
A Missing Puzzle in Dissociative Electron Attachment to Biomolecules: The Detection of Radicals


1
Radiation Laboratory, University of Notre Dame, Notre Dame, IN 46556, USA
2
Department of Physics, University of Notre Dame, Notre Dame, IN 46556, USA
Atoms 2021, 9(4), 77; https://0-doi-org.brum.beds.ac.uk/10.3390/atoms9040077
Submission received: 30 August 2021
/
Revised: 1 October 2021
/
Accepted: 3 October 2021
/
Published: 7 October 2021
(This article belongs to the Special Issue Electron Scattering in Gases –from Cross Sections to Plasma Modeling)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Ionizing radiation releases a flood of low-energy electrons that often causes the fragmentation of the molecular species it encounters. Special attention has been paid to the electrons’ contribution to DNA damage via the dissociative electron attachment (DEA) process. Although numerous research groups worldwide have probed these processes in the past, and many significant achievements have been made, some technical challenges have hindered researchers from obtaining a complete picture of DEA. Therefore, this research perspective calls urgently for the implementation of advanced techniques to identify non-charged radicals that form from such a decomposition of gas-phase molecules. Having well-described DEA products offers a promise to benefit society by straddling the boundary between physics, chemistry, and biology, and it brings the tools of atomic and molecular physics to bear on relevant issues of radiation research and medicine.
1. Background and Knowledge Gap
Over the past several decades, significant resources in the atomic and molecular physics community have been directed towards the understanding of collisional processes with biomolecular targets. They are of great importance in radiation research and provide a breadth of potential interests to the life sciences and their applications. However, to translate the outcomes of these molecular processes into a cellular environment, it is necessary to advance our knowledge of scattering processes. A comprehensive understanding of the physical and chemical processes involved at the molecular level remains elusive, even in terms of a single collision between a particle and a target molecule. To achieve a better picture of such physical events, one of the areas that has shown a significant increase in interest, particularly in the electron-scattering community, is a fundamental understanding of electron–biomolecule interactions.
In general, electron–molecule interactions occur in a plethora of physicochemical processes in all types of matter, including living matter, because electrons, together with generated ions and excited molecules, are the most abundant products of ionizing radiation [1]. If the electrons are produced in the condensed phase, they are often referred to as “secondary electrons.” These secondary electrons are created as a result of high-energy photon absorption or during the passage of impinging particles and are due to further inelastic collisions with electrons bound in matter [1]. Moreover, it is commonly accepted that secondary electrons with a lesser amount of energy than the ionization energy of water (~12.5 eV) are “low-energy electrons (LEEs).” A single LEE that interacts with atoms or molecules can determine physical and chemical transformations. For example, it can induce the cleavage of a chemical bond, thus damaging molecules and generating a population of reactive species such as ions and radicals. Subsequently, these reactive species may interact within the medium and lead to form new products or induce further damage.
The pioneering work of Sanche and coworkers in 2000 demonstrated that LEEs can induce severe DNA damage [2]. Since then, a “boom in scientific interest” has emerged and has led to the exploration of the effects of electron scattering from molecules of biological relevance in the gas and condensed phases. During collisions at energies below the molecular target’s ionization threshold, LEEs can be scattered elastically or can lead to rotational, vibrational, and electronic excitation or electron attachment to the target. The excitation can then lead to a neutral dissociation process and the attachment to a dissociative electron attachment (DEA) process (Figure 1). Both quantum processes occur at specific energies, which are referred to as resonances; they correspond to the various energy levels of the transient state and can result in the formation of at least one or more radicals if molecular bond breakage occurs [3].
In Sanche’s pioneering work, resonant structures were also observed in the quantum yields measured in the formation of strand breaks in DNA caused by LEEs as a function of incident electron energy. They were compared to the DEA yields for the two condensed-phase analogues of the DNA constituents, thymine and tetrahydrofurfuryl alcohol, and for water [2]. In the experiments with the DNA analogues, the dissociation patterns observed were the ones in which the molecule lost a hydrogen anion, which is the anion that is then detected with mass spectrometry. The resemblance of both yields, that is, for electron-induced single-strand breaks (SSBs) and double-strand breaks (DSBs) in DNA, and the dissociation of the condensed-phase compounds, prompted the authors to conclude that DNA damage can be initiated by resonant electron attachment to different locations in DNA that is followed by bond dissociation. Already at that time, the DEA process had been intensively studied for a few decades because of ready access to mass-spectrometric tools. Then, the DEA studies have been even more extensive for molecules with a wide range of complexity, from simple diatomic molecules to more complex biomolecular systems in the gas and condensed phases, as well as in clusters [4]. Furthermore, the current state of the art in research on LEE collisions with biomolecules, including DEA studies, was reviewed recently [5]. The authors collected and summarized the main findings in this field, showing the way they are relevant to fundamental and applied studies, highlighting recent experimental and theoretical developments, and attempting to indicate further needs to advance the field. One of the needs in technical development they indicated is the necessity to detect radicals formed during DEA. In the past, only a handful of studies had been published that attempted to investigate radicals attributable to LEE impact, particularly in the condensed phase. Researchers have realized that radical detection would provide a complete description of the dissociation processes that must be employed to determine the mechanism of radiation damage in any biological system, including DNA.
2. Radiation Damage to DNA
Generally, radiation damage to DNA in an aqueous solution can be subdivided into direct damage and indirect damage [6]. The term “direct damage” refers to the ionization of DNA itself or the electron/hole transfer to DNA from its immediate solvent shell. The term “indirect damage” refers to DNA’s attack by the highly reactive species produced by radiation, including free radicals. Free radicals are atomic or molecular species that contain at least one unpaired electron in their structure, and they are stable in vacuo. All reactive species, both radicals and non-radicals (which, contrary to radicals, are closed-shell species), are products of the dissociative excitation and ionization of water and other cellular components present in the surrounding medium, and their possible reaction mechanisms have been studied extensively [6]. However, despite this effort, there remains controversy over the importance and contribution of both direct and indirect effects to DNA damage in vivo. In addition to the complexity of the cellular environment and radiation quality (particle type and energy), there are many factors that contribute to this challenge, such as the different model systems used or different theoretical and experimental approaches taken. Typical radiation chemistry experiments have shown that the doses that are used to damage genetic compounds in the solution are at least one order or even four orders of magnitude greater in these experiments than those that are used to kill mammalian cells [7]. This discrepancy in the level of doses raises a major concern about the precise estimation of radiation effects. Informal estimates of indirect damage to DNA derive from the fact that water, which constitutes 70% to 80% of the cellular mass, absorbs most of the energy of the impinging ionization radiation (~99%) and leads to the production of three types of radicals, the hydroxyl radical (OH), hydrogen (H), and solvated electrons that can attack DNA within a picosecond of diffusion time [6]. Indeed, the early studies on OH formed because of the radiolysis of water showed that indirect effects were responsible for killing approximately 70% of cells [7]. For more than half a century, a vast number of experimental studies had been dedicated to acquiring an understanding of radiation-induced DNA damage by radiolytic species, both radicals and non-radicals, including reactive oxygen and nitrogen species, and providing detailed pathways of their reaction mechanisms [8]. Simultaneously, theoretical attempts were made to describe the action of radiation on DNA through modeling along with the rapid increase in the development of programming languages and computer coding in recent decades attributable to the access to faster and higher-performing technology [1]. Thus, water is the main source of reactive species in an aqueous solution; however, other compounds in a cell located in close proximity to DNA can also provide reactive species, which then induce damage to DNA. For example, proteins wrapped around DNA can play the role of a double-edged sword. On the one hand, they provide physical shielding from ionizing radiation, which protects DNA and decreases damage to it [9]. However, on the other hand, the constituents of a protein can release free radicals upon exposure to ionizing radiation [10], and some model studies have shown that amino acids can increase DNA damage [11].
