Potential Application of Tetrapleura tetraptera and Hibiscus sabdariffa (Malvaceae) in Designing Highly Flavoured and Bioactive Pito with Functional Properties


Abstract
:1. Introduction
2. Proximate Composition of Pito
3. Tetrapleura Tetraptera
3.1. Nutritional Profile
3.2. Phytochemical Composition
3.3. Biological Activities
3.3.1. Cytotoxic and Anti-Proliferative Activities
3.3.2. Hepatoprotective Activities
3.3.3. Anti-Inflammatory Effects
3.3.4. Antidiabetic Activity
3.3.5. Antioxidant Activity (AOA)
3.3.6. Biocidal Activities
3.4. Toxicological Aspect
4. Hibiscus Sabdariffa (Malvaceae)
4.1. Nutritional Profile
4.2. Phytochemical Composition
4.3. Biological Activities
4.3.1. Anti-Inflammatory Effects
4.3.2. Antioxidant Activities (AOA)
4.3.3. Antidiabetic and Anti-Hypertensive Effects
4.3.4. Biocidal Activities
4.4. Toxicological Aspect
5. Brewing Processes with Tetrapleura tetraptera and Hibiscus sabdariffa (Malvaceae)
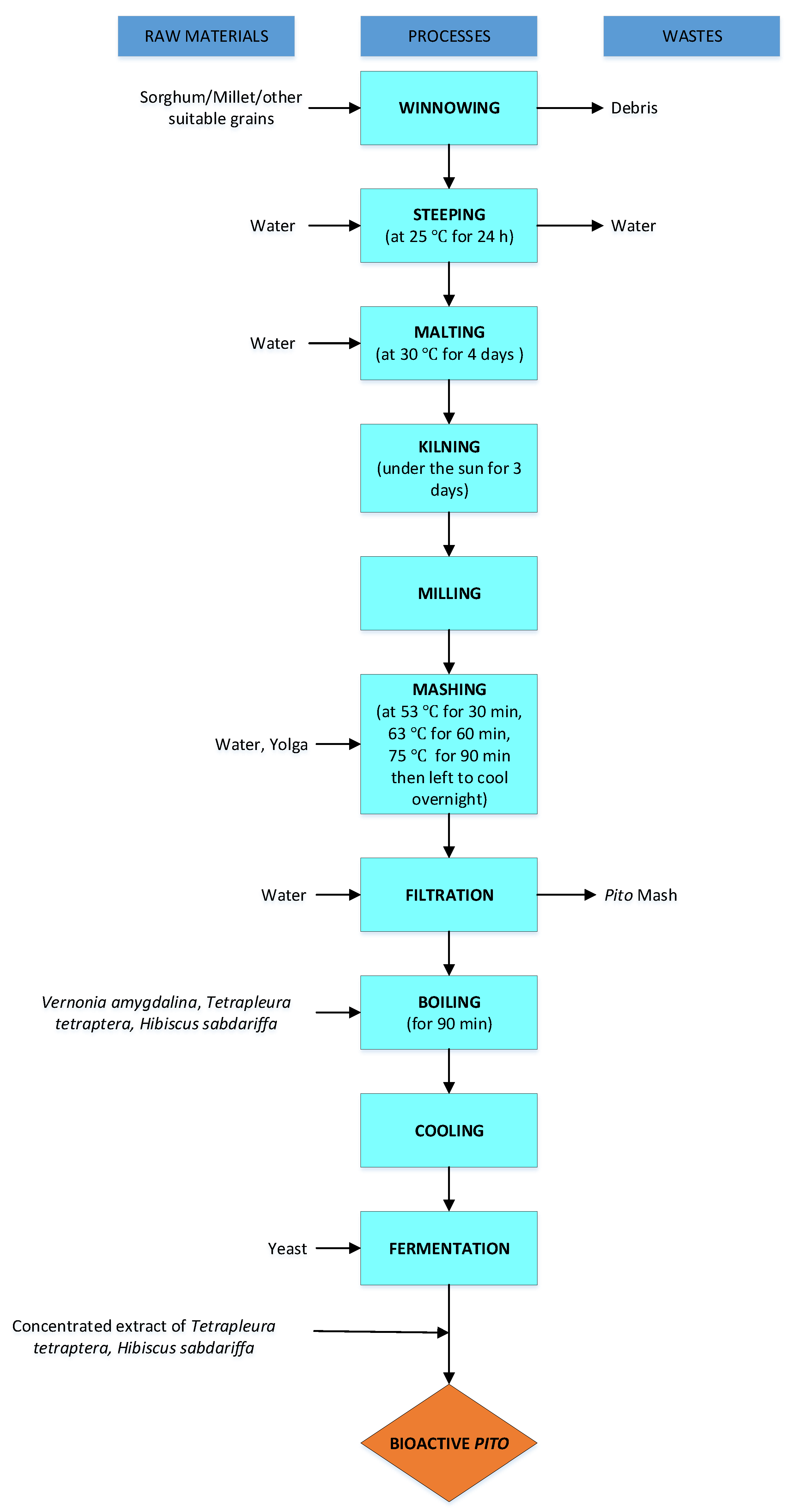
5.1. Indigenous Malting, Wort Production, and Fermentation
5.2. The Fate of Bioactive Compounds during Brewing
5.3. Encapsulation of Bioactive Compounds for Brewing Bioactive Pito (Scientific Outlook)
5.4. Pito Characterization
5.4.1. Colour
5.4.2. Alcohol Content (ABV)
5.4.3. pH
5.4.4. Volatile Compounds
5.4.5. Antioxidant Activity
6. Shelf Life of Bioactive Pito
7. Incidence of Heavy Metals in Pito Samples
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adadi, P.; Kovaleva, E.G.; Glukhareva, T.V.; Shatunova, S.A. Biotechnological production of non-traditional beer. AIP Conf. Proc. 2017, 1886, 1–13. [Google Scholar]
- Demuyakor, B.; Ohta, Y. Characteristics of pito yeasts from Ghana. Food Microbiol. 1991, 8, 183–193. [Google Scholar] [CrossRef]
- van der Aa Kühle, A.; Jesperen, L.; Glover, R.L.; Diawara, B.; Jakobsen, M. Identification and characterization of Saccharomyces cerevisiae strains isolated from West African sorghum beer. Yeast 2001, 18, 1069–1079. [Google Scholar] [CrossRef]
- Kolawole, O.; Kayode, R.; Akinduyo, B. Proximate and microbial analyses of burukutu and pito produced in Ilorin, Nigeria. Afr. J. Biotechnol. 2007, 6, 587–590. [Google Scholar]
- Lyumugabe, F.; Kamaliza, G.; Bajyana, E.; Thonart, P. Microbiological and physico-chemical characteristic of Rwandese traditional beer “Ikigage”. Afr. J. Biotechnol. 2010, 9, 4241–4246. [Google Scholar]
