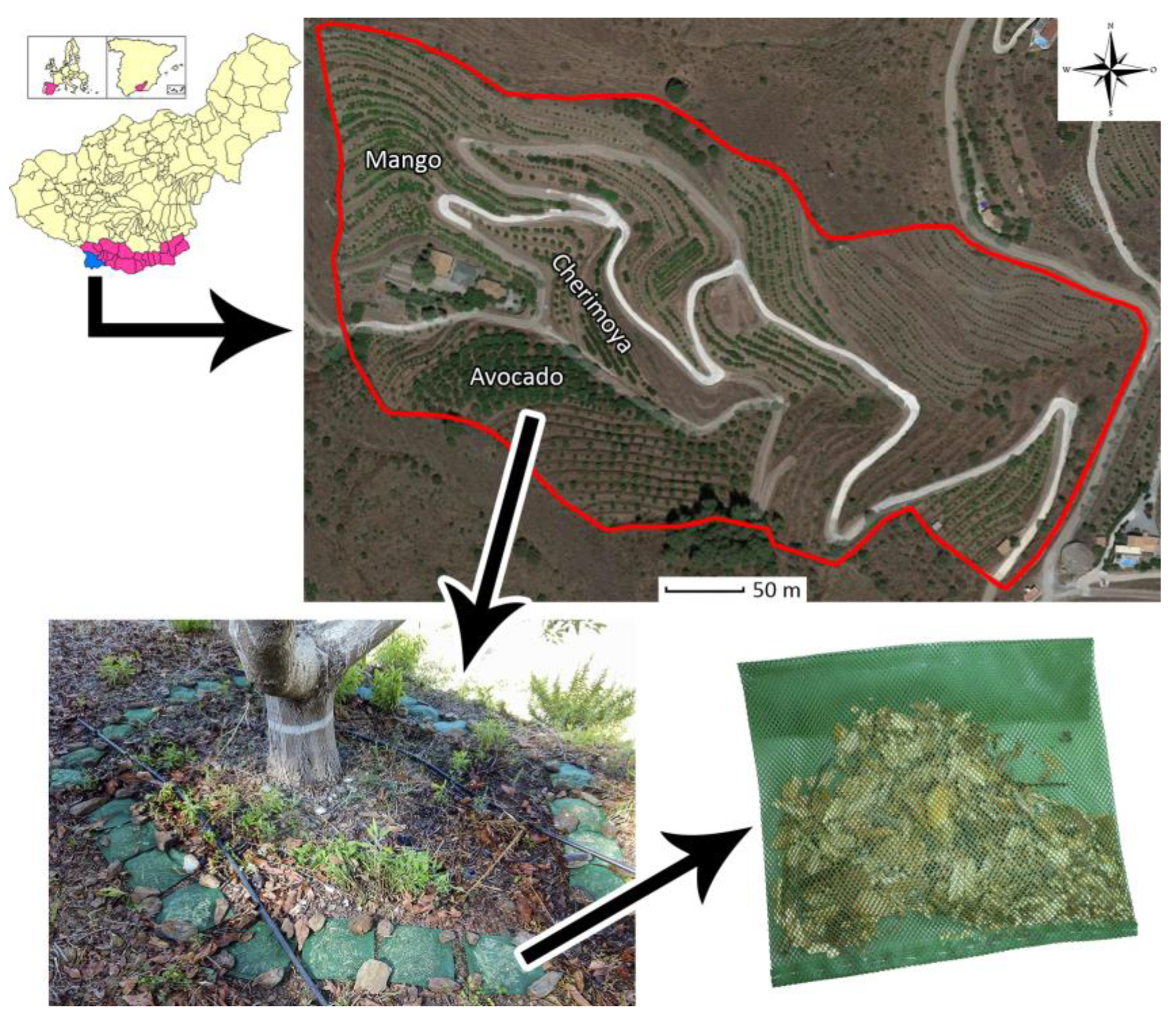
2.1. Study Sites and Experimental Design
2.2. Sample Preparation and Laboratory Analysis
2.3. Nutrient Release
2.4. Analysis of Soil Enzymatic Activities
2.5. Statistical Analysis
3. Results
3.1. Pruning Waste
3.1.1. Initial and Final Composition of the Pruning Waste
3.1.2. Weight Loss
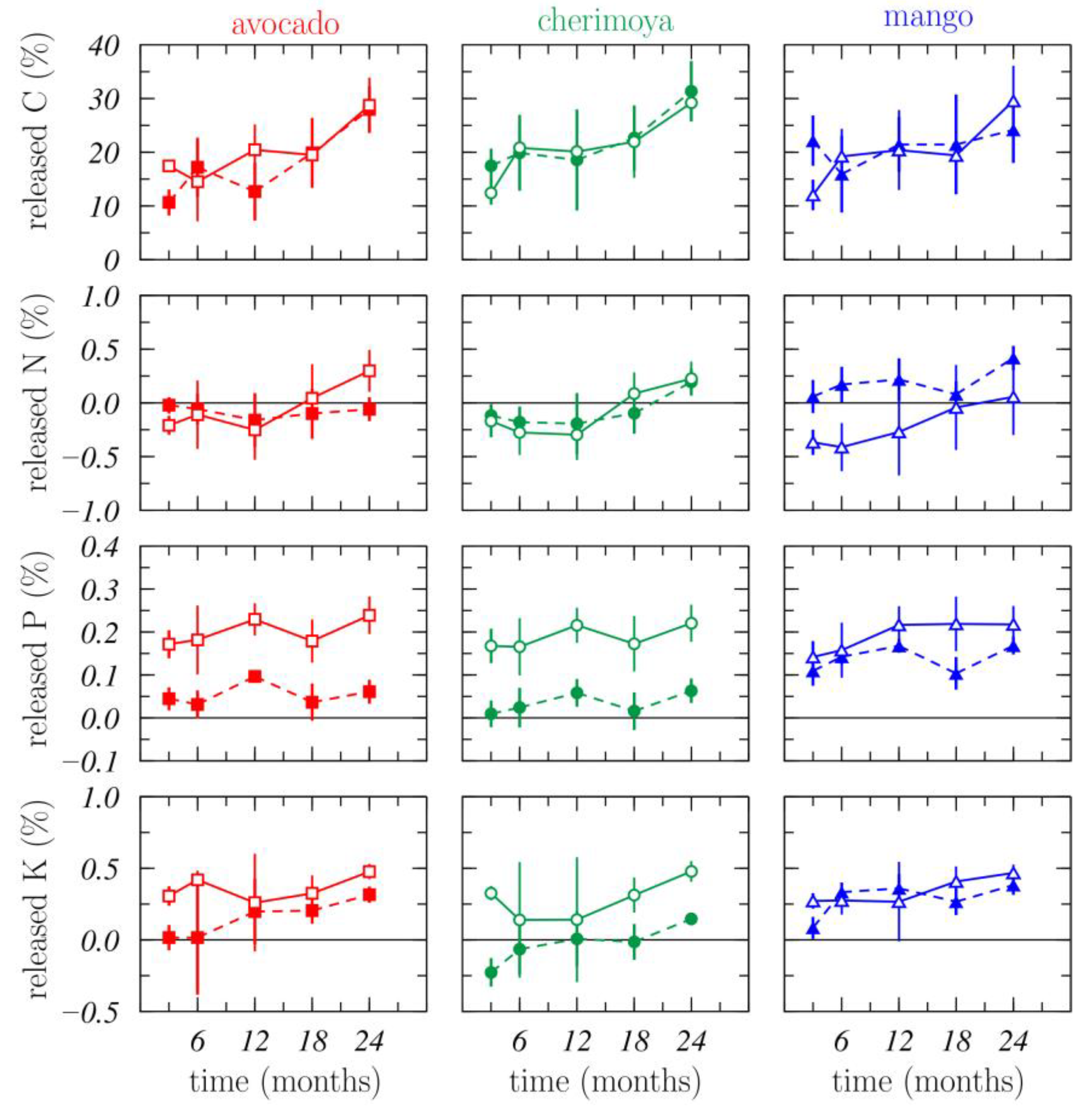
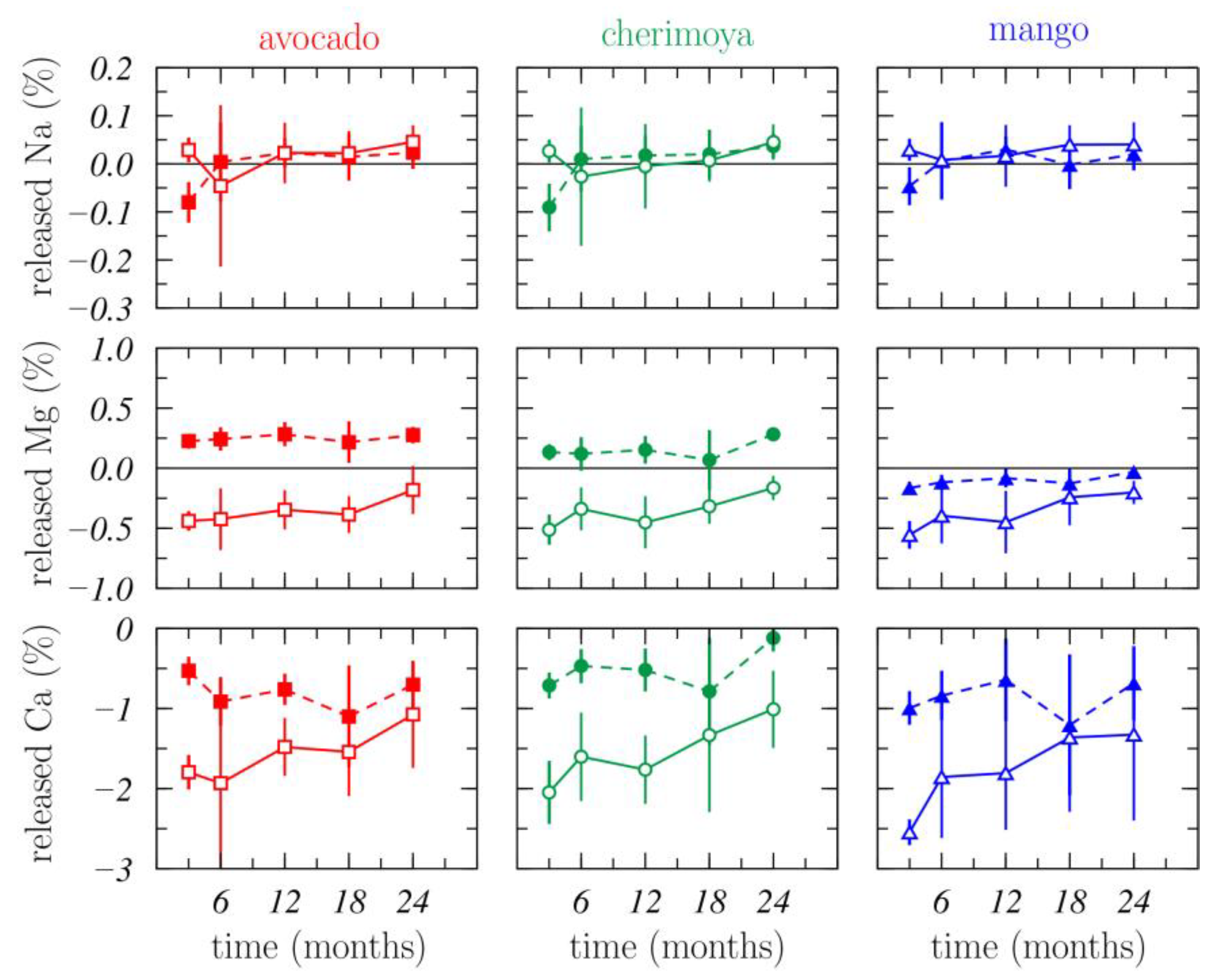
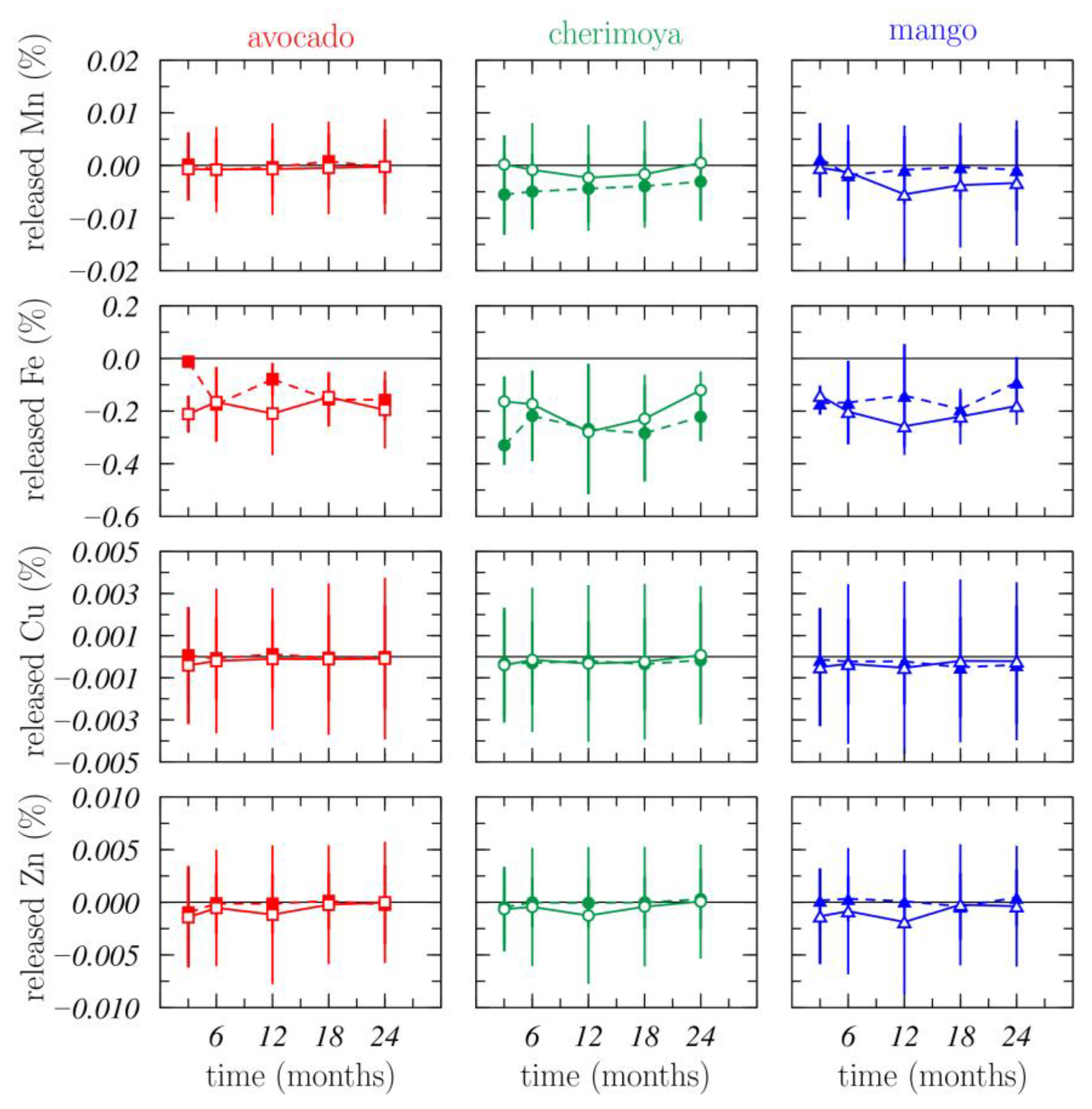
3.1.3. Nutrient Release
3.1.4. Nutrient Dynamics
3.2. Soils
3.2.1. Time Course of Soil Properties
3.2.2. Soil Enzymatic Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kuokkanen, A.; Mikkilä, M.; Kuisma, M.; Kahiluoto, H.; Linnanen, L. The need for policy to address the food system lock-in: A case study of the Finnish context. J. Clean. Prod. 2017, 140, 933–944. [Google Scholar] [CrossRef]
- van der Wiel, B.Z.; Weijma, J.; van Middelaar, C.E.; Kleinke, M.; Buisman, C.J.N.; Wichern, F. Restoring nutrient circularity: A review of nutrient stock and flow analyses of local agro-food-waste systems. Resour. Conserv. Recycl. X 2019, 3, 100014. [Google Scholar] [CrossRef]
- Bellarby, J.; Surridge, B.W.; Haygarth, P.M.; Liu, K.; Siciliano, G.; Smith, L.; Rahn, C.; Meng, F. The stocks and flows of nitrogen, phosphorus and potassium across a 30-year time series for agriculture in Huantai county, China. Sci. Total Environ. 2018, 619, 606–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Zhang, W.; Sheng, R.; Liu, Y.; Hou, H.; Liu, F.; Ma, G.; Wei, W.; Qin, H. Long-term partial replacement of mineral fertilizer with in situ crop residues ensures continued rice yields and soil fertility: A case study of a 27-year field experiment in subtropical China. Sci. Total Environ. 2021, 787, 147523. [Google Scholar] [CrossRef] [PubMed]
- Hansen, V.; Eriksen, J.; Jensen, L.S.; Thorup-Kristensen, K.; Magid, J. Towards integrated cover crop management: N, P and S release from aboveground and belowground residues. Agric. Ecosyst. Environ. 2021, 313, 107392. [Google Scholar] [CrossRef]