In addition to studies on radical-induced DNA damage, there have been wide discussions on the role of secondary electrons that are precursors of solvated electrons [12]. The quanta of radiation can generate up to several thousand secondary electrons per event, with kinetic energies as high as half of the energy of the primary quanta down to 0 eV, which includes the formation of LEEs (see Figure 2). LEEs also dominate the secondary electron emission distribution from biomolecular targets exposed to different energies of primary radiation [13,14].
Another physical process, i.e., intermolecular Coulomb decay (ICD) [15], which is one of the well-established mechanisms involved in biological systems, can also produce free electrons [16]. This process can be initiated by photon absorption by a molecule with a higher ionization potential than that of a neighboring molecule in a weakly coupled system. The excitation energy of the molecule is released by energy exchange with the neighboring molecule, which leads to the emission of an electron from the neighboring molecule rather than from the molecule excited initially. The kinetic energy of the emitted electron is typically on the order of several electron volts (<15 eV) [17]. For example, a recent study of the impact of slow, highly-charged ions (Xe40+ ion with 0.6 keV) on graphene reported that up to 80 electrons per ion can be produced in a single event [17]. Moreover, because of the Coulomb explosion of two cationic radicals that are formed during this process, the system decomposes. Therefore, ICD has been proposed to be another important factor in base-pair fragmentation [18] and DNA-strand breaks [19,20]. For example, it has been estimated that the ICD that produces radicals and LEEs may contribute up to 50% of the SSBs at the DNA–water interface during low-energy ionization events.
Undoubtedly, the complexity of the radiation-induced processes in the cellular environment, and the secondary electrons’ contribution to the damage of living matter, including the process of DEA [21,22,23], remains the subject of vigorous debate among physicists and chemists. Beyond the physical and chemical changes induced by ionizing radiation, ionizing radiation has also two general types of adverse biological effects: deterministic and stochastic [24,25]. Deterministic effects cause immediate changes in a cell (e.g., cell death), and their severity occurs only above a certain threshold of radiation dose. Stochastic effects cause long-term changes, and although their probability of occurrence increases with the radiation dose, their severity is the same regardless of how low the dose of radiation was to which the cell was exposed. Hence, any single stochastic event even with a low cross section for its appearance but leading to DNA damage attributable to ionizing radiation can ultimately lead to fatal results in the cell [25]. Thus, all processes that involve LEEs play essential roles in radiation damage to biomolecules and contribute to the alteration and/or fragmentation of biomolecules; they therefore need to be understood and clearly identified [26,27].
3. The State of the Art of Detection Techniques for Neutrals Formed due to LEE Impacts
Generally, the electron attachment to any molecule triggers several dissociation pathways of a transient negative ion (TNI) (see Figure 1). In the DEA process, the resulting fragments are a negative ion and its counterpart. The counterpart can be a single radical that has no charge or several radical fragments, which are neutrals. Most DEA experimental studies employ mass spectrometric instrumentation to detect negative fragments from gas-phase molecules or those desorbed from films [5]. Only a few studies have reported the detection of non-charged fragments because such detections require certain techniques to be incorporated into existing DEA experimental setups or the modification of these setups (which increases the costs of such apparatus significantly). In some cases, changes to these very sensitive set-ups are impossible to make. Despite these challenging technical issues, some results have been reported for neutral desorbed from thin films of nitromethane [28], thymine, bromo-uracil-substituted oligonucleotides [29], modified forms of 11-mercaptoundecanoic acid [30], and DNA [31]. In these cases, electron or photon ionization mass spectrometry was used. In addition, to obtain more information about any species remaining on the irradiated films that could not be desorbed, other in vacuo or ex vacuo analysis techniques were used [27]. However, there has been only one report on detecting radicals that were non-charged species formed from the DEA process of molecules in the gas phase [32]. This study used a double-step ionization technique to identify the neutrals from the dissociation of carbon tetrachloride (CCl4) at close to a 0 eV electron impact. Previously, a similar approach was taken at much higher electron energies than those at which DEA occurs for many plasma-related compounds [33,34,35,36,37,38,39,40]. In these plasma studies, appearance mass spectrometry was used, which is based on the difference between the appearance potential for the ionization of radicals and that for the dissociative ionization of precursor molecules [40]. It has been used widely in the diagnostics of neutral species in plasmas [41]. This technique has been applied successfully in cross-sectional measurements of electron-impact neutral dissociation of gas-phase methane (CH4) into CH3 and CH2 radicals [33,34]; carbon tetrafluoride (CF4) into CF3, CF2, and CF [35]; silicon tetrafluoride (SiF4) into SiF3, SiF, and Si [36]; trifluoromethane (CHF3) into CF3, CF2, CF, CHF2, and CHF [37]; and sulfur hexafluoride (SF6) into SF3, SF2, and SF [38,39]. To date, no studies of biomolecules in the gas phase have been performed, while the neutral products have been studied only for halogenated compounds [32,33,34,35,36,37,38,39,40] or small molecules, particularly hydrocarbons [41]. However, the neutrals formed from these compounds were not produced at energies corresponding to DEA, which occurs below 12 eV, but at much higher energies, above 14 eV and most often at approximately 70 eV, an electron energy region with the highest cross-section for ionization.
In all of these gas-phase studies, neutrals were detected indirectly by ionizing them, and charged products were seen with mass spectrometry. To achieve this, either one electron source or two ionization sources were used. In a study published on DEA to CCl4, the experimental setup was the same as for the conventional gas-phase DEA experiments with one electron source, which was pulsed [32]. In this study, a target molecule was subjected to an electron beam of alternating energies to induce different types of interactions at each step. In each step, the electron beam’s desired combination of energy and pulse frequency was applied. Moreover, the ionic fragments acquired in each step could also be specified with respect to their mass and charge (Figure 3). Thus, in the first step, the electron energy was set at the known resonance peak for the DEA processes and was kept the same throughout the measurement. In contrast, while the electron energy of the second step was scanned, the electron energy was changed at a fixed increment in each iteration of the two steps to determine the energy at which the signal from the expected radical fragment occurred. This energy corresponded to the ionization threshold of the species that was needed to form a cation. The iteration of both steps was repeated until the cation signal appeared. However, rather than repeating these steps using only one electron source, another electron source could be used in the second step to ionize radicals. The choice to use one or two sources would depend on accessible resources, the electron sources’ electronic properties, such as electron beam frequency and energy range, and the experimental set-up’s geometry.
Regardless of the experimental arrangements, the general idea is that the neutral precursor can be identified by comparing the threshold energies necessary to ionize a given species. Typically, the appearance energy (i.e., the minimum energy required for the molecule to dissociate and for its ionization to provide a cationic fragment simultaneously) is much higher than the ionization energy (i.e., the minimum energy required to remove the valence electron) (see Figure 3). Another way to detect radicals using mass spectrometry is by electron attachment that leads to the formation of an anion. In some cases, identifying neutrals by the electron attachment method is preferable when the electron affinity is high for a given radical, as will be mentioned below for the nucleobase moieties [42].
Other ionization techniques can be used to detect radicals, as in the case of desorbed species produced via DEA in the condensed phase, where the photon beam was used to ionize the products [28,31]. In addition to these two ionization methods, i.e., electron ionization and photoionization, technical approaches can be adopted by detecting the emission light from the excited neutral species, such as using Fourier transform infrared spectroscopy or laser-induced fluorescence. These techniques can be complementary to those in which mass spectrometry is involved.