- Bansah, D. Traditional Brewing of Pito: Process and Product Characteristics. Master’s Thesis, University of Ghana, Legon, Ghana, 1990. [Google Scholar]
- Sefa-Dedeh, S.; Sanni, A.; Tetteh, G.; Sakyi-Dawson, E. Yeasts in the traditional brewing of pito in Ghana. World J. Microbiol. Biotechnol. 1999, 15, 593–597. [Google Scholar] [CrossRef]
- Adesina, S.K.; Iwalewa, E.O.; Johnny, I.I. Tetrapleura tetraptera Taub-ethnopharmacology, chemistry, medicinal and nutritional values-a review. J. Pharm. Res. Int. 2016, 1–22. [Google Scholar] [CrossRef]
- Adadi, P.; Barakova, N.V.; Muravyov, K.Y.; Krivoshapkina, E.F. Designing selenium functional foods and beverages: A review. Food Res. Int. 2019, 120, 708–725. [Google Scholar] [CrossRef]
- Ojewole, J.A.; Adewunmi, C.O. Anti-inflammatory and hypoglycaemic effects of Tetrapleura tetraptera (Taub)[fabaceae] fruit aqueous extract in rats. J. Ethnopharmacol. 2004, 95, 177–182. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Ebuehi, O.A.; Adeboyejo, F.O.; Aliyu, M.; Elemo, G.N. Modulatory effect of fibre-enriched cake on alloxan-induced diabetic toxicity in rat brain tissues. Toxicol. Rep. 2014, 1, 445–449. [Google Scholar] [CrossRef] [Green Version]
- Erukainure, O.L.; Onifade, O.F.; Odjobo, B.O.; Olasehinde, T.A.; Adesioye, T.A.; Tugbobo-Amisu, A.O.; Adenekan, S.O.; Okonrokwo, G.I. Ethanol extract of Tetrapleura tetraptera fruit peels: Chemical characterization, and antioxidant potentials against free radicals and lipid peroxidation in hepatic tissues. J. Taibah Univ. Sci. 2017, 11, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Igwe, O.; Akabuike, H. Free radical scavenging activity, phytochemistry and antimicrobial properties of Tetrapleura tetraptera Seeds. Int. Res. J. Chem. Chem. Sci. 2016, 3, 37–42. [Google Scholar]
- Akintola, O.; Bodede, A.; Ogunbanjo, O. Nutritional and medicinal importance of Tetrapleura tetraptera fruits (Aridan). Afr. J. Sci. Res. 2015, 4, 33–38. [Google Scholar]
- Ngassoum, M.; Jirovetz, L.; Buchbauer, G. SPME/GC/MS analysis of headspace aroma compounds of the Cameroonian fruit Tetrapleura tetraptera (Thonn.) Taub. Eur. Food Res. Technol. 2001, 213, 18–21. [Google Scholar] [CrossRef]
- Udourioh, G.A.; Etokudoh, M.F. Essential oils and fatty acids composition of dry fruits of Tetrapleura tetraptera. J. Appl. Sci. Environ. Manag. 2014, 18, 419–424. [Google Scholar]
- NATUPrekesedrink. Available online: https://www.facebook.com/natuprekesedrink/ (accessed on 11 February 2020).