- Sukitprapanon, T.S.; Jantamenchai, M.; Tulaphitak, D.; Vityakon, P. Nutrient composition of diverse organic residues and their long-term effects on available nutrients in a tropical sandy soil. Heliyon 2020, 6, e05601. [Google Scholar] [CrossRef] [PubMed]
- Mikula, K.; Izydorczyk, G.; Skrzypczak, D.; Mironiuk, M.; Moustakas, K.; Witek-Krowiak, A.; Chojnacka, K. Controlled release micronutrient fertilizers for precision agriculture—A review. Sci. Total Environ. 2020, 712, 136365. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.J.; Smith, P. Carbon losses from soil and its consequences for land-use management. Sci. Total Environ. 2007, 382, 165–190. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Ncube, B.; Mulidzi, R.; Lewu, F.B. Management impact and benefit of cover crops on soil quality: A review. Soil Tillage Res. 2020, 204, 104717. [Google Scholar] [CrossRef]
- Stavi, I.; Argaman, E. No-till systems: Gains and drawbacks for carbon sequestration, ecosystem services and environmental health. Carbon Manag. 2014, 5, 123–125. [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Yu, Q.; Zhang, Y.; Wang, R.; Li, J.; Wang, X. No tillage increases soil organic carbon storage and decreases carbon dioxide emission in the crop residue-returned farming system. J. Environ. Manag. 2020, 261, 110261. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-Fernández, R.; De Torres, M.R.R.; Román-Vázquez, J.; González-Fernández, P.; Carbonell-Bojollo, R. Macronutrients released during the decomposition of pruning residues used as plant cover and their effect on soil fertility. J. Agric. Sci. 2015, 153, 615–630. [Google Scholar] [CrossRef]
- Prescott, C.E. Do rates of litter decomposition tell us anything we really need to know? For. Ecol. Manag. 2005, 220, 66–74. [Google Scholar] [CrossRef]
- Smil, V. Crop Residues: Agriculture’s Largest Harvest: Crop residues incorporate more than half of the world’s agricultural phytomass. Bioscience 1999, 49, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Moritsuka, N.; Yanai, J.; Mori, K.; Kosaki, T. Biotic and abiotic processes of nitrogen immobilization in the soil-residue interface. Soil Biol. Biochem. 