4. Molecular Targets of Opportunity
Because no experimental studies have described all of the products formed by DEA, many possibilities are open for the advancement of our understanding of this process and are necessary to provide its complete picture. Thus far, in attempts to provide detailed fragmentation channels, neutral species are deduced based upon anionic species formed in the experiment and predicted by computational quantum modeling methods, which calculate the most energetically favorable fragmentation channels. This is determined by the reaction enthalpies of possible products; however, this approach disregards the formation of TNI as a precursor species before dissociation.
4.1. Nucleobabses
Beyond the fundamental description of DEA processes, detecting neutral radicals is essential from the perspective of DNA radiation damage, particularly in the case of DSBs by LEEs. It has been shown that the formation of a TNI can lead to SSBs in DNA attributable to the direct interaction of an LEE with the DNA’s sugar–phosphate backbone or due to electron capture by a nucleobase and charge transfer to the DNA backbone [2]. However, it is still unclear how a single LEE with an energy of between 5 and 15 eV can cause a DSB. The molecular description of the mechanism for DSBs can support more complex decomposition pathways [43] that involve direct and indirect effects of LEEs [44,45], and it still requires further investigation. One of the possible pathways is radical formation via DEA in close proximity to the sugar–phosphate backbone. Thus, these neutral radicals can be produced from the DNA itself or the surrounding molecules. For example, one of the most abundant anionic species for all nucleobases (NB) is the anion fragment formed via H loss in the following reaction:
in which is the TNI, H is the hydrogen radical, and is a closed-shell anion of the nucleobase with H loss. Like nucleobases, all studied amino acids also yield H loss upon electron impact as the most abundant dissociation channel [5]. The resulting hydrogen radicals can interact with the DNA strand and cause damage [11], because the H radical is an electrophilic species with a strong preference for attacking electron-rich sites, although at a lower rate than the OH radical [6].
Interestingly, the detection of neutral products of the channel complementary to Reaction 1, in which the hydrogen anion is formed (Reaction 2), can be of great significance in severe damage to DNA.
in which is the TNI, is the closed-shell hydrogen anion, and is a neutral radical of the nucleobase without hydrogen. The resulting radical is an excellent electron acceptor and can receive electrons from the neighboring constituents to form the closed-shell species. The values of adiabatic electron affinity of the neutral radicals of the nucleobases (U—uracil, T—thymine, A—adenine, C—cytosine, G—guanine) without hydrogen follow the order (), which differs somewhat from the order of electron affinity for the closed-shell nucleobases [42]. The formation of these radicals and their further interaction with DNA can potentially lead to a strand break or dimer formation (e.g., the thymine dimer). Although the formation of from nucleobases is well established, no measurements have been performed to detect the neutral in a DEA process.
Therefore, one of the important groups of molecules that would shed light on LEE effects on DNA damage are the nucleobases. Radicals created via DEA to nucleobases can have severe consequences, because the pyrimidine dimer, particularly thymine dimer, is a common result of ultraviolet radiation damage to DNA [6]. DEA to thymine has been studied extensively, and their anionic fragments have been identified and characterized well with respect to their resonant structures [46,47,48,49,50,51,52,53]. Therefore, the initial effort could focus on detecting radicals of nucleobases without hydrogen (Reaction 2). To identify formation, the electron energy should be set at one of the resonant energies, i.e., above 4 eV, where is observed for nucleobases (first-step ionization). A mass spectrum in the positive mode could be recorded at an electron ionization energy of 70 eV (second-step ionization), which for most molecular species has the highest cross-section for ionization. The presence of indicates solely the formation of in Reaction 2, as in earlier electron ionization studies, no was formed from . To confirm these assignments, the complementary negative ion mode can also be performed, in which the electron energy can be scanned at the fixed mass corresponding to . This allows observation of the resonant formation of anions from (second-step ionization). Because the electron affinity for and the probability of the closed-shell anion formation are high [54], the resonant structure in ion yields is expected. A similar methodology can be used to detect other neutral fragments from nucleobases.
4.2. Water
As mentioned above, OH radicals from surrounding water molecules can also cause indirect DNA damage, and it has been suggested that an electron energy above 5 eV causes DSBs that are correlated with the presence of H2O–DNA complexes [45].
Therefore, it is of great importance to detect neutral radicals from DEA to water, which is a simple, yet vital, system for understanding the chemical reactivity that leads to DSBs in DNA. An early attempt to study free radicals from water dissociation was performed at an electron impact of 100 eV [55]. DEA to gas-phase water has been studied extensively and remeasured frequently by several groups, and cross sections for DEA have been compiled recently [56]. Three anionic fragments, i.e., H‾, OH‾, and O‾, produced from intact water molecules were observed experimentally:
To study neutral fragments (stated in parentheses in Reactions 3–5) formed through DEA to water, the electron energy should be set at the resonance energy where a specific negative ion was observed (first-step ionization), as shown in Figure 4. Then, the mass spectrometer should be set at the mass corresponding to the counterpart neutral fragment of the anion observed while the electron energy is scanned either in the positive or negative modes of the mass spectrometer (second-step ionization). The positive mode (cation detection) can determine the threshold energy for the neutral fragments’ ionization. One must keep in mind that the energy scans can include the contribution from water molecules present in the chamber (hereafter denoted as background H2O) while performing the second-step ionization. However, the appearance energies and anionic resonances of cations and anions produced from background H2O, respectively, have been documented well [56,57], and they differ from those expected from neutral fragments. Therefore, the signals obtained from background H2O can be subtracted. The scheme of the experimental procedure is presented in Figure 4 for Reaction 3. For example, the appearance energy for OH+ from gas-phase water is 18.2 eV, whereas the ionization of the OH fragment is approximately 5 eV lower [57]. Therefore, by scanning the ion yield for mass 17 as a function of electron energy, one can deduce whether the cation observed derives from an intact water molecule or the counter-product of the DEA process. The negative mode (anion detection) can be used to observe the resonant formation of an anion from neutral fragments. It is also expected that the anion yield features formed from gas-phase water differ from those fragments, because the electron affinity for an intact molecule differs from that of neutral fragments. It is important to note that there are reports of radical detection from water dissociation attributable to LEE-induced neutral dissociation, but not via DEA [56].
5. Conclusions
A continuing quest to understand fundamental phenomena induced by ionizing radiation, particularly LEEs, which are invariable primary products in any irradiated matter, is still the ongoing focus of the radiation research community. Although these free or quasi-free electrons do not travel very far because of their many inelastic collisions, and because they become thermalized within approximately 1 picosecond, they play an essential role in the dissociation of molecules along their way and in the production of longer-lived species such as radicals [22,58,59].
Despite the extensive research by atomic and molecular physics groups on LEE interactions with gas-phase and condensed-phased biomolecules, some fundamental aspects remain unexamined at the molecular level. One of these is detecting radicals from the DEA process, which is a missing puzzle for obtaining a complete picture of this process. Because stable radicals formed in this process are non-charged species, it is not possible to detect them directly by mass spectrometry, which is used commonly to study DEA. Therefore, modified mass spectrometric techniques or other currently available technical advances need to be used to reveal detailed fragmentation patterns. Obtaining fully described patterns can be incorporated into the database of electronic properties of biomolecules, which are invaluable to build accurate theoretical and computational models of radiation effects [5,60]. This can reveal new mechanistic information on DNA damage during irradiation and can be used to plan radiotherapy treatment. The discovery of all products formed via DEA will transform our fundamental understanding of LEE interactions with biomolecular systems and has the great potential to yield physical information on the chemistry and biology of radiation-induced damage of living cells. Thus, a thorough understanding of this basic and significant collisional process may lead more broadly to enhanced medical applications in the fields of radiotherapy, radiodiagnostics, and radiation protection.
