
The Fungicide Chlorothalonil Changes the Amphibian Skin Microbiome: A Potential Factor Disrupting a Host Disease-Protective Trait

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tadpole Collection and Maintenance
2.2. Chlorothalonil Exposure and Sampling
2.3. DNA Extraction and 16S rRNA Gene Amplicon Sequencing
2.4. Bioinformatics
2.5. Statistical Analysis in R Environment
3. Results
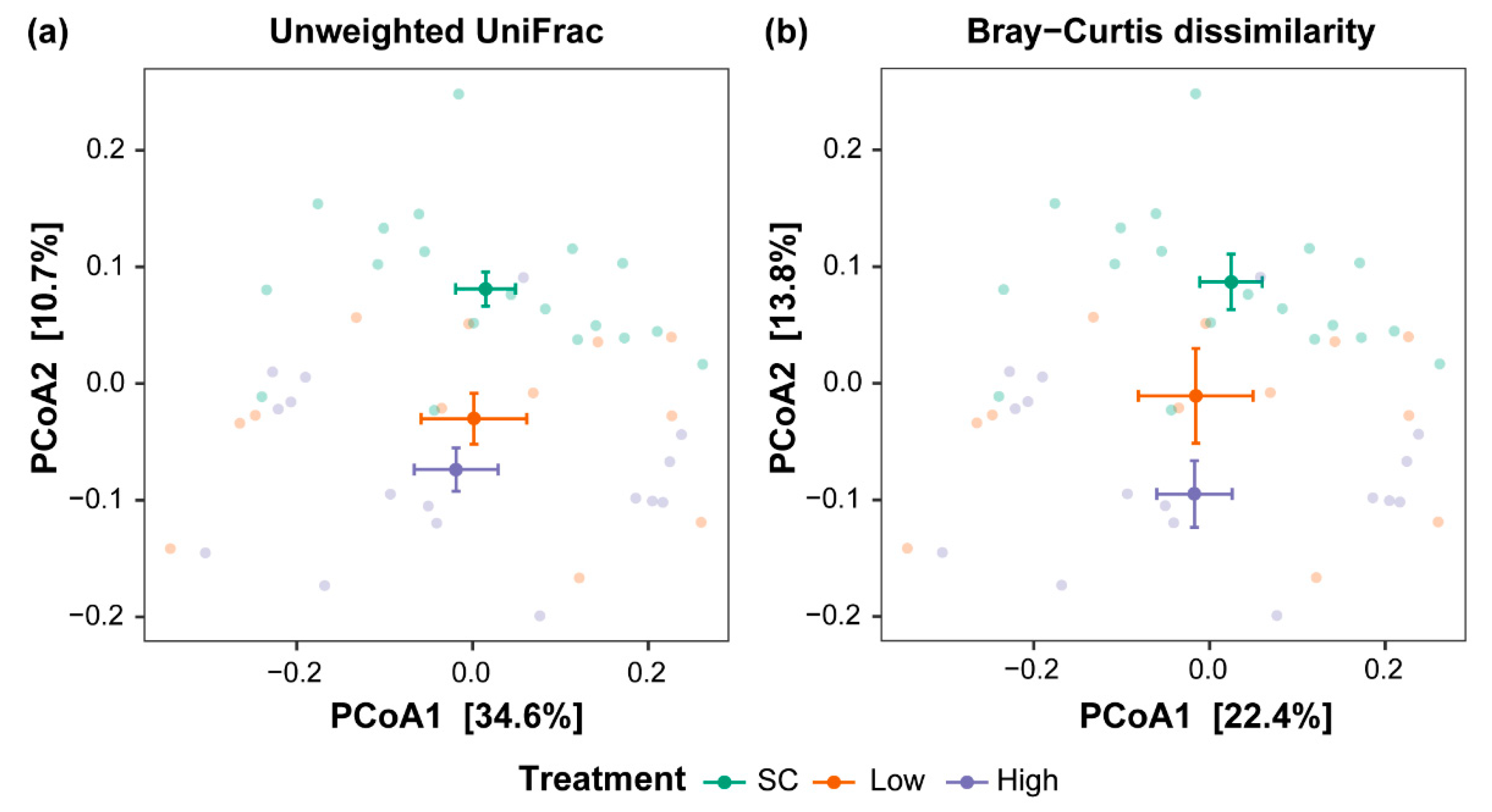
3.1. Chlorothalonil Disturbs the Skin Microbiome Beta Diversity
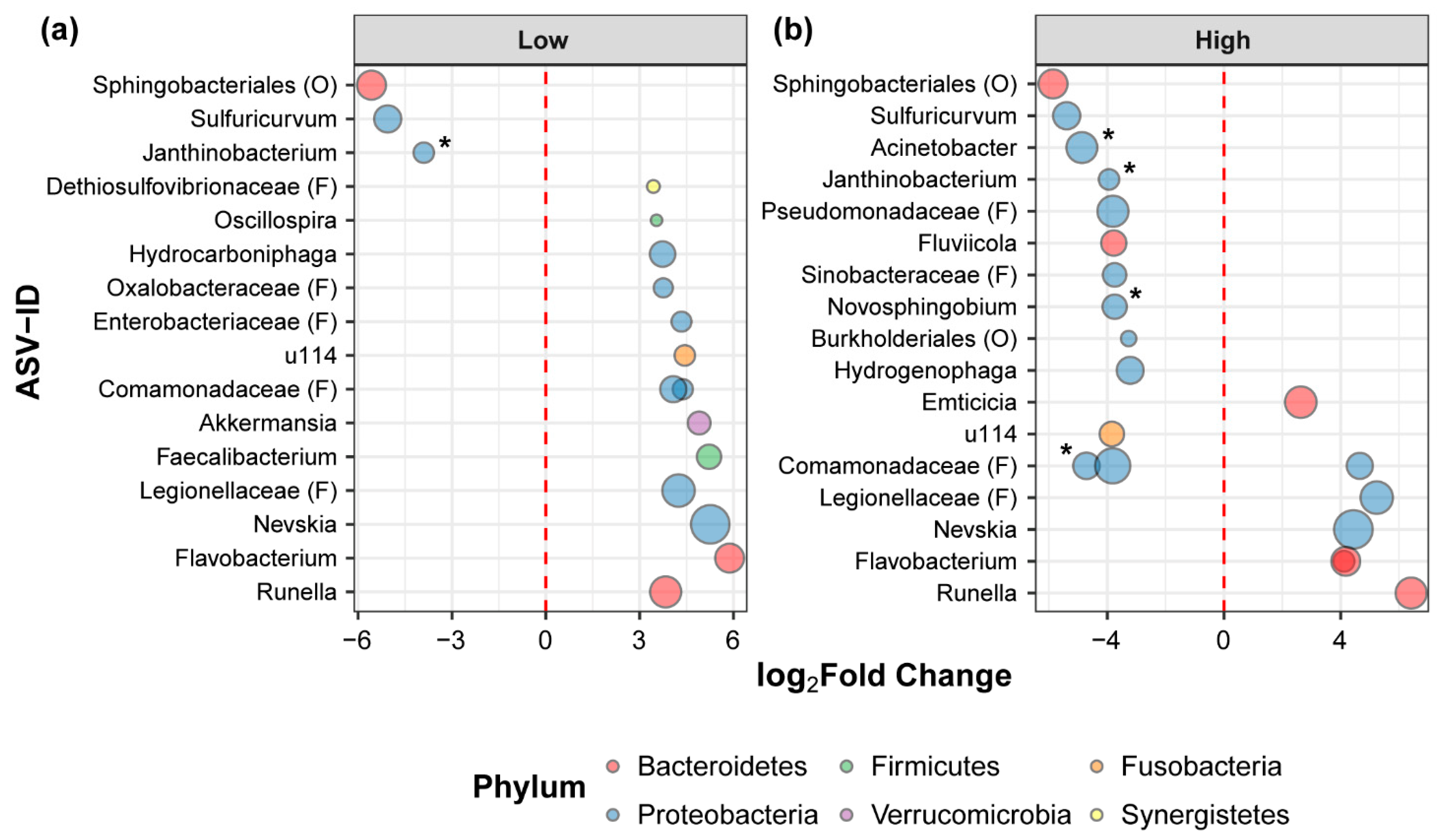
3.2. Chlorothalonil Shifts Relative Abundance of Bacterial Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Densmore, C.L.; Green, D.E. Diseases of amphibians. ILAR J. 2007, 48, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Bower, D.S.; Lips, K.R.; Schwarzkopf, L.; Georges, A.; Clulow, S. Amphibians on the brink. Science 2017, 357, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Scheele, B.C.; Skerratt, L.F.; Grogan, L.F.; Hunter, D.A.; Clemann, N.; McFadden, M.; Newell, D.; Hoskin, C.J.; Gillespie, G.R.; Heard, G.W.; et al. After the epidemic: Ongoing declines, stabilizations and recoveries in amphibians afflicted by chytridiomycosis. Biol. Conserv. 2017, 206, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A. Global amphibian declines, disease, and the ongoing battle between Batrachochytrium fungi and the immune system. Herpetologica 2020, 76, 178–188. [Google Scholar] [CrossRef]
- Briggs, C.J.; Knapp, R.A.; Vredenburg, V.T. Enzootic and epizootic dynamics of the chytrid fungal pathogen of amphibians. Proc. Natl. Acad. Sci. USA 2010, 107, 9695–9700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, A.; der Sluijs, A.S.; Blooi, M.; Bert, W.; Ducatelle, R.; Fisher, M.C.; Woeltjes, A.; Bosman, W.; Chiers, K.; Bossuyt, F.; et al. Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc. Natl. Acad. Sci. USA 2013, 201307356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Micro. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- McLaren, M.R.; Callahan, B.J. Pathogen resistance may be the principal evolutionary advantage provided by the microbiome. Philos. Trans. R. Soc. B. Biol. Sci. 2020, 375, 20190592. [Google Scholar] [CrossRef]
- Rebollar, E.A.; Martínez-Ugalde, E.; Orta, A.H. The amphibian skin microbiome and its protective role against chytridiomycosis. Herpetologica 2020, 76, 167–177. [Google Scholar] [CrossRef]
- Becker, M.H.; Brucker, R.M.; Schwantes, C.R.; Harris, R.N.; Minbiole, K.P.C. The bacterially produced metabolite violacein is associated with survival of amphibians infected with a lethal fungus. Appl. Environ. Microbiol. 2009, 75, 6635–6638. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.N.; Brucker, R.M.; Walke, J.B.; Becker, M.H.; Schwantes, C.R.; Flaherty, D.C.; Lam, B.A.; Woodhams, D.C.; Briggs, C.J.; Vredenburg, V.T.; et al. Skin microbes on frogs prevent morbidity and mortality caused by a lethal skin fungus. ISME J. 2009, 3, 818–824. [Google Scholar] [CrossRef]