2004, 36, 1141–1148. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Li-Hua, T.U.; Hong-Ling, H.U.; Ting-Xing, H.U.; Zhang, J.; Xian-Wei, L.I.; Li, L.I.U.; Yin-Long, X.I.A.O.; Gang, C.H.E.N.; Ren-Hong, L.I. Litterfall, litter decomposition, and nutrient dynamics in two subtropical bamboo plantations of China. Pedosphere 2014, 24, 84–97. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Aguilar Bustamante, V. Análisis de datos provenientes de ensayos de descomposición y mineralización de residuos vegetales. Calera 2005, 5, 50–54. [Google Scholar]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Zibilske, L.M.; Makus, D.J. Black oat cover crop management effects on soil temperature and biological properties on a Mollisol in Texas, USA. Geoderma 2009, 149, 379–385. [Google Scholar] [CrossRef]
- Feng, H.; Sekaran, U.; Wang, T.; Kumar, S. On-farm assessment of cover cropping effects on soil C and N pools, enzyme activities, and microbial community structure. J. Agric. Sci. 2021, 1–11. [Google Scholar] [CrossRef]
- Reyes-Martín, M.P.; Martínez-Cartas, M.L.; Ortiz-Bernad, I.; San-Emeterio, L.M.; Fernández-Ondoño, E. Mineralization of bagged pruning waste in agrosystem on the subtropical coast of Andalusia (Spain). J. Agric. Sci. 2020, 158, 634–645. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015; International soil classification system for naming soils and creating legends for soil maps; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; 192p. [Google Scholar]
- Falconer, J.G.; Wright, J.W.; Beall, H.W. The decomposition of certain types of forest litter under field conditions. Am. J. Bot. 1933, 196–203. [Google Scholar] [CrossRef]
- Kurz-Besson, C.; Coûteaux, M.M.; Thiéry, J.M.; Berg, B.; Remacle, J. A comparison of litterbag and direct observation methods of Scots pine needle decomposition measurement. Soil Biol. Biochem. 2005, 37, 2315–2318. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part 2 Chemical and Microbiological Properties; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; Volume 2, pp. 403–430. [Google Scholar]