- Plotto, A.; Mazaud, F.; Röttger, A.; Steffel, K. Hibiscus: Post-Production Management for Improved Market Access; Food and Agriculture Organization of the UN (FAO): Rome, Italy, 2004. [Google Scholar]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.–A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [Green Version]
- Morton, J.F.; Dowling, C.F. Fruits of Warm Climates; Creative Resources Systems: Miami, FL, USA, 1987; Volume 20534. [Google Scholar]
- Ross, I.A. Hibiscus sabdariffa. In Medicinal Plants of the World; Springer: Berlin/Heidelberg, Germany, 2003; pp. 267–275. [Google Scholar]
- Bako, I.; Mabrouk, M.; Abubakar, A. Antioxidant effect of ethanolic seed extract of Hibiscus sabdariffa linn (Malvaceae) alleviate the toxicity induced by chronic administration of sodium nitrate on some haematological parameters in wistars rats. Adv. J. Food Sci. Technol. 2009, 1, 39–42. [Google Scholar]
- Bolade, M.; Oluwalana, I.; Ojo, O. Commercial practice of roselle (Hibiscus sabdariffa L.) beverage production: Optimization of hot water extraction and sweetness level. World J. Agric. Sci. 2009, 5, 126–131. [Google Scholar]
- Ismail, A.; Ikram, E.H.K.; Nazri, H.S.M. Roselle (Hibiscus sabdariffa L.) seeds-nutritional composition, protein quality and health benefits. Food 2008, 2, 1–16. [Google Scholar]
- Okoro, C.E. Production of red wine from roselle (Hibiscus sabdariffa) and pawpaw (Carica papaya) using palm-wine yeast (Saccharomyces cerevisiae). Niger. Food J. 2007, 25, 158–164. [Google Scholar] [CrossRef]
- Rao, P.U. Nutrient composition and biological evaluation of mesta (Hibiscus sabdariffa) seeds. Plant Foods Hum. Nutr. 1996, 49, 27–34. [Google Scholar] [PubMed]
- Tsai, P.-J.; McIntosh, J.; Pearce, P.; Camden, B.; Jordan, B.R. Anthocyanin and antioxidant capacity in Roselle (Hibiscus sabdariffa L.) extract. Food Res. Int. 2002, 35, 351–356. [Google Scholar] [CrossRef]
- Nwahia, C.R.; Opara, C.C. Production of wine from zobo (Hibiscus sabdariffa) flower juice. J. Biochem. Technol. 2012, 3, 436–437. [Google Scholar]
- Alobo, A.P.; Offonry, S. Characteristics of coloured wine produced from roselle (Hibiscus sabdariffa) calyx extract. J. Inst. Brew. 2009, 115, 91–94. [Google Scholar] [CrossRef]
- Ali, S.A.E.; Mohamed, A.H.; Mohammed, G.E.E. Fatty acid composition, anti-inflammatory and analgesic activities of Hibiscus sabdariffa Linn. seeds. J. Adv. Vet. Anim. Res. 2014, 1, 50–57. [Google Scholar] [CrossRef]
- Eggensperger, H.; Wilker, M. Hibiscus-Extrakt: Ein hautverträglicher Wirkstoffkomplex aus AHA’s und polysacchariden. Teil 1. Parfümerie Und Kosmet. 1996, 77, 522–523. [Google Scholar]
- Amaya-Cruz, D.; Peréz-Ramírez, I.F.; Pérez-Jiménez, J.; Nava, G.M.; Reynoso-Camacho, R. Comparison of the bioactive potential of Roselle (Hibiscus sabdariffa L.) calyx and its by-product: Phenolic characterization by UPLC-QTOF MSE and their anti-obesity effect in vivo. Food Res. Int. 2019, 126, 108589. [Google Scholar] [CrossRef]
- Ali, B.H.; Wabel, N.A.; Blunden, G. Phytochemical, pharmacological and toxicological aspects of Hibiscus sabdariffa L.: A review. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2005, 19, 369–375. [Google Scholar]
- Adadi, P.; Barakova, N.V.; Krivoshapkina, E.F. Scientific approaches to improving artisan methods of producing local food condiments in Ghana. Food Control 2019. [Google Scholar] [CrossRef]
- Adadi, P.; Kovaleva, E.; Glukhareva, T.; Shatunova, S.; Petrov, A. Production and analysis of non-traditional beer supplemented with sea buckthorn. Agron. Res. 2017, 15, 1831–1845. [Google Scholar]
- Ducruet, J.; Rébénaque, P.; Diserens, S.; Kosińska-Cagnazzo, A.; Héritier, I.; Andlauer, W. Amber ale beer enriched with goji berries–The effect on bioactive compound content and sensorial properties. Food Chem. 2017, 226, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Kawa-Rygielska, J.; Adamenko, K.; Kucharska, A.Z.; Prorok, P.; Piórecki, N. Physicochemical and antioxidative properties of Cornelian cherry beer. Food Chem. 2019, 281, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Adazabra, A.N.; Ntiforo, A.; Bamford, S.A. Analysis of essential elements in P ito—A cereal food drink and its brands by the single-comparator method of neutron activation analysis. Food Sci. Nutr. 2014, 2, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Egemba, K.; Etuk, V.E. A kinetic study of burukutu fermentation. J. Eng. Appl. Sci. 2007, 2, 1193–1198. [Google Scholar]
- Ezeonu, C.S.; Nwokwu, C.D.U.; Kadiri, B. Comparative physico-chemical analysis of locally brewed beer (Burukutu) from corn, millet and sorghum. Am. J. Sci. Technol. 2017, 3, 43–48. [Google Scholar]
- Kumar, A.; Nair, A.; Reddy, A.; Garg, A. Analysis of essential elements in Pragya-peya—A herbal drink and its constituents by neutron activation. J. Pharm. Biomed. Anal. 2005, 37, 631–638. [Google Scholar] [CrossRef]