- Piovia-Scott, J.; Rejmanek, D.; Woodhams, D.C.; Worth, S.J.; Kenny, H.; McKenzie, V.; Lawler, S.P.; Foley, J.E. Greater species richness of bacterial skin symbionts better suppresses the amphibian fungal pathogen Batrachochytrium dendrobatidis. Microb. Ecol. 2017, 74, 217–226. [Google Scholar] [CrossRef]
- Rebollar, E.A.; Bridges, T.; Hughey, M.C.; Medina, D.; Belden, L.K.; Harris, R.N. Integrating the role of antifungal bacteria into skin symbiotic communities of three neotropical frog species. ISME J. 2019, 13, 1763–1775. [Google Scholar] [CrossRef] [PubMed]
- West, A.G.; Waite, D.W.; Deines, P.; Bourne, D.G.; Digby, A.; McKenzie, V.J.; Taylor, M.W. The microbiome in threatened species conservation. Biol. Conserv. 2019, 229, 85–98. [Google Scholar] [CrossRef]
- Jiménez, R.R.; Sommer, S. The amphibian microbiome: Natural range of variation, pathogenic dysbiosis, and role in conservation. Biodivers. Conserv. 2017, 26, 763–786. [Google Scholar] [CrossRef]
- McCoy, K.A.; Peralta, A.L. Pesticides could alter amphibian skin microbiomes and the effects of Batrachochytrium dendrobatidis. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Paris, L.; Peghaire, E.; Moné, A.; Diogon, M.; Debroas, D.; Delbac, F.; El Alaoui, H. Honeybee gut microbiota dysbiosis in pesticide/parasite co-exposures is mainly induced by Nosema ceranae. J. Invertebr. Pathol. 2020, 172, 107348. [Google Scholar] [CrossRef] [PubMed]
- Daly, G.L.; Lei, Y.D.; Teixeira, C.; Muir, D.C.G.; Castillo, L.E.; Wania, F. Accumulation of current-use pesticides in neotropical montane forests. Environ. Sci. Technol. 2007, 41, 1118–1123. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey bee gut microbiome is altered by in-hive pesticide exposures. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Kohl, K.D.; Cary, T.L.; Karasov, W.H.; Dearing, M.D. Larval exposure to polychlorinated biphenyl 126 (PCB-126) causes persistent alteration of the amphibian gut microbiota. Environ. Toxicol. Chem. 2015, 34, 1113–1118. [Google Scholar] [CrossRef]
- Sigler, W.V.; Turco, R.F. The impact of chlorothalonil application on soil bacterial and fungal populations as assessed by denaturing gradient gel electrophoresis. Appl. Soil Ecol. 2002, 21, 107–118. [Google Scholar] [CrossRef]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. The influence of chlorothalonil on the activity of soil microorganisms and enzymes. Ecotoxicology 2018, 27, 1188–1202. [Google Scholar] [CrossRef]