- Jackson, M.L.; Beltrán, J. Análisis Químico de Suelos (No. 631.41 J335 1982.); Omega: Barcelona, Spain, 1964. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Tyurin, I.V. Analytical procedure for a comparative study of soil humus. Trudy Poch. Inst. Dokuchaev. 1951, 33, 5–21. [Google Scholar]
- Håkansson, I. A method for characterizing the state of compactness of the plough layer. Soil Tillage Res. 1990, 16, 105–120. [Google Scholar] [CrossRef]
- Entry, J.A.; Rose, C.L.; Cromack, K., Jr. Litter decomposition and nutrient release in ectomycorrhizal mat soils of a Douglas fir ecosystem. Soil Biol. Biochem. 1991, 23, 285–290. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1994; Volume 5, pp. 775–833. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Austin, A.T. Differential effects of precipitation on production and decomposition along a rainfall gradient in Hawaii. Ecology 2002, 83, 328–338. [Google Scholar] [CrossRef]
- Austin, A.T.; Vivanco, L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 2006, 442, 555–558. [Google Scholar] [CrossRef]
- Weerakkody, J.; Parkinson, D. Input, accumulation and turnover of organic matter, nitrogen and phosphorus in surface organic layers of an upper montane rainforest in Sri Lanka. Pedobiologia 2006, 50, 377–383. [Google Scholar] [CrossRef]
- Pourhassan, N.; Bruno, S.; Jewell, M.D.; Shipley, B.; Roy, S.; Bellenger, J.P. Phosphorus and micronutrient dynamics during gymnosperm and angiosperm litters decomposition in temperate cold forest from Eastern Canada. Geoderma 2016, 273, 25–31. [Google Scholar] [CrossRef]
- Bahnmann, B.; Mašínová, T.; Halvorsen, R.; Davey, M.L.; Sedlák, P.; Tomšovský, M.; Baldrian, P. Effects of oak, beech and spruce on the distribution and community structure of fungi in litter and soils across a temperate forest. Soil Biol. Biochem. 2018, 119, 162–173. [Google Scholar] [CrossRef]
- Smith, W.K.; Gao, W.E.I.; Steltzer, H.; Wallenstein, M.D.; Tree, R. Moisture availability influences the effect of ultraviolet-B radiation on leaf litter decomposition. Glob. Chang. Biol. 2010, 16, 484–495. [Google Scholar] [CrossRef]