Funding
The work was supported by the U.S. Department of Energy Office of Science, Office of Basic Energy Sciences under Award Number DE-FC02-04ER15533. This is contribution number NDRL 5335 from the Notre Dame Radiation Laboratory.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Plante, I. A review of simulation codes and approaches for radiation chemistry. Phys. Med. Biol. 2021, 66, 03TR02. [Google Scholar] [CrossRef]
- Boudaïffa, B.; Cloutier, P.; Hunting, D.; Huels, M.A.; Sanche, L. Resonant Formation of DNA Strand Breaks by Low-Energy (3 to 20 eV) Electrons. Science 2000, 287, 1658–1660. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, D.S.; Rescigno, T.N. Breaking up is hard to do. Nat. Phys. 2018, 14, 109–110. [Google Scholar] [CrossRef]
- Fabrikant, I.I.; Eden, S.; Mason, N.J.; Fedor, J. Chapter Nine—Recent Progress in Dissociative Electron Attachment: From Diatomics to Biomolecules. In Advances in Atomic, Molecular, and Optical Physics; Arimondo, E., Lin, C.C., Yelin, S.F., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 66, pp. 545–657. [Google Scholar]
- Gorfinkiel, J.D.; Ptasinska, S. Electron scattering from molecules and molecular aggregates of biological relevance. J. Phys. B At. Mol. Opt. Phys. 2017, 50, 182001. [Google Scholar] [CrossRef] [Green Version]
- Sonntage, C.V. Free-Radical-Induced DNA Damage and Its Repair, 1st ed.; Springer: Berlin, Germany, 2006; p. 523. [Google Scholar]
- Ward, J.F. Radiolytic damage to genetic material. J. Chem. Educ. 1981, 58, 135–139. [Google Scholar] [CrossRef]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Radic. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Spotheim-Maurizot, M.; Davídková, M. Radiation damage to DNA in DNA–protein complexes. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2011, 711, 41–48. [Google Scholar] [CrossRef]
- Sagstuen, E.; Sanderud, A.; Hole, E.O. The Solid-State Radiation Chemistry of Simple Amino Acids, Revisited. Radiat. Res. 2004, 162, 112–119. [Google Scholar] [CrossRef]
- Ptasińska, S.; Li, Z.; Mason, N.J.; Sanche, L. Damage to amino acid–nucleotide pairs induced by 1 eV electrons. Phys. Chem. Chem. Phys. 2010, 12, 9367–9372. [Google Scholar] [CrossRef]
- Alizadeh, E.; Sanche, L. Precursors of solvated electrons in radiobiological physics and chemistry. Chem. Rev. 2012, 112, 5578–5602. [Google Scholar] [CrossRef]
- Moretto-Capelle, P.; Le Padellec, A. Electron spectroscopy in proton collisions with dry gas-phase uracil base. Phys. Rev. A 2006, 74, 062705. [Google Scholar] [CrossRef]
- Padellec, A.L.; Moretto-Capelle, P.; Richard-Viard, M.; Champeaux, J.; Cafarelli, P. Ionization and fragmentation of DNA, RNA bases induced by proton impact. J. Phys. Conf. Ser. 2008, 101, 012007. [Google Scholar] [CrossRef]
- Jahnke, T.; Hergenhahn, U.; Winter, B.; Dörner, R.; Frühling, U.; Demekhin, P.V.; Gokhberg, K.; Cederbaum, L.S.; Ehresmann, A.; Knie, A.; et al. Interatomic and Intermolecular Coulombic Decay. Chem. Rev. 2020, 120, 11295–11369. [Google Scholar] [CrossRef] [PubMed]
- Harbach, P.H.P.; Schneider, M.; Faraji, S.; Dreuw, A. Intermolecular Coulombic Decay in Biology: The Initial Electron Detachment from FADH− in DNA Photolyases. J. Phys. Chem. Lett. 2013, 4, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Schwestka, J.; Niggas, A.; Creutzburg, S.; Kozubek, R.; Heller, R.; Schleberger, M.; Wilhelm, R.A.; Aumayr, F. Charge-Exchange-Driven Low-Energy Electron Splash Induced by Heavy Ion Impact on Condensed Matter. J. Phys. Chem. Lett. 2019, 10, 4805–4811. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Guo, D.; Ma, X.; Zhu, X.; Feng, W.; Yan, S.; Zhao, D.; Gao, Y.; Zhang, S.; Ren, X.; et al. Damaging Intermolecular Energy and Proton Transfer Processes in Alpha-Particle-Irradiated Hydrogen-Bonded Systems. Angew. Chem. Int. Ed. 2018, 57, 17023–17027. [Google Scholar] [CrossRef] [PubMed]
- Grieves, G.A.; Orlando, T.M. Intermolecular Coulomb Decay at Weakly Coupled Heterogeneous Interfaces. Phys. Rev. Lett. 2011, 107, 016104. [Google Scholar] [CrossRef] [Green Version]
- Gokhberg, K.; Kolorenč, P.; Kuleff, A.I.; Cederbaum, L.S. Site- and energy-selective slow-electron production through intermolecular Coulombic decay. Nature 2014, 505, 661–663. [Google Scholar] [CrossRef] [Green Version]
- Kohanoff, J.; McAllister, M.; Tribello, G.A.; Gu, B. Interactions between low energy electrons and DNA: A perspective from first-principles simulations. J. Phys. Condens. Matter 2017, 29, 383001. [Google Scholar] [CrossRef]
- Ma, J.; Kumar, A.; Muroya, Y.; Yamashita, S.; Sakurai, T.; Denisov, S.A.; Sevilla, M.D.; Adhikary, A.; Seki, S.; Mostafavi, M. Observation of dissociative quasi-free electron attachment to nucleoside via excited anion radical in solution. Nat. Commun. 2019, 10, 102. [Google Scholar] [CrossRef]
- Dong, Y.; Liao, H.; Gao, Y.; Cloutier, P.; Zheng, Y.; Sanche, L. Early Events in Radiobiology: Isolated and Cluster DNA Damage Induced by Initial Cations and Nonionizing Secondary Electrons. J. Phys. Chem. Lett. 2021, 12, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, N.W.; Parrish, J.M.; Sheha, E.D.; Singh, K. Intraoperative risks of radiation exposure for the surgeon and patient. Ann. Transl. Med. 2021, 9, 84. [Google Scholar] [CrossRef]
- Gauduel, Y.; Glinec, Y.; Malka, V. Femtoradical events in aqueous molecular environments: The tenuous borderline between direct and indirect radiation damages. J. Phys. Conf. Ser. 2008, 101, 012004. [Google Scholar] [CrossRef]
- Gao, Y.; Zheng, Y.; Sanche, L. Low-Energy Electron Damage to Condensed-Phase DNA and Its Constituents. Int. J. Mol. Sci. 2021, 22, 7879. [Google Scholar] [CrossRef]
- Bazin, M.; Ptasinska, S.; Bass, A.D.; Sanche, L.; Burean, E.; Swiderek, P. Electron induced dissociation in the condensed-phase nitromethane: II. Desorption of neutral fragments. J. Phys. Condens. Matter 2010, 22, 084003. [Google Scholar] [CrossRef]
- Abdoul-Carime, H.; Dugal, P.C.; Sanche, L. Damage induced by 1-30 eV electrons on thymine- and bromouracil-substituted oligonucleotides. Radiat. Res. 2000, 153, 23–28. [Google Scholar] [CrossRef]