- Raman, P. Chlorothalonil. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014. [Google Scholar]
- Chaves, A.; Shea, D.; Cope, W.G. Environmental fate of chlorothalonil in a costa rican banana plantation. Chemosphere 2007, 69, 1166–1174. [Google Scholar] [CrossRef]
- Castillo, L.E.; Ruepert, C.; Solis, E. Pesticide residues in the aquatic environment of banana plantation areas in the North Atlantic zone of Costa Rica. Environ. Toxicol. Chem. 2000, 19, 1942–1950. [Google Scholar] [CrossRef]
- Caux, P.-Y.; Kent, R.A.; Fan, G.T.; Stephenson, G.L. Environmental fate and effects of chlorothalonil: A canadian perspective. Crit. Rev. Env. Sci. Technol. 1996, 26, 45–93. [Google Scholar] [CrossRef]
- Tillman, R.W.; Siegel, M.R.; Long, J.W. Mechanism of action and fate of the fungicide chlorothalonil (2,4,5,6-tetrachloroisophthalonitrile) in biological systems: I. Reactions with cells and subcellular components of Saccharomyces pastorianus. Pestic. Biochem. Phys. 1973, 3, 160–167. [Google Scholar] [CrossRef]
- Callicott, K.; Hooper-Bùi, L. An investigation of environmental concentrations of atrazine, chlorothalonil, and fipronil in a Mississippi river-influenced marsh in Southern Louisiana. Microchem. J. 2019, 146, 1241–1248. [Google Scholar] [CrossRef]
- Datta, S.; Hansen, L.; McConnell, L.; Baker, J.; LeNoir, J.; Seiber, J.N. Pesticides and PCB contaminants in fish and tadpoles from the Kaweah River Basin, California. Bull. Environ. Contam. Toxicol. 1998, 60, 829–836. [Google Scholar] [CrossRef]
- Smalling, K.L.; Reeves, R.; Muths, E.; Vandever, M.; Battaglin, W.A.; Hladik, M.L.; Pierce, C.L. Pesticide concentrations in frog tissue and wetland habitats in a landscape dominated by agriculture. Sci. Total Environ. 2015, 502, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Acquaroni, M.; Svartz, G.; Pérez Coll, C. Developmental toxicity asessment of a chlorothalonil-based fungicide in a native amphibian species. Arch. Environ. Contam. Toxicol. 2020. [Google Scholar] [CrossRef]
- Yu, S.; Wages, M.R.; Cobb, G.P.; Maul, J.D. Effects of chlorothalonil on development and growth of amphibian embryos and larvae. Environ. Pollut. 2013, 181, 329–334. [Google Scholar] [CrossRef]
- McMahon, T.A.; Halstead, N.T.; Johnson, S.; Raffel, T.R.; Romansic, J.M.; Crumrine, P.W.; Boughton, R.K.; Martin, L.B.; Rohr, J.R. The fungicide chlorothalonil is nonlinearly associated with corticosterone levels, immunity, and mortality in amphibians. Environ. Health Perspec. 2011, 119, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Méndez, M.; Obando, P.; Pinnock-Branford, M.; Ruepert, C.; Castillo, L.E.; Mena, F.; Alvarado, G. Acute, chronic and biochemical effects of chlorothalonil on Agalychnis callidryas, Isthmohyla pseudopuma and Smilisca baudinii tadpoles. Environ. Sci. Pollut. Res. Int. 2016, 23, 21238–21248. [Google Scholar] [CrossRef]