- Marinho, O.A.; Martinelli, L.A.; Duarte-Neto, P.J.; Mazzi, E.A.; King, J.Y. Photodegradation influences litter decomposition rate in a humid tropical ecosystem, Brazil. Sci. Total Environ. 2020, 715, 136601. [Google Scholar] [CrossRef]
- Tejada, M.; Benítez, C. Effects of different organic wastes on soil biochemical properties and yield in an olive grove. Appl. Soil Ecol. 2020, 146, 103371. [Google Scholar] [CrossRef]
- Baker, N.R.; Allison, S.D. Ultraviolet photodegradation facilitates microbial litter decomposition in a Mediterranean climate. Ecology 2015, 96, 1994–2003. [Google Scholar] [CrossRef] [PubMed]
- Hristovski, S.; Berg, B.; Melovski, L. Limitless decomposition in leaf litter of Common beech: Patterns, nutrients’ and heavy metal’s dynamics. Pedobiologia 2014, 57, 131–138. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Decomposition as a process. In Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2008; pp. 11–33. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC (dS m−1) | BD (kg dm−3) | OC (%) | N (%) | C/N | Clay (%) | Silt (%) | Sand (%) | |
|---|---|---|---|---|---|---|---|---|---|
| Avocado | 7.0 (0.1) a | 0.5 (0.1) a | 1.1 (0.0) a | 1.9 (0.6) a | 0.1 (0.0) a | 12.4 (1.7) a | 9.8 (0.3) a | 39.8 (0.7) a | 50.4 (1.0) ab |
| Cherimoya | 8.2 (0.4) b | 0.4 (0.2) a | 1.2 (0.0) a | 0.8 (0.2) b | 0.1 (0.0) ab | 12.8 (1.0) a | 7.1 (0.1) b | 38.6 (1.5) ab | 54.3 (1.4) b |
| Mango | 8.5 (0.2) b | 0.1 (0.0) a | 1.3 (0.0) a | 1.0 (0.3) b | 0.1 (0.0) b | 13.7 (1.1) a | 9.5 (1.2) a | 41.1 (0.6) b | 49.3 (1.6) a |
| Initial Nutrient Composition | Final Nutrient Composition | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AA | CC | MM | GG | AA | CC | MM | GA | GC | GM | |
| Fe (mg/kg) | 109.9 (12.6) a | 123.7 (6.3) a | 201 (11.6) a | 123.7 (26.9) a | 3306.2 (955.2) ab | 5638.1 (2270.6) b | 2517.5 (2188.0) ab | 4686.6 (3095.8) b | 3236.2 (1302.8) ab | 4160.8 (864.3) ab |
| Cu (mg/kg) | 6.6 (0.0) a | 4.7 (0.5) a | 4.7 (0.5) a | 5.8 (0.0) a | 13.4 (4.6) a | 15.9 (8.4) a | 18 (6.0) a | 15.2 (9.4) a | 12.2 (4.4) a | 16.6 (1.9) a |