- Houplin, J.; Amiaud, L.; Humblot, V.; Martin, I.; Matar, E.; Azria, R.; Pradier, C.-M.; Lafosse, A. Selective terminal function modification of SAMs driven by low-energy electrons (0–15 eV). Phys. Chem. Chem. Phys. 2013, 15, 7220–7227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Aleksandrov, A.; Orlando, T.M. Probing low-energy electron induced DNA damage using single photon ionization mass spectrometry. Int. J. Mass Spectrom. 2008, 277, 314–320. [Google Scholar] [CrossRef]
- Li, Z.; Milosavljević, A.R.; Carmichael, I.; Ptasinska, S. Characterization of Neutral Radicals from a Dissociative Electron Attachment Process. Phys. Rev. Lett. 2017, 119, 053402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Toyoda, H.; Sugai, H. Electron-Impact Dissociation of Methane into CH3 and CH2 Radicals I. Relative Cross Sections. Jpn. J. Appl. Phys. 1991, 30, 2908. [Google Scholar] [CrossRef]
- Nakano, T.; Toyoda, H.; Sugai, H. Electron-Impact Dissociation of Methane into CH3 and CH2 Radicals II. Absolute Cross Sections. Jpn. J. Appl. Phys. 1991, 30, 2912. [Google Scholar] [CrossRef]
- Nakano, T.; Sugai, H. Partial Cross Sections for Electron Impact Dissociation of CF4 into Neutral Radicals. Jpn. J. Appl. Phys. 1992, 31, 2919. [Google Scholar] [CrossRef]
- Nakano, T.; Sugai, H. Cross section measurements for electron-impact dissociation of SiF4 into neutral radicals. J. Phys. D Appl. Phys. 1993, 26, 1909. [Google Scholar] [CrossRef]
- Fisher, E.R.; Kickel, B.L.; Armentrout, P.B. Collision-induced dissociation and charge transfer reactions of SF+x (x = 1 − 5): Thermochemistry of sulfur fluoride ions and neutrals. J. Chem. Phys. 1992, 97, 4859. [Google Scholar] [CrossRef]
- Iio, M.; Goto, M.; Sugai, H. Relative Cross Sections for Electron—Impact Dissociation of SF6 into SFx (x = 1 − 3) Neutral Radicals. Contrib. Plasma Phys. 1995, 35, 405–413. [Google Scholar] [CrossRef]
- Christophorou, L.G.; Olthoff, J.K. Electron Interactions With SF6. J. Phys. Chem. Ref. Data 2000, 29, 267. [Google Scholar] [CrossRef] [Green Version]
- Sugai, H.; Toyoda, H. Appearance mass spectrometry of neutral radicals in radio frequency plasmas. J. Vac. Sci. Technol. A 1992, 10, 1193. [Google Scholar] [CrossRef]
- Benedikt, J.; Kersten, H.; Piel, A. Foundations of measurement of electrons, ions and species fluxes toward surfaces in low-temperature plasmas. Plasma Sources Sci. Technol. 2021, 30, 033001. [Google Scholar] [CrossRef]
- Berdys, J.; Anusiewicz, I.; Skurski, P.; Simons, J. Damage to Model DNA Fragments from Very Low-Energy (<1 eV) Electrons. J. Am. Chem. Soc. 2004, 126, 6441–6447. [Google Scholar] [CrossRef]
- Hahn, M.B.; Meyer, S.; Schröter, M.-A.; Seitz, H.; Kunte, H.-J.; Solomun, T.; Sturm, H. Direct electron irradiation of DNA in a fully aqueous environment. Damage determination in combination with Monte Carlo simulations. Phys. Chem. Chem. Phys. 2017, 19, 1798–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh, E.; Orlando, T.M.; Sanche, L. Biomolecular Damage Induced by Ionizing Radiation: The Direct and Indirect Effects of Low-Energy Electrons on DNA. Annu. Rev. Phys. Chem. 2015, 66, 379–398. [Google Scholar] [CrossRef] [PubMed]
- Orlando, T.M.; Oh, D.; Chen, Y.; Aleksandrov, A.B. Low-energy electron diffraction and induced damage in hydrated DNA. J. Chem. Phys. 2008, 128, 195102. [Google Scholar] [CrossRef] [PubMed]
- Denifl, S.; Sulzer, P.; Zappa, F.; Moser, S.; Kräutler, B.; Echt, O.; Bohme, D.K.; Märk, T.D.; Scheier, P. Isotope effects in dissociative electron attachment to the DNA base thymine. Int. J. Mass Spectrom. 2008, 277, 296–299. [Google Scholar] [CrossRef]
- Kopyra, J.; Koenig-Lehmann, C.; Illenberger, E. On the absolute value for the cross-section of dissociative electron attachment (DEA) to the DNA base thymine. Int. J. Mass Spectrom. 2009, 281, 89–91. [Google Scholar] [CrossRef]
- Ptasinska, S.; Denifl, S.; Scheier, P.; Illenberger, E.; Märk, T.D. Bond- and Site-Selective Loss of H Atoms from Nucleobases by Very-Low-Energy Electrons (<3 eV). Angew. Chem. Int. Ed. 2005, 44, 6941–6943. [Google Scholar] [CrossRef]
- Ptasińska, S.; Denifl, S.; Grill, V.; Märk, T.D.; Scheier, P.; Gohlke, S.; Huels, M.A.; Illenberger, E. Bond-Selective H− Ion Abstraction from Thymine. Angew. Chem. Int. Ed. 2005, 44, 1647–1650. [Google Scholar] [CrossRef]
- Chernyshova, I.V.; Kontrosh, E.E.; Shpenik, O.B. Collisions of Slow Electrons with Thymine Molecules. Opt. Spectrosc. 2018, 125, 845–852. [Google Scholar] [CrossRef]
- Ptasińska, S.; Denifl, S.; Grill, V.; Märk, T.D.; Illenberger, E.; Scheier, P. Bond- and Site-Selective Loss of H− from Pyrimidine Bases. Phys. Rev. Lett. 2005, 95, 093201. [Google Scholar] [CrossRef] [Green Version]
- Ptasińska, S.; Denifl, S.; Mróz, B.; Probst, M.; Grill, V.; Illenberger, E.; Scheier, P.; Märk, T.D. Bond selective dissociative electron attachment to thymine. J. Chem. Phys. 2005, 123, 124302. [Google Scholar] [CrossRef]
- Burrow, P.D.; Gallup, G.A.; Scheer, A.M.; Denifl, S.; Ptasinska, S.; Märk, T.; Scheier, P. Vibrational Feshbach resonances in uracil and thymine. J. Chem. Phys. 2006, 124, 124310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Leszczynski, J.; Schaefer, H.F. Interactions of Electrons with Bare and Hydrated Biomolecules: From Nucleic Acid Bases to DNA Segments. Chem. Rev. 2012, 112, 5603–5640. [Google Scholar] [CrossRef] [PubMed]
- Melton, C.E. Radiolysis of water vapor in a wide range radiolysis source of a mass spectrometer. I. Individual and total cross sections for the production of positive ions, negative ions, and free radicals by electrons. J. Phys. Chem. 1970, 74, 582–587. [Google Scholar] [CrossRef]
- Song, M.-Y.; Cho, H.; Karwasz, G.P.; Kokoouline, V.; Nakamura, Y.; Tennyson, J.; Faure, A.; Mason, N.J.; Itikawa, Y. Cross Sections for Electron Collisions with H2O. J. Phys. Chem. Ref. Data 2021, 50, 023103. [Google Scholar] [CrossRef]
- NIST Chemistry WebBook. Available online: http://webbook.nist.gov (accessed on 30 August 2021).