- Duodu, G.O.; Amartey, E.O.; Asumadu-Sakyi, A.B.; Adjei, C.A.; Quashie, F.K.; Nsiah-Akoto, I.; Ayanu, G. Mineral profile of pito from Accra, Tamale, Bolgatanga and Wa in Ghana. Food Public Health 2012, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Konfo, C.T.R.; Chabi, N.W.; Dahouenon-Ahoussi, E.; Cakpo-Chichi, M.; Soumanou, M.M.; Sohounhloue, D.C.K. Improvement of African traditional sorghum beers quality and potential applications of plants extracts for their stabilization: A review. J. Microbiol. Biotechnol. Food Sci. 2015, 5, 190. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, T.O.; Iliasu, G.A.; Adeleye, A.O.; Abdussalam, F.A.; Akinpelu, S.A.; Ogunbode, S.M.; Jimoh, S.O.; Oloyede, O.B. Nutritional and antioxidant dispositions of sorghum/millet-based beverages indigenous to Nigeria. Food Sci. Nutr. 2014, 2, 597–604. [Google Scholar] [CrossRef]
- Chevassus-Agnes, S.; Favier, J.; Joseph, A. Traditional technology and nutritive value of Cameroon sorghum beers. Cah. Onarest 1979, 2, 83–112. [Google Scholar]
- Nout, M. Composition of foods-African traditional beers. Food Lab. Newsl. 1987, 8, 18–20. [Google Scholar]
- Akin-Idowu, P.E.; Ibitoye, D.O.; Ademoyegun, O.T.; Adeniyi, O.T. Chemical composition of the dry fruit of Tetrapleura tetraptera and its potential impact on human health. J. Herbsspices Med. Plants 2011, 17, 52–61. [Google Scholar] [CrossRef]
- Abugri, D.A.; Pritchett, G. Determination of chlorophylls, carotenoids, and fatty acid profiles of Tetrapleura tetraptera seeds and their health implication. J. Herbsspices Med. Plants 2013, 19, 391–400. [Google Scholar] [CrossRef]
- Oguoma, O.; Ezeifeka, G.; Adeleye, S.; Oranusi, S.; Amadi, E. Antimicrobial activity, proximate and amino acids analysis of Tetrapleura Tetraptera. Niger. J. Microbiol. 2015, 27, 2709–2718. [Google Scholar]
- Leitzmann, C. Characteristics and health benefits of phytochemicals. Complementary Med. Res. 2016, 23, 69–74. [Google Scholar] [CrossRef]
- Godfrey, N.E. Proximate and phytochemical composition of the pulp of Tetrapleura tetraptera fruits consumed in Abakaliki, Nigeria. Int. J. Eng. Res. Technol. (Ijert) 2015, 4, 1286–1294. [Google Scholar]
- Irondi, E.A.; Oboh, G.; Agboola, S.O.; Boligon, A.A.; Athayde, M.L. Phenolics extract of Tetrapleura tetraptera fruit inhibits xanthine oxidase and Fe2+-induced lipid peroxidation in the kidney, liver, and lungs tissues of rats in vitro. Food Sci. Hum. Wellness 2016, 5, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Irondi, A.; Anokam, K.; Chukwuma, P. Phenological variation in the in-vitro antioxidant properties and alpha-amylase inhibitory activity of Tetrapleura tetraptera Pod. Int. J. Pharm. Sci. Drug Res. 2013, 5, 108–112. [Google Scholar]
- Chung, K.-T.; Wong, T.Y.; Wei, C.-I.; Huang, Y.-W.; Lin, Y. Tannins and human health: A review. Crit. Rev. Food Sci. Nutr. 1998, 38, 421–464. [Google Scholar] [CrossRef]
- Ashok, P.K.; Upadhyaya, K. Tannins are astringent. J. Pharmacogn. Phytochem. 2012, 1, 45–50. [Google Scholar]
- Igwe, O.U.; Okwu, D.E. Phytochemical composition and anti-inflammatory activities of Brachystegia eurycoma seeds and stem bark. Der Pharma Chem. 2013, 5, 224–228. [Google Scholar]
- Enujiugha, V.; Agbede, J. Nutritional and anti-nutritional characteristics of African oil bean (Pentaclethra macrophylla Benth) seeds. Appl. Trop. Agric. 2000, 5, 11–14. [Google Scholar]
- Mahmoud, M.H.; Abou-Arab, A.A.; Abu-Salem, F.M.; Aymerich, T.; Picouet, P.; Monfort, J.; Decker, E.; Chan, W.; Livisay, S.; Butterfield, D. Biochemistry, Practical/Research Method: A Fundamental Approach. Am. J. Food Technol. 2008, 12, 114–129. [Google Scholar]
- Rakić, S.; Maletić, R.O.; Perunović, M.N.; Svrzić, G. Influence of thermal treatment on tannin content and antioxidation effect of oak acorn Quercus cerris extract. J. Agric. Sci. Belgrade 2004, 49, 97–107. [Google Scholar] [CrossRef]
- Rausch, W.-D.; Liu, S.; Gille, G.; Radad, K. Neuroprotective effects of ginsenosides. Acta Neurobiol Exp (Wars) 2006, 66, 369–375. [Google Scholar] [PubMed]
- Sodipo, O.; Akinniyi, J.A.; Ogunbameru, J. Studies on certain characteristics of extracts of bark of Pausinystalia johimbe and Pausinystalia macroceras (K Schum) Pierre ex Beille. Glob. J. Pure Appl. Sci. 2000, 6, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Okolie, N.P.; Falodun, A.; Davids, O. Evaluation of the antioxidant activity of root extract of pepper fruit (Dennetia tripetala), and it’s potential for the inhibition of lipid peroxidation. Afr. J. Tradit. Complementary Altern. Med. 2014, 11, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, K.; Ghasemi, Y.; Ebrahimzadeh, M.A. Antioxidant activity, phenol and flavonoid contents of 13 citrus species peels and tissues. Pak. J. Pharm. Sci. 2009, 22, 277–281. [Google Scholar]