| Mn (mg/kg) | 49.2 (3.8) cde | 20.4 (2.8) e | 58.6 (2.6) bcde | 38.1 (0.8) de | 101.1 (35.3) abcd | 125.5 (43.7) abc | 132.2 (17.8) ab | 89.6 (35.4) bcde | 79.7 (27.0) bcde | 158.6 (56.5) a |
| Zn (mg/kg) | 13.8 (1.0) a | 10.5 (2.0) a | 11.1 (0.5) a | 14.6 (2.6) ab | 32.5 (7.0) ab | 18.7 (7.7) ab | 14.4 (10.2) a | 33.1 (21.8) ab | 33.5 (11.1) ab | 38.7 (2.9) b |
| P (%) | 0.1 (0.0) bc | 0.1 (0.0) bc | 0.2 (0.0) b | 0.3 (0.0) a | 0.1 (0.0) b | 0.1 (0.0) bc | 0 (0.0) c | 0.1 (0.1) bc | 0.2 (0.0) b | 0.1 (0.0) b |
| K (%) | 0.5 (0.0) ab | 0.3 (0.0) c | 0.5 (0.0) ab | 0.6 (0.0) a | 0.4 (0.1) bc | 0.3 (0.0) c | 0.2 (0.1) c | 0.2 (0.0) c | 0.2 (0.1) c | 0.3 (0.1) c |
| Na (%) | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.1) a | 0.1 (0.0) a | 0.1 (0.0) a | 0.1 (0.0) a |
| Ca (%) | 0.3 (0.0) a | 0.2 (0.0) a | 0.2 (0.0) a | 0.2 (0.0) a | 2 (0.6) ab | 0.7 (0.4) a | 2.0 (1.2) ab | 2.9 (1.4) b | 3.0 (1.3) b | 3.0 (1.5) b |
| Mg (%) | 0.5 (0.0) ab | 0.4 (0.1) abc | 0.0 (0.0) c | 0.0 (0.0) c | 0.5 (0.1) ab | 0.3 (0.1) abc | 0.1 (0.1) bc | 0.5 (0.4) ab | 0.5 (0.2) a | 0.5 (0.1) a |
| C (%) | 47.4 (0.1) a | 47.4 (0.2) a | 45.7 (0.1) ab | 45.6 (0.1) ab | 39.4 (4.8) abc | 35.8 (3.8) bc | 42.1 (3.7) ab | 36.1 (7.8) bc | 38.5 (3.8) abc | 32.6 (5.8) c |
| N (%) | 0.5 (0.1) de | 0.7 (0.1) cde | 0.6 (0.0) cde | 0.8 (0.1) bcde | 1.2 (0.1) abc | 1.1 (0.2) abcd | 0.4 (0.3) e | 1.1 (0.4) abcd | 1.3 (0.3) ab | 1.4 (0.3) a |
| C/N | 90.2 (9.9) ab | 70.3 (8.9) ab | 73.5 (0.9) ab | 59.1 (4.6) ab | 33.1 (4.8) b | 32.7 (8.3) b | 151.5 (112.4) a | 42.3 (24.5) b | 31.1 (9.9) b | 22.9 (1.9) b |
| Own | Garden | ||||||
|---|---|---|---|---|---|---|---|
| k (Years−1) | T50 (Years) | T95 (Years) | k (Years−1) | T50 (Years) | T95 (Years) | ||
| Fe | A | −1.34 | - | - | 0.15 | 4.57 | 19.75 |
| C | 0.20 | 3.48 | 15.03 | 0.20 | 3.47 | 15.00 | |
| M | 0.09 | 7.92 | 34.21 | −0.16 | - | - | |
| Cu | A | −0.02 | - | - | 0.24 | 2.86 | 12.36 |
| C | 0.12 | 5.75 | 24.83 | 0.30 | 2.30 | 9.93 | |
| M | −0.25 | - | - | 0.15 | 4.70 | 20.30 | |
| Mn | A | −0.02 | - | - | 0.06 | 11.12 | 48.05 |