- Arumainayagam, C.R.; Lee, H.L.; Nelson, R.B.; Haines, D.R.; Gunawardane, R.P. Low-energy electron-induced reactions in condensed matter. Surf. Sci. Rep. 2010, 65, 1–44. [Google Scholar] [CrossRef]
- Tsuchida, H.; Kai, T.; Kitajima, K.; Matsuya, Y.; Majima, T.; Saito, M. Relation between biomolecular dissociation and energy of secondary electrons generated in liquid water by fast heavy ions. Eur. Phys. J. D 2020, 74, 212. [Google Scholar] [CrossRef]
- Mason, N.J. Electron Induced Processing; Applications and Data Needs. AIP Conf. Proc. 2007, 901, 74–84. [Google Scholar] [CrossRef]
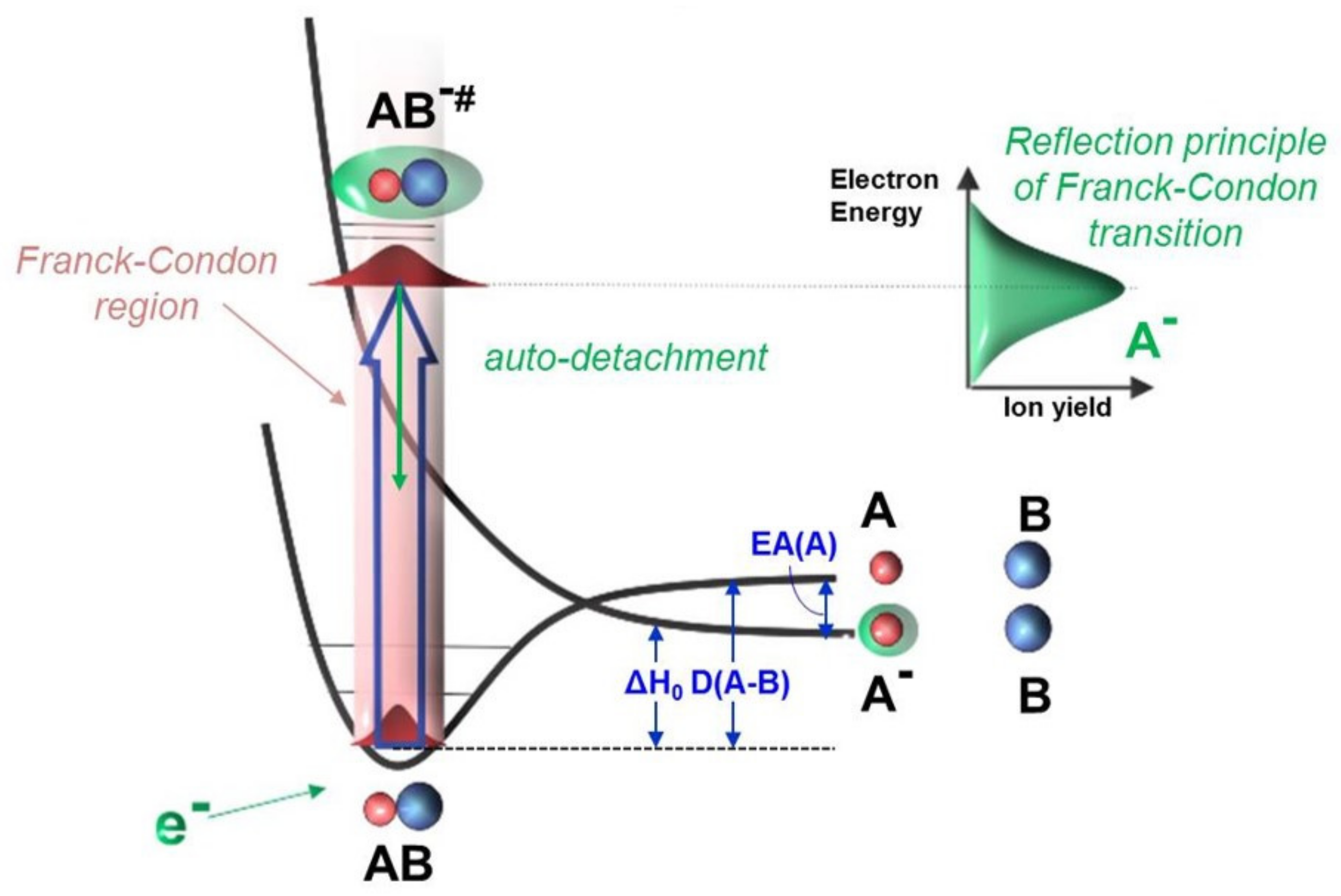
Figure 1.
Schematic diagram for the DEA process: in which AB is a molecular target, is a vibrationally excited transient negative ion (TNI), is a negative ion product, B is a neutral radical formed, is the reaction enthalpy, EA(A) is the electron affinity of neutral fragment A, and D(A-B) is the bond (A-B) dissociation energy. First, the TNI is formed through a vertical transition from the electronic ground state of the neutral molecule to the potential energy surface of the negative ion within the Franck–Condon region. Then, primarily, the TNI will decay into the neutral molecule by electron auto-detachment. If the lifetime of TNI is long enough that the doorway of dissociation becomes accessible, the system stabilizes by breaking down into an anion fragment () and a neutral fragment (B) or multiple neutral fragments. Experimentally, the ion yield of is monitored by scanning the electron energy and is shown as a resonant peak that reflects the initial Franck–Condon transition.
Figure 1.
Schematic diagram for the DEA process: in which AB is a molecular target, is a vibrationally excited transient negative ion (TNI), is a negative ion product, B is a neutral radical formed, is the reaction enthalpy, EA(A) is the electron affinity of neutral fragment A, and D(A-B) is the bond (A-B) dissociation energy. First, the TNI is formed through a vertical transition from the electronic ground state of the neutral molecule to the potential energy surface of the negative ion within the Franck–Condon region. Then, primarily, the TNI will decay into the neutral molecule by electron auto-detachment. If the lifetime of TNI is long enough that the doorway of dissociation becomes accessible, the system stabilizes by breaking down into an anion fragment () and a neutral fragment (B) or multiple neutral fragments. Experimentally, the ion yield of is monitored by scanning the electron energy and is shown as a resonant peak that reflects the initial Franck–Condon transition.