- Rohr, J.R.; Brown, J.; Battaglin, W.A.; McMahon, T.A.; Relyea, R.A. A pesticide paradox: Fungicides indirectly increase fungal infections. Ecol. Appl. 2017, 27, 2290–2302. [Google Scholar] [CrossRef]
- Savage, J.M. The Amphibians and Reptiles of Costa Rica: A Herpetofauna between Two Continents, between Two Seas; The University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Whitfield, S.M.; Alvarado, G.; Abarca, J.; Zumbado, H.; Zuñiga, I.; Wainwright, M.; Kerby, J. Differential patterns of Batrachochytrium dendrobatidis infection in relict amphibian populations following severe disease-associated declines. Dis. Aquat. Org. 2017, 126, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, R.R.; Alvarado, G.; Estrella, J.; Sommer, S. Moving beyond the host: Unraveling the skin microbiome of endangered costa rican amphibians. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabor, C.R.; Knutie, S.A.; Roznik, E.A.; Rohr, J.R. Are the adverse effects of stressors on amphibians mediated by their effects on stress hormones? Oecologia 2018, 186, 393–404. [Google Scholar] [CrossRef]
- Harrison, X.A.; Price, S.J.; Hopkins, K.; Leung, W.T.M.; Sergeant, C.; Garner, T.W.J. Diversity-stability dynamics of the amphibian skin microbiome and susceptibility to a lethal viral pathogen. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Bataille, A.; Lee-Cruz, L.; Tripathi, B.; Kim, H.; Waldman, B. Microbiome variation across amphibian skin regions: Implications for chytridiomycosis mitigation efforts. Microb. Ecol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Córdoba Gamboa, L.; Solano Diaz, K.; Ruepert, C.; van Wendel de Joode, B. Passive monitoring techniques to evaluate environmental pesticide exposure: Results from the infant’s environmental health study (ISA). Environ. Res. 2020, 184, 109243. [Google Scholar] [CrossRef] [PubMed]
- Grabuski, J. Pesticides in Ontario: A Critical Assessment of Potential Toxicity of Urban Use Products to Wildlife, with Consideration for Endocrine Disruption. Volume 3: Phenoxy Herbicides, Chlorothalonil and Chlorpyrifos; Technical Report Series No. 410; Environment Canada: Dufferin Downsview, ON, Canada.
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Guillaume, B.F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, M.H.; et al. Vegan: Community Ecology Package, R Package Version 2.5-4. 2019. [Google Scholar]