- Özçelik, B.; Kartal, M.; Orhan, I. Cytotoxicity, antiviral and antimicrobial activities of alkaloids, flavonoids, and phenolic acids. Pharm. Biol. 2011, 49, 396–402. [Google Scholar] [CrossRef]
- Amadi, S.W.; Zhang, Y.; Wu, G. Research progress in phytochemistry and biology of Aframomum species. Pharm. Biol. 2016, 54, 2761–2770. [Google Scholar] [CrossRef] [Green Version]
- Fecchio, D.; Sirois, P.; Russo, M.; Jancar, S. Studies on inflammatory response induced by Ehrlich tumor in mice peritoneal cavity. Inflammation 1990, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Ozaslan, M.; Karagoz, I.; Lawal, R.; Kilic, I.; Cakir, A.; Odesanmi, O.; Guler, I.; Ebuehi, O. Cytotoxic and anti-proliferative activities of the Tetrapleura tetraptera fruit extract on ehrlich ascites tumor cells. Int. J. Pharmacol. 2016, 12, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Atawodi, S.E.-O.; Yakubu, O.E.; Liman, M.L.; Iliemene, D.U. Effect of methanolic extract of Tetrapleura tetraptera (Schum and Thonn) Taub leaves on hyperglycemia and indices of diabetic complications in alloxan–induced diabetic rats. Asian Pac. J. Trop. Biomed. 2014, 4, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Onda, E.; Sonibare, M.; Ajayi, A.; Umukoro, S. Anti–inflammatory and Antioxidant Effects of Tetrapleura tetraptera (Schumach & Thonn.) Taub. Fruit Extract in Carrageenan/Kaolin-induced Acute Monoarthritis in Rats. Niger. J. Pharm. Res. 2018, 13, 157–166. [Google Scholar]
- Komlaga, G. Isolation and characterisation of the chemical constituents and the hypoglycaemic potentials of the fruit of Tetrapleura Tetraptera T.(Mimosaceae). Masters’s Thesis, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana, 2004. [Google Scholar]
- Kuate, D.; Kengne, A.P.N.; Biapa, C.P.N.; Azantsa, B.G.K.; Muda, W.A.M.B.W. Tetrapleura tetraptera spice attenuates high-carbohydrate, high-fat diet-induced obese and type 2 diabetic rats with metabolic syndrome features. Lipids Health Dis. 2015, 14, 50. [Google Scholar] [CrossRef] [Green Version]
- Famobuwa, O.; Lajide, L.; Owolabi, B.; Osho, I.; Amuho, U. Antioxidant activity of the fruit and stem bark of Tetrapleura tetraptera Taub (Mimosaceae). J. Pharm. Res. Int. 2016, 1–4. [Google Scholar] [CrossRef]
- Joel, J.; Sheena, O.; Martins, O.; Onyemauche, N.; Emmanuel, A. Comparative antioxidant capacity of aqueous and ethanol fruit extracts of Tetrapleura tetraptera. J. Biol. Sci. 2017, 17, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Moukette, B.M.; Pieme, A.C.; Biapa, P.C.N.; Njimou, J.R.; Stoller, M.; Bravi, M.; Yonkeu Ngogang, J. In vitro ion chelating, antioxidative mechanism of extracts from fruits and barks of Tetrapleura tetraptera and their protective effects against fenton mediated toxicity of metal ions on liver homogenates. Evid. -Based Complementary Altern. Med. 2015, 14. [Google Scholar] [CrossRef] [Green Version]
- Badu, M.; Mensah, J.K.; Boadi, N.O. Antioxidant activity of methanol and ethanol/water extracts of Tetrapleura tetraptera and Parkia biglobosa. Int. J. Pharm. Biol. Sci. 2012, 3, 312–321. [Google Scholar]
- Nwaichi, E.; Anyanwu, P. Effect of heat treatment on the antioxidant properties o f Tetrapleura tetraptera, Xylopia aethiopica and Piper guineense. Int. J. Biotechnol. Food Sci. 2013, 1, 1–5. [Google Scholar]
- Sonibare, M.A.; EE, E.E.O.; Ajayi, A.M.; Umukoro, S. In vitro antioxidant and membrane stabilization activities of fruit extract and fractions of Tetrapleura tetraptera (Schumach & Thonn.) Taub. J. Pharm. Bioresour. 2015, 12, 95–105. [Google Scholar]
- Nwidu, L.L.; Alikwe, P.C.N.; Elmorsy, E.; Carter, W.G. An investigation of potential sources of nutraceuticals from the Niger Delta areas, Nigeria for attenuating oxidative stress. Medicines 2019, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gberikon, G.; Adeoti, I.; Aondoackaa, A. Effect of ethanol and aqueous solutions as extraction solvents on phytochemical screening and antibacterial activity of fruit and stem bark extracts of Tetrapleura tetrapteraon Streptococcus salivarus and Streptococcus mutans. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 404–410. [Google Scholar]
- Achi, O. Composition and antibacterial activities of Tetrapleura tetraptera Taub. pod extracts. Res. J. Microbiol. 2010, 5, 1138–1144. [Google Scholar]
- Noamesi, B.K.; Mensah, J.F.; Bogale, M.; Dagne, E.; Adotey, J. Antiulcerative properties and acute toxicity profile of some African medicinal plant extracts. J. Ethnopharmacol. 1994, 42, 13–18. [Google Scholar] [CrossRef]