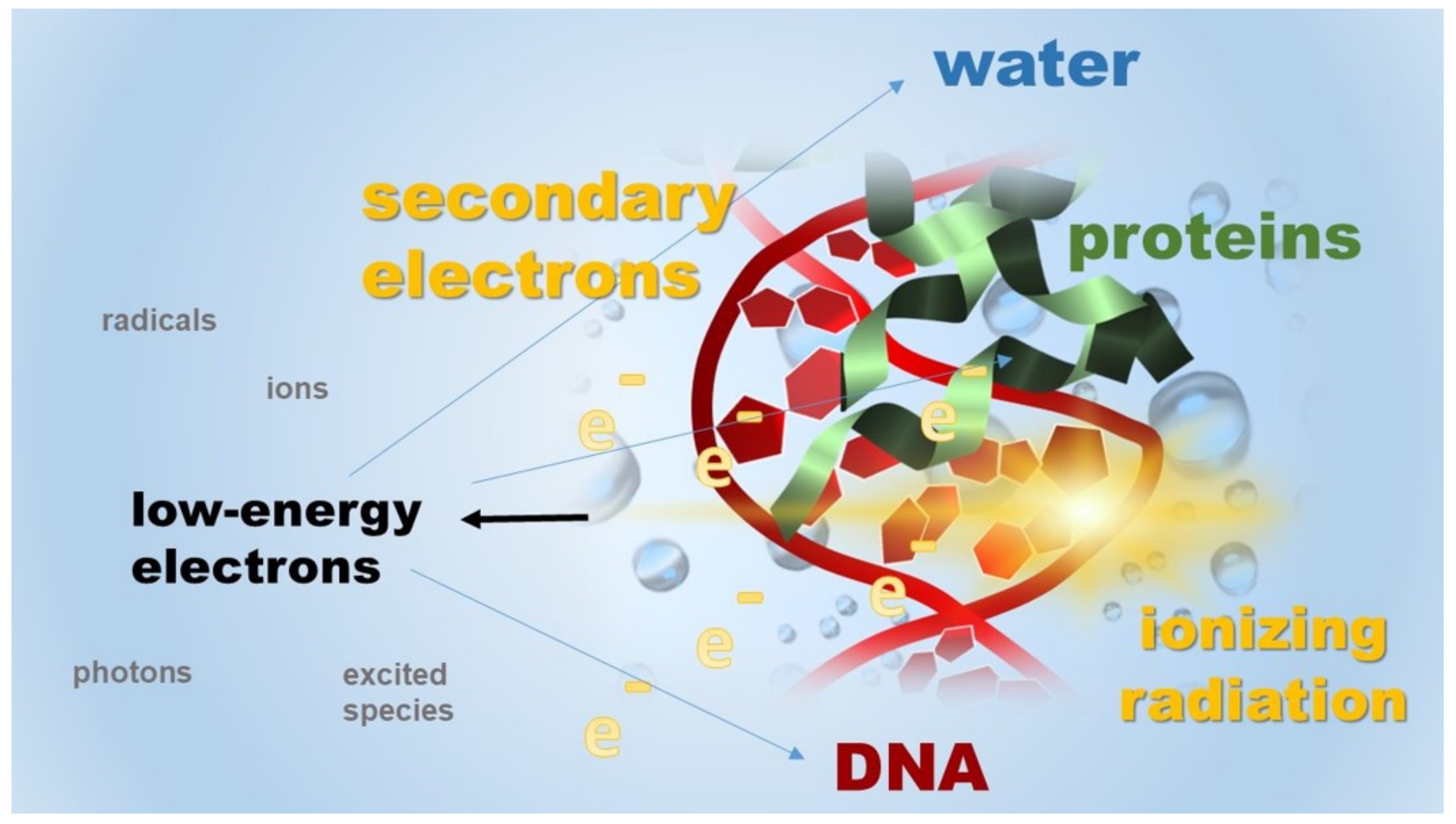
Figure 2.
An illustrative representation of LEEs produced from interactions between high-energy radiation and cellular constituents. The inelastic collisions of these LEEs with surrounding molecules cause their damage, e.g., DNA damage, and produce distinct energetic species, i.e., excited species, ions, and radicals that are further driving forces in a wide variety of radiation-induced chemical reactions.
Figure 2.
An illustrative representation of LEEs produced from interactions between high-energy radiation and cellular constituents. The inelastic collisions of these LEEs with surrounding molecules cause their damage, e.g., DNA damage, and produce distinct energetic species, i.e., excited species, ions, and radicals that are further driving forces in a wide variety of radiation-induced chemical reactions.

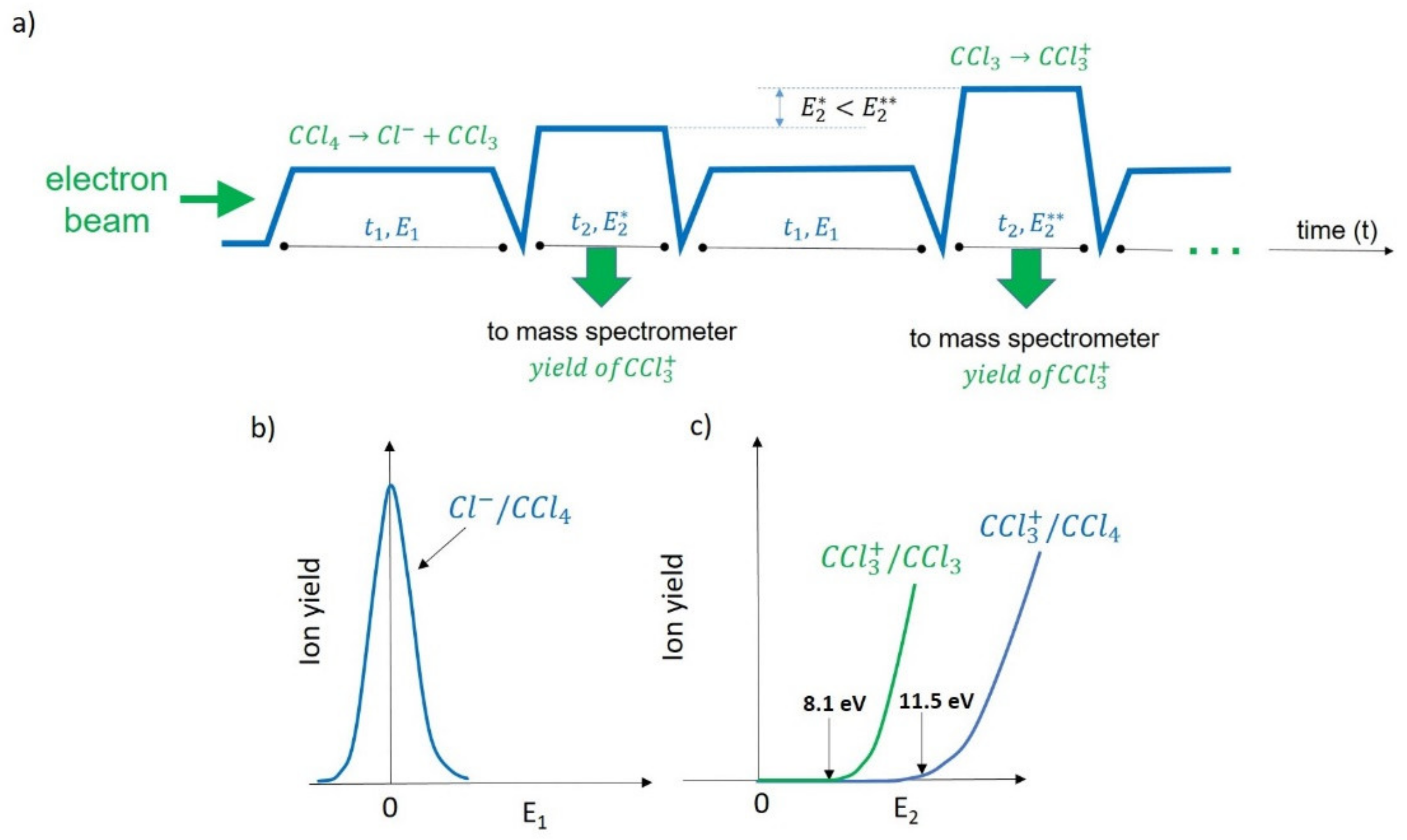
Figure 3.
(a) A time flow of the experiment for the neutral products from DEA to CCl4 using a stepwise electron spectroscopy technique [32]; (b) the Cl− ion formation by DEA to CCl4; (c) the ionization threshold for the ion from the radical and the appearance energy of the fragment due to the dissociation of upon the electron impact. In the first step of the acquisition scheme (a), the electron beam energy E1 was set at approximately 0 eV, and the electrons interact with the molecular beam in the collision region during time t1. The Cl− ions (formed by DEA to CCl4 (b)) were acquired during the same period t1. Then, the electron energy was increased to E2, which was slightly below the ionization energy of . The energy E2 continued to be increased in each iteration until the ion signal for was clearly detectable. During the second step (t2), the electron beam of energy E2 interacts with the mix of CCl4 and the products that resulted from the interaction in the first step, i.e., Cl− and . The mass spectrometer was set to detect the ion yield of during the second step t2. Because there was a mix of CCl4, Cl−, and during the second step inside the electron–molecule interaction region, the acquired could be produced by both the electron impact fragmentation of CCl4 molecules (from the target beam) and the electron impact ionization of the fragment formed during time t1. However, there was a difference between the appearance energy of from CCl4 and the ionization energy of the (c). In other words, the appearance energy of was over 3 eV higher than the ionization energy of . During the experiment, the electron energies in the steps, E1 and E2, were controllable. The same procedure can be applied to detect or exclude other possible neutrals, e.g., CCl, CCl2, and so on, produced in the DEA process by properly adjusting the E2 value. In addition, the period of t1 and t2 is controllable as well. For example, it was set such that t1 and t2 were equal to 1 s and 0.1 s, respectively, in the experiment published for the detection from DEA to CCl4 [32].