- Del Re, A.C. Compute.es: Compute Effect Sizes, R Package Version 0.2-2. 2013. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Woodhams, D.C.; Alford, R.A.; Antwis, R.E.; Archer, H.; Becker, M.H.; Belden, L.K.; Bell, S.C.; Bletz, M.; Daskin, J.H.; Davis, L.R.; et al. Antifungal isolates database of amphibian skin-associated bacteria and function against emerging fungal pathogens. Ecology 2015, 96, 595. [Google Scholar] [CrossRef] [Green Version]
- Muletz-Wolz, C.R.; Fleischer, R.C.; Lips, K.R. Fungal disease and temperature alter skin microbiome structure in an experimental salamander system. Mol. Ecol. 2019, 28, 2917–2931. [Google Scholar] [CrossRef]
- Zaneveld, J.R.; McMinds, R.; Vega Thurber, R. Stress and stability: Applying the Anna Karenina Principle to animal microbiomes. Nat. Microbiol. 2017, 2, 17121. [Google Scholar] [CrossRef]
- Bletz, M.C.; Kelly, M.; Sabino-Pinto, J.; Bales, E.; Van Praet, S.; Bert, W.; Boyen, F.; Vences, M.; Steinfartz, S.; Pasmans, F.; et al. Disruption of skin microbiota contributes to salamander disease. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180758. [Google Scholar] [CrossRef] [PubMed]
- Krynak, K.L.; Burke, D.J.; Benard, M.F. RodeoTM herbicide negatively affects blanchard’s cricket frogs (Acris blanchardi) survival and alters the skin-associated bacterial community. J. Herpetol. 2017, 51, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Downing, H.F.; DeLorenzo, M.E.; Fulton, M.H.; Scott, G.I.; Madden, C.J.; Kucklick, J.R. Effects of the agricultural pesticides atrazine, chlorothalonil, and endosulfan on South Florida microbial assemblages. Ecotoxicology 2004, 13, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Siddique, T.; Saleem, M.; Arshad, M.; Khalid, A. Chapter 5: Impact of pesticides on soil microbial diversity, enzymes, and biochemical reactions. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2009; Volume 102, pp. 159–200. [Google Scholar]
- Imfeld, G.; Vuilleumier, S. Measuring the effects of pesticides on bacterial communities in soil: A critical review. Eur. J. Soil Biol. 2012, 49, 22–30. [Google Scholar] [CrossRef]
- Oever, J.T.; Netea, M.G. The bacteriome-mycobiome interaction and antifungal host defense. Eur. J. Immunol. 2014, 44, 3182–3191. [Google Scholar] [CrossRef]
- Chin, V.K.; Yong, V.C.; Chong, P.P.; Amin Nordin, S.; Basir, R.; Abdullah, M. Mycobiome in the gut: A multiperspective review. Mediat. Inflamm. 2020, 2020, e9560684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Hamel, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of action and possible impact on nontarget microorganisms. ISRN Ecol. 2011, 2011, e130289. [Google Scholar] [CrossRef] [Green Version]
- Davidson, C.; Benard, M.F.; Shaffer, H.B.; Parker, J.M.; O’Leary, C.; Conlon, J.M.; Rollins-Smith, L.A. Effects of chytrid and carbaryl exposure on survival, growth and skin peptide defenses in foothill yellow-legged frogs. Environ. Sci. Technol. 2007, 41, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.H.; Harris, R.N. Cutaneous bacteria of the redback salamander prevent morbidity associated with a lethal disease. PLoS ONE 2010, 5, e10957. [Google Scholar] [CrossRef] [Green Version]