- Aladesanmi, A.J. Tetrapleura tetraptera: Molluscicidal activity and chemical constituents. Afr. J. Tradit. Complementary Altern. Med. 2007, 4, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Odesanmi, S.; Lawal, R.; Ojokuku, S. Effects of ethanolic extract of Tetrapleura tetraptera on liver function profile and histopathology in male Dutch white rabbits. Int. J. Trop. Med. 2009, 4, 136–139. [Google Scholar]
- Effiong, G.; Udoh, I.; Essien, G.; Ajibola, D.; Archibong, K. Effect of aqueous extract of Tetrapleura tetraptera on excision wounds in albino rats. Int. Res. J. Plant Sci. 2014, 5, 57–60. [Google Scholar]
- Juhari, N.H.; Petersen, M.A. Physicochemical Properties and Oxidative Storage Stability of Milled Roselle (Hibiscus sabdariffa L.) Seeds. Molecules 2018, 23, 385. [Google Scholar] [CrossRef] [Green Version]
- Salah, E.M.; Hayat, Z.E. Proximate composition of Karkadeh (Hibiscus sabdariffa) seeds and some functional properties of seed protein isolate as influenced by pH and NaCl. Int. J. Food Sci. Nutr. 2009, 60, 183–194. [Google Scholar] [CrossRef]
- Ayodeji, P.; Onoja, U.; Olanrewaju, O.; Aniebiet, T. Effects of Sorghum bicolor, Carica papaya and Hibiscus sabdariffa leaves extracts on some haematological indices of cyclophosphamide-induced anaemic albino rats. Niger. J. Nutr. Sci. 2019, 40, 142–151. [Google Scholar]
- Adanlawo, I.; Ajibade, V. Nutritive value of the two varieties of roselle (Hibiscus sabdariffa) calyces soaked with wood ash. Pak. J. Nutr. 2006, 5, 555–557. [Google Scholar]
- Nnam, N.; Onyeke, N. Chemical composition of two varieties of sorrel (Hibiscus sabdariffa L.), calyces and the drinks made from them. Plant Foods Hum. Nutr. 2003, 58, 1–7. [Google Scholar] [CrossRef]
- Kubuga, C.K.; Hong, H.G.; Song, W.O. Hibiscus sabdariffa meal improves iron status of childbearing age women and prevents stunting in their toddlers in Northern Ghana. Nutrients 2019, 11, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.-T.; Phan-Thi, H.; Pham-Hoang, B.-N.; Ho, P.-T.; Tran, T.T.T.; Waché, Y. Encapsulation of Hibiscus sabdariffa L. anthocyanins as natural colours in yeast. Food Res. Int. 2018, 107, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Jabeur, I.; Pereira, E.; Barros, L.; Calhelha, R.C.; Soković, M.; Oliveira, M.B.P.; Ferreira, I.C. Hibiscus sabdariffa L. as a source of nutrients, bioactive compounds and colouring agents. Food Res. Int. 2017, 100, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Mckay, D. Can hibiscus tea lower blood pressure. Agro Food Ind. Hi-Tech 2009, 20, 40–42. [Google Scholar]
- Baxter, K.; Driver, S.; Williamson, E. Stockley’s Herbal Medicines Interactions; Pharmaceutical Press: London, UK, 2013. [Google Scholar]
- Salah, A.; Gathumbi, J.; Vierling, W. Inhibition of intestinal motility by methanol extracts of Hibiscus sabdariffa L.(Malvaceae) in rats. Phytother. Res. 2002, 16, 283–285. [Google Scholar] [CrossRef]
- Alarcón-Alonso, J.; Zamilpa, A.; Aguilar, F.A.; Herrera-Ruiz, M.; Tortoriello, J.; Jimenez-Ferrer, E. Pharmacological characterization of the diuretic effect of Hibiscus sabdariffa Linn (Malvaceae) extract. J. Ethnopharmacol. 2012, 139, 751–756. [Google Scholar] [CrossRef]
- Maganha, E.G.; da Costa Halmenschlager, R.; Rosa, R.M.; Henriques, J.A.P.; de Paula Ramos, A.L.L.; Saffi, J. Pharmacological evidences for the extracts and secondary metabolites from plants of the genus Hibiscus. Food Chem. 2010, 118, 1–10. [Google Scholar] [CrossRef]
- Fernández-Arroyo, S.; Rodríguez-Medina, I.C.; Beltrán-Debón, R.; Pasini, F.; Joven, J.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Quantification of the polyphenolic fraction and in vitro antioxidant and in vivo anti-hyperlipemic activities of Hibiscus sabdariffa aqueous extract. Food Res. Int. 2011, 44, 1490–1495. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Herranz-López, M.; Barrajón-Catalán, E.; Arráez-Román, D.; Gonzálezlvarez, I.; Bermejo, M.; Gutiérrez, A.F.; Micol, V.; Segura-Carretero, A. Permeability study of polyphenols derived from a phenolic-enriched Hibiscus sabdariffa extract by UHPLC-ESI-UHR-Qq-TOF-MS. Int. J. Mol. Sci. 2015, 16, 18396–18411. [Google Scholar] [CrossRef] [PubMed]
- Meraiyebu, A.; Olaniyan, O.; Eneze, C.; Anjorin, Y.; Dare, J. Anti-inflammatory activity of methanolic extract of Hibiscus sabdariffa on carrageenan induced inflammation in wistar rat. Int. J. Pharm. Sci. Invent. 2013, 2, 22–24. [Google Scholar]
- Obouayeba, A.P.; Meité, S.; Boyvin, L.; Yeo, D.; Kouakou, T.H.; N’Guessan, J.D. Cardioprotective and anti-inflammatory activities of a polyphenols enriched extract of Hibiscus sabdariffa petal extracts in wistar rats. J. Pharmacogn. Phytochem. 2015, 4, 57–63. [Google Scholar]
- Jabeur, I.; Pereira, E.; Caleja, C.; Calhelha, R.C.; Soković, M.; Catarino, L.; Barros, L.; Ferreira, I.C. Exploring the chemical and bioactive properties of Hibiscus sabdariffa L. calyces from Guinea-Bissau (West Africa). Food Funct. 2019, 10, 2234–2243. [Google Scholar] [CrossRef] [Green Version]