| C | 0.22 | 3.12 | 13.47 | 0.12 | 5.78 | 24.99 | |
| M | −0.27 | - | - | −0.34 | - | - | |
| Zn | A | 0.17 | 4.10 | 17.71 | 0.34 | 2.03 | 8.78 |
| C | 0.36 | 1.93 | 8.34 | 0.23 | 3.05 | 13.17 | |
| M | 0.13 | 5.46 | 23.58 | 0.20 | 3.43 | 14.82 | |
| P | A | 0.33 | 2.08 | 8.98 | 0.38 | 1.82 | 7.85 |
| C | 0.38 | 1.83 | 7.92 | 0.28 | 2.50 | 10.80 | |
| M | 0.61 | 1.14 | 4.95 | 0.45 | 1.55 | 6.69 | |
| K | A | 0.57 | 1.21 | 5.22 | 0.43 | 1.61 | 6.98 |
| C | 0.80 | 0.87 | 3.76 | 0.46 | 1.51 | 6.54 | |
| M | 0.58 | 1.20 | 5.17 | 0.58 | 1.19 | 5.13 | |
| Na | A | 0.53 | 1.30 | 5.61 | 0.14 | 4.97 | 21.49 |
| C | 0.60 | 1.16 | 5.00 | 0.07 | 9.88 | 42.69 | |
| M | 0.43 | 1.60 | 6.93 | 0.11 | 6.56 | 28.35 | |
| Ca | A | −0.13 | - | - | 0.20 | 3.48 | 15.03 |
| C | 0.56 | 1.23 | 5.32 | 0.32 | 2.19 | 9.46 | |
| M | 0.16 | 4.31 | 18.63 | 0.40 | 1.74 | 7.54 | |
| Mg | A | 0.11 | 6.32 | 27.30 | 0.26 | 2.64 | 11.42 |
| C | 0.46 | 1.51 | 6.54 | 0.51 | 1.35 | 5.83 | |
| M | 0.64 | 1.08 | 4.69 | 0.50 | 1.39 | 6.02 | |
| C | A | 0.32 | 2.14 | 9.25 | 0.17 | 4.04 | 17.45 |
| C | 0.30 | 2.33 | 10.07 | 0.38 | 1.80 | 7.80 | |
| M | 0.07 | 9.83 | 42.49 | 0.26 | 2.70 | 11.67 | |
| N | A | −0.04 | - | - | 0.36 | 1.93 | 8.33 |
| C | 0.27 | 2.61 | 11.28 | 0.34 | 2.03 | 8.77 | |
| M | 0.46 | 1.52 | 6.57 | 0.25 | 2.82 | 12.20 | |
| pH | EC (dS m−1) | BD (kg dm−3) | OC (%) | N (%) | C/N | |
|---|---|---|---|---|---|---|
| AA | 6.7 (0.2) a | 0.4 (0.0) a | 0.5 (0.2) ab | 9.8 (0.4) c | 0.7 (0.0) e | 14.5 (0.5) a |
| GA | 7.2 (0.2) bc | 1.3 (0.0) e | 0.4 (0.1) a | 7.8 (1.9) b | 0.5 (0.1) d | 13.5 (2.1) a |
| SA | 7.1 (0.0) bc | 0.5 (0.0) b | 0.8 (0.1) bc | 1.4 (0.5) a | 0.2 (0.0) b | 10.3 (4.0) a |
| CC | 7.8 (0.1) e | 1.9 (0.1) f | 0.7 (0.1) abc | 2.4 (1.0) a | 0.2 (0.0) bc | 11.2 (3.8) a |
| GC | 7.6 (0.1) de | 0.5 (0.0) b | 0.8 (0.1) c | 2.3 (1.3) a | 0.1 (0.0) a | 29.0 (22.9) a |
| SC | 7.0 (0.1) b | 0.7 (0.0) c | 0.7 (0.1) abc | 2.2 (0.5) a | 0.2 (0.0) b | 14.0 (0.6) a |
| MM | 7.3 (0.1) c | 0.6 (0.0) b | 0.7 (0.1) abc | 3.1 (1.6) a | 0.2 (0.0) b | 13.9 (0.8) a |