- Parsons, P.P. Chapter 91—Mammalian toxicokinetics and toxicity of chlorothalonil. In Hayes’ Handbook of Pesticide Toxicology, 3rd ed.; Krieger, R., Ed.; Academic Press: New York, NY, USA, 2010; pp. 1951–1966. ISBN 978-0-12-374367-1. [Google Scholar]
- Penagos, H.G. Contact dermatitis caused by pesticides among banana plantation workers in Panama. Int. J. Occup. Environ. Health 2002, 8, 14–18. [Google Scholar] [CrossRef]
- Penagos, H.; Ruepert, C.; Partanen, T.; Wesseling, C. Pesticide patch test series for the assessment of allergic contact dermatitis among banana plantation workers in Panama. Dermatitis 2004, 15, 137–145. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; et al. Peer review of the pesticide risk assessment of the active substance chlorothalonil. EFSA J. 2018, 16, e05126. [Google Scholar] [CrossRef]
- Baker, B.S. The Role of microorganisms in atopic dermatitis. Clin. Exp. Immunol. 2006, 144, 1–9. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Beta Diversity Metric | Comparison | R2 | p-Value | Adjusted p-Value | Cohen’s d |
|---|---|---|---|---|---|
| Unweighted UniFrac | SC vs. Low | 0.06 | 0.07 | 0.21 | 0.61 (−0.15, 1.39) |
| SC vs. High | 0.09 | 0.01 | 0.02 * | 0.89 (0.17, 1.62) | |
| Low vs. High | 0.04 | 0.30 | 0.90 | 0.23 (−0.55, 1.02) | |
| Bray–Curtis dissimilarity | SC vs. Low | 0.04 | 0.16 | 0.46 | 0.49 (−0.27, 1.26) |
| SC vs. High | 0.10 | 0.001 | 0.003 * | 0.98 (0.25, 1.71) | |
| Low vs. High | 0.05 | 0.12 | 0.35 | 0.37 (−0.42, 1.17) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez, R.R.; Alvarado, G.; Ruepert, C.; Ballestero, E.; Sommer, S. The Fungicide Chlorothalonil Changes the Amphibian Skin Microbiome: A Potential Factor Disrupting a Host Disease-Protective Trait. Appl. Microbiol. 2021, 1, 26-37. https://0-doi-org.brum.beds.ac.uk/10.3390/applmicrobiol1010004
Jiménez RR, Alvarado G, Ruepert C, Ballestero E, Sommer S. The Fungicide Chlorothalonil Changes the Amphibian Skin Microbiome: A Potential Factor Disrupting a Host Disease-Protective Trait. Applied Microbiology. 2021; 1(1):26-37. https://0-doi-org.brum.beds.ac.uk/10.3390/applmicrobiol1010004
Chicago/Turabian StyleJiménez, Randall R., Gilbert Alvarado, Clemens Ruepert, Erick Ballestero, and Simone Sommer. 2021. "The Fungicide Chlorothalonil Changes the Amphibian Skin Microbiome: A Potential Factor Disrupting a Host Disease-Protective Trait" Applied Microbiology 1, no. 1: 26-37. https://0-doi-org.brum.beds.ac.uk/10.3390/applmicrobiol1010004