- Dafallah, A.A.; Al-Mustafa, Z. Investigation of the anti-inflammatory activity of Acacia nilotica and Hibiscus sabdariffa. Am. J. Chin. Med. 1996, 24, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.; Le, H.; Pham, D.; Tran, T. Evaluation of physical, nutritional and sensorial properties cookie supplied with Hibiscus sabdariffa L. seed powder (without shell). Int. Food Res. J. 2018, 25, 1281–1287. [Google Scholar]
- Tseng, T.-H.; Wang, C.-J.; Kao, E.-S. Hibiscus protocatechuic acid protects against oxidative damage induced by tert-butylhydroperoxide in rat primary hepatocytes. Chem. -Biol. Interact. 1996, 101, 137–148. [Google Scholar] [CrossRef]
- Aganbi, E.; Onyeukwu, B.; Avwioroko, J.; Tonukari, J. Effect of fermentation on sensory, nutritional and antioxidant properties of mixtures of aqueous extracts of Hibiscus sabdariffa (zobo) and Raphia hookeri (raffia) wine. Niger. J. Sci. Environ. 2017, 15, 66–74. [Google Scholar]
- Usoh, I.; Akpan, E.; Etim, E.; Farombi, E. Antioxidant actions of dried flower extracts of Hibiscus sabdariffa L. on sodium arsenite-induced oxidative stress in rats. Pak. J. Nutr. 2005, 4, 135–141. [Google Scholar]
- Farombi, E.O.; Fakoya, A. Free radical scavenging and antigenotoxic activities of natural phenolic compounds in dried flowers of Hibiscus sabdariffa L. Mol. Nutr. Food Res. 2005, 49, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.; Ige, O. Hypolipidemic and antioxidant effects of ethanolic extract from dried calyx of Hibiscus sabdariffa in alloxan-induced diabetic rats. Fundam. Clin. Pharmacol. 2007, 21, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Duh, P.-D.; Yen, G.-C. Antioxidative activity of three herbal water extracts. Food Chem. 1997, 60, 639–645. [Google Scholar] [CrossRef]
- Mohd-Esa, N.; Hern, F.S.; Ismail, A.; Yee, C.L. Antioxidant activity in different parts of roselle (Hibiscus sabdariffa L.) extracts and potential exploitation of the seeds. Food Chem. 2010, 122, 1055–1060. [Google Scholar] [CrossRef]
- Mossalam, H.H.; Abd-El Aty, O.A.; Morgan, E.N.; Youssaf, S.; Mackawy, A.M.H. Biochemical and ultra structure studies of the antioxidant effect of aqueous extract of hibiscus sabdariffa on the nephrotoxicity induced by organophosphorous pesticide (malathion) on the adult albino rats. J. Am. Sci. 2011, 7, 561–572. [Google Scholar]
- Olaleye, M.T.; Rocha, B.J. Acetaminophen-induced liver damage in mice: Effects of some medicinal plants on the oxidative defense system. Exp. Toxicol. Pathol. 2008, 59, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Rosemary, R.; Haro, G. Antidiabetic effect of roselle calyces extract (Hibiscus sabdariffa L.) in streptozotocin induced mice. Int. J. Pharmtech Res. 2014, 6, 1703–1711. [Google Scholar]
- Sachdewa, A.; Khemani, L. Effect of Hibiscus rosa sinensis Linn. ethanol flower extract on blood glucose and lipid profile in streptozotocin induced diabetes in rats. J. Ethnopharmacol. 2003, 89, 61–66. [Google Scholar] [CrossRef]
- Sachdewa, A.; Nigam, R.; Khemani, L. Hypoglycemic effect of Hibiscus rosa sinensis L. leaf extract in glucose and streptozotocin induced hyperglycemic rats. Indian J. Exp. Biol. 2001, 39, 284–286. [Google Scholar]
- Mozaffari-Khosravi, H.; Jalali-Khanabadi, B.; Afkhami-Ardekani, M.; Fatehi, F.; Noori-Shadkam, M. The effects of sour tea (Hibiscus sabdariffa) on hypertension in patients with type II diabetes. J. Hum. Hypertens. 2009, 23, 48–54. [Google Scholar] [CrossRef] [Green Version]
- McKay, D.L.; Chen, C.O.; Saltzman, E.; Blumberg, J.B. Hibiscus sabdariffa L. tea (tisane) lowers blood pressure in prehypertensive and mildly hypertensive adults. J. Nutr. 2010, 140, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Obiefuna, P.; Owolabi, O.; Adegunloye, B.; Obiefuna, I.; Sofola, O. The petal extract of Hibiscus sabdariffa produces relaxation of isolated rat aorta. Int. J. Pharmacogn. 1994, 32, 69–74. [Google Scholar] [CrossRef]
- Ojeda, D.; Jiménez-Ferrer, E.; Zamilpa, A.; Herrera-Arellano, A.; Tortoriello, J.; Alvarez, L. Inhibition of angiotensin convertin enzyme (ACE) activity by the anthocyanins delphinidin-and cyanidin-3-O-sambubiosides from Hibiscus sabdariffa. J. Ethnopharmacol. 2010, 127, 7–10. [Google Scholar] [CrossRef]
- Mojiminiyi, F.; Adegunloye, B.; Egbeniyi, Y.; Okolo, R. An investigation of the diuretic effect of an aqueous extract of the petals of Hibiscus sabdariffa. J. Med. Med. Sci. 2000, 2, 77–80. [Google Scholar]
- Inuwa, I.; Ali, B.H.; Al-Lawati, I.; Beegam, S.; Ziada, A.; Blunden, G. Long-term ingestion of Hibiscus sabdariffa calyx extract enhances myocardial capillarization in the spontaneously hypertensive rat. Exp. Biol. Med. 2012, 237, 563–569. [Google Scholar] [CrossRef]