| GM | 7.4 (0.1) cd | 0.4 (0.0) a | 0.7 (0.1) abc | 2.7 (0.3) a | 0.2 (0.0) bc | 15.0 (2.5) a |
| SM | 7.3 (0.0) c | 0.8 (0.0) d | 0.6 (0.1) abc | 3.2 (0.8) a | 0.3 (0.0) c | 12.9 (3.0) a |
| Glucosidase (µmol PNP g−1 h−1) | Dehydrogenase (µg INTF g−1 h−1) | Urease (µmol N-NH4+ g−1 h−1) | Phosphomonoesterase (µmol PNP g−1 h−1) | |
|---|---|---|---|---|
| AA | 0.3 (0.1) c | 7.4 (1.4) c | 1.2 (0.3) abc | 0.2 (0.0) a |
| GA | 0.3 (0.0) bc | 8.3 (1.1) c | 2.1 (0.7) c | 0.2 (0.1) a |
| SA | 0.1 (0.0) a | 2.8 (1.8) a | 0.5 (0.3) a | 0.1 (0.0) a |
| CC | 0.2 (0.0) bc | 7.0 (1.0) c | 1.5 (0.2) bc | 0.2 (0.1) a |
| GC | 0.2 (0.1) bc | 6.7 (1.0) bc | 0.8 (0.3) ab | 0.2 (0.1) a |
| SC | 0.2 (0.1) ab | 2.9 (1.0) ab | 0.5 (0.2) ab | 0.2 (0.1) a |
| MM | 0.2 (0.0) abc | 4.6 (1.6) abc | 0.9 (0.2) ab | 0.1 (0.0) a |
| GM | 0.2 (0.1) abc | 4.8 (2.0) abc | 1.2 (0.1) ab | 0.1 (0.0) a |
| SM | 0.2 (0.0) bc | 4.7 (1.0) abc | 0.7 (0.3) ab | 0.2 (0.1) a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Martín, M.P.; Ortiz-Bernad, I.; Lallena, A.M.; San-Emeterio, L.M.; Martínez-Cartas, M.L.; Ondoño, E.F. Reuse of Pruning Waste from Subtropical Fruit Trees and Urban Gardens as a Source of Nutrients: Changes in the Physical, Chemical, and Biological Properties of the Soil. Appl. Sci. 2022, 12, 193. https://0-doi-org.brum.beds.ac.uk/10.3390/app12010193
Reyes-Martín MP, Ortiz-Bernad I, Lallena AM, San-Emeterio LM, Martínez-Cartas ML, Ondoño EF. Reuse of Pruning Waste from Subtropical Fruit Trees and Urban Gardens as a Source of Nutrients: Changes in the Physical, Chemical, and Biological Properties of the Soil. Applied Sciences. 2022; 12(1):193. https://0-doi-org.brum.beds.ac.uk/10.3390/app12010193
Chicago/Turabian StyleReyes-Martín, Marino Pedro, Irene Ortiz-Bernad, Antonio M. Lallena, Layla M. San-Emeterio, M. Lourdes Martínez-Cartas, and Emilia Fernández Ondoño. 2022. "Reuse of Pruning Waste from Subtropical Fruit Trees and Urban Gardens as a Source of Nutrients: Changes in the Physical, Chemical, and Biological Properties of the Soil" Applied Sciences 12, no. 1: 193. https://0-doi-org.brum.beds.ac.uk/10.3390/app12010193