Figure 3.
(a) A time flow of the experiment for the neutral products from DEA to CCl4 using a stepwise electron spectroscopy technique [32]; (b) the Cl− ion formation by DEA to CCl4; (c) the ionization threshold for the ion from the radical and the appearance energy of the fragment due to the dissociation of upon the electron impact. In the first step of the acquisition scheme (a), the electron beam energy E1 was set at approximately 0 eV, and the electrons interact with the molecular beam in the collision region during time t1. The Cl− ions (formed by DEA to CCl4 (b)) were acquired during the same period t1. Then, the electron energy was increased to E2, which was slightly below the ionization energy of . The energy E2 continued to be increased in each iteration until the ion signal for was clearly detectable. During the second step (t2), the electron beam of energy E2 interacts with the mix of CCl4 and the products that resulted from the interaction in the first step, i.e., Cl− and . The mass spectrometer was set to detect the ion yield of during the second step t2. Because there was a mix of CCl4, Cl−, and during the second step inside the electron–molecule interaction region, the acquired could be produced by both the electron impact fragmentation of CCl4 molecules (from the target beam) and the electron impact ionization of the fragment formed during time t1. However, there was a difference between the appearance energy of from CCl4 and the ionization energy of the (c). In other words, the appearance energy of was over 3 eV higher than the ionization energy of . During the experiment, the electron energies in the steps, E1 and E2, were controllable. The same procedure can be applied to detect or exclude other possible neutrals, e.g., CCl, CCl2, and so on, produced in the DEA process by properly adjusting the E2 value. In addition, the period of t1 and t2 is controllable as well. For example, it was set such that t1 and t2 were equal to 1 s and 0.1 s, respectively, in the experiment published for the detection from DEA to CCl4 [32].

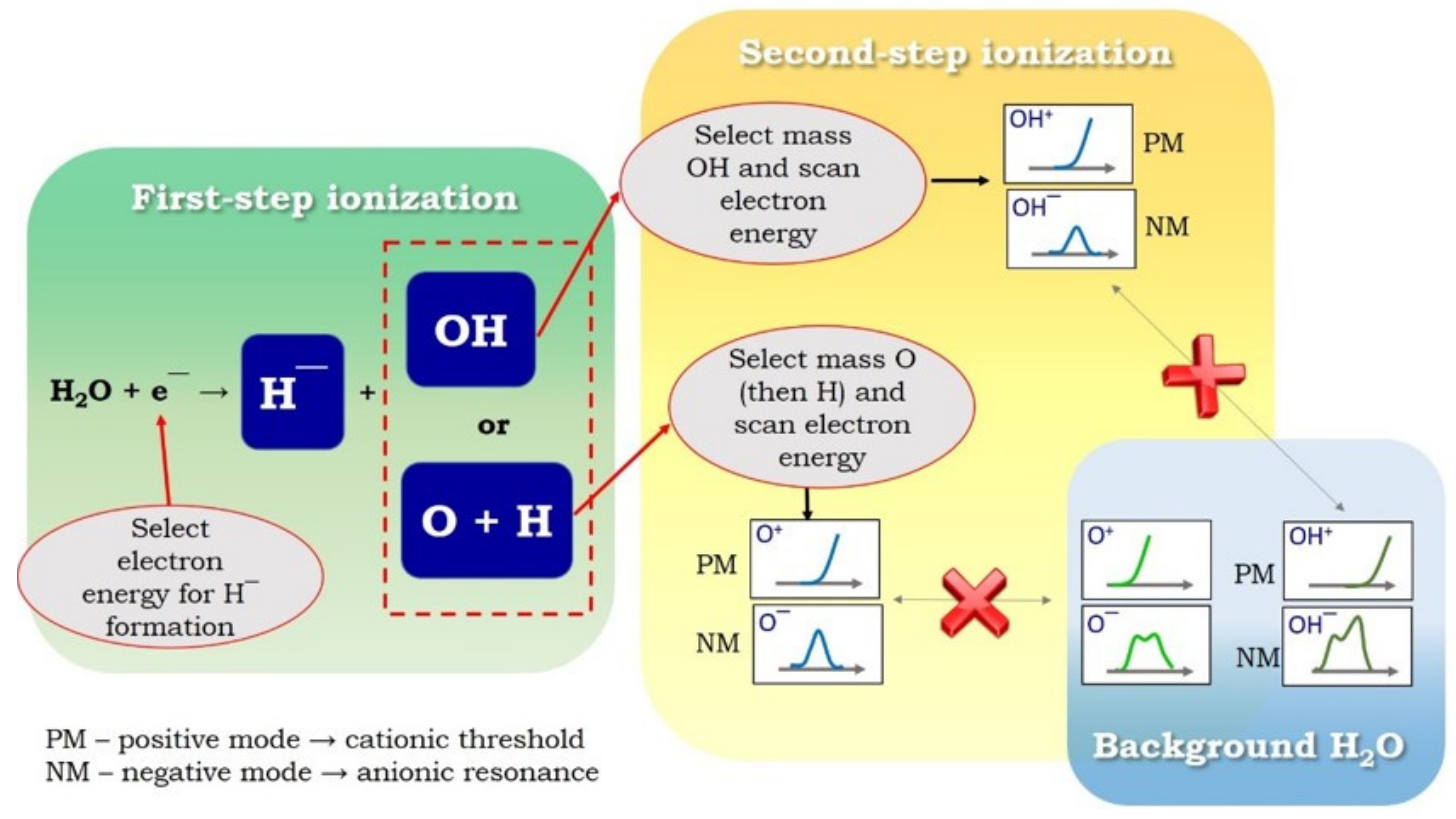
Figure 4.
Experimental procedure for neutral fragment identification from water using the two-step ionization method (see the description in the text). The cross mark indicates that the ionization threshold for a neutral fragment differs from the appearance energy of the same fragment formed directly from water and that the resonance structure for anion formation will show different spectra.
Figure 4.
Experimental procedure for neutral fragment identification from water using the two-step ionization method (see the description in the text). The cross mark indicates that the ionization threshold for a neutral fragment differs from the appearance energy of the same fragment formed directly from water and that the resonance structure for anion formation will show different spectra.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ptasinska, S. A Missing Puzzle in Dissociative Electron Attachment to Biomolecules: The Detection of Radicals. Atoms 2021, 9, 77. https://0-doi-org.brum.beds.ac.uk/10.3390/atoms9040077
AMA Style
Ptasinska S. A Missing Puzzle in Dissociative Electron Attachment to Biomolecules: The Detection of Radicals. Atoms. 2021; 9(4):77. https://0-doi-org.brum.beds.ac.uk/10.3390/atoms9040077
Chicago/Turabian StylePtasinska, Sylwia. 2021. "A Missing Puzzle in Dissociative Electron Attachment to Biomolecules: The Detection of Radicals" Atoms 9, no. 4: 77. https://0-doi-org.brum.beds.ac.uk/10.3390/atoms9040077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.