- Adegunloye, B.; Omoniyi, J.; Owolabi, O.; Ajagbonna, O.; Sofola, O.; Coker, H. Mechanisms of the blood pressure lowering effect of the calyx extract of Hibiscus sabdariffa in rats. Afr. J. Med. Med Sci. 1996, 25, 235–238. [Google Scholar]
- Ajay, M.; Chai, H.; Mustafa, A.; Gilani, A.H.; Mustafa, M.R. Mechanisms of the anti-hypertensive effect of Hibiscus sabdariffa L. calyces. J. Ethnopharmacol. 2007, 109, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-N.; Chan, K.-C.; Lin, W.-T.; Su, S.-L.; Wang, C.-J.; Peng, C.-H. Hibiscus sabdariffa inhibits vascular smooth muscle cell proliferation and migration induced by high glucose—A mechanism involves connective tissue growth factor signals. J. Agric. Food Chem. 2009, 57, 3073–3079. [Google Scholar] [CrossRef]
- Puro, K.-U.; Aochen, C.; Ghatak, S.; Das, S.; Sanjukta, R.; Mahapatra, K.P.; Jha, A.K.; Shakuntala, I.; Sen, A. Studies on the therapeutic properties of Roselle (Hibiscus sabdariffa) calyx: A popular ingredient in the cuisine of North East India. Int. J. Food Sci. Nutr. 2017, 2, 01–06. [Google Scholar]
- Fullerton, M.; Khatiwada, J.; Johnson, J.U.; Davis, S.; Williams, L.L. Determination of antimicrobial activity of sorrel (Hibiscus sabdariffa) on Esherichia coli O157: H7 isolated from food, veterinary, and clinical samples. J. Med. Food 2011, 14, 950–956. [Google Scholar] [CrossRef] [Green Version]
- Fakeye, T.O.; Pal, A.; Bawankule, D.; Yadav, N.; Khanuja, S. Toxic effects of oral administration of extracts of dried calyx of Hibiscus sabdariffa L. (Malvaceae). Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2009, 23, 412–416. [Google Scholar]
- Abubakar, M.; Lawal, A.; Suleiman, B.; Abdullahi, K. Hepatorenal toxicity studies of sub-chronic administration of calyx aqueous extracts of Hibiscus sabdariffa in albino rats. Bayero J. Pure Appl. Sci. 2010, 3, 16–19. [Google Scholar] [CrossRef]
- Sireeratawong, S.; Itharat, A.; Khonsung, P.; Lertprasertsuke, N.; Jaijoy, K. Toxicity studies of the water extract from the calyces of Hibiscus sabdariffa L. in rats. Afr. J. Tradit. Complementary Altern. Med. 2013, 10, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Onyenekwe, P.; Ajani, E.; Ameh, D.; Gamaniel, K. Antihypertensive effect of roselle (Hibiscus sabdariffa) calyx infusion in spontaneously hypertensive rats and a comparison of its toxicity with that in Wistar rats. Cell Biochem. Funct. 1999, 17, 199–206. [Google Scholar] [CrossRef]
- Gaya, I.; Mohammad, O.; Suleiman, A.; Maje, M.; Adekunle, A. Toxicological and lactogenic studies on the seeds of Hibiscus sabdariffa Linn (Malvaceae) extract on serum prolactin levels of albino wistar rats. Internet J. Endocrinol. 2009, 5, 1–6. [Google Scholar]
- Akindahunsi, A.; Olaleye, M. Toxicological investigation of aqueous-methanolic extract of the calyces of Hibiscus sabdariffa L. J. Ethnopharmacol. 2003, 89, 161–164. [Google Scholar] [CrossRef]
- Adadi, P.; Kovaleva, E.; Glukhareva, T.; Barakova, N. Production and investigations of antioxidant rich beverage: Utilizing Monascus purpureus IHEM LY2014-0696 and various malts. Agron. Res 2018, 16, 1312–1321. [Google Scholar]
- Nsengumuremyi, D.; Adadi, P.; Ukolova, M.V.; Barakova, N.V. Effects of ultradisperse humic sapropel suspension on microbial growth and fermentation parameters of barley dstillate. Fermentation 2019, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Sore, A. ALBERT SORE’S MEMOIR-Pito: Nothern Ghana’s Traditional Brew. Available online: https://www.a1radioonline.com/6494/albert-sores-memoir-pito-nothern-ghanas-traditional-brew/index.html (accessed on 11 February 2020).
- Lyumugabe, F.; Uyisenga, J.P.; Songa, E.B.; Thonart, P. Production of traditional sorghum beer “Ikigage” using Saccharomyces cerevisae, Lactobacillus fermentum and Issatckenkia orientalis as starter cultures. Food Nutr. Sci. 2014, 6, 507–515. [Google Scholar]
- Mbatchou, V.C.; Nabayire, K.O.; Akuoko, Y. Vernonia amygdalina Leaf: Unveiling its antacid and carminative properties In Vitro. Curr. Sci. 2017, 3, 148–155. [Google Scholar]
- Erasto, P.; Grierson, D.S.; Afolayan, A.J. Evaluation of antioxidant activity and the fatty acid profile of the leaves of Vernonia amygdalina growing in South Africa. Food Chem. 2007, 104, 636–642. [Google Scholar] [CrossRef]
- Erasto, P.; Grierson, D.; Afolayan, A. Antioxidant constituents in Vernonia amygdalina. Leaves. Pharm. Biol. 2007, 45, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Habtamu, A.; Melaku, Y. Antibacterial and antioxidant compounds from the flower extracts of Vernonia amygdalina. Adv. Pharmacol. Sci. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I.; Kabbashi, N.A. Extraction and characterization of bioactive compounds in Vernonia amygdalina leaf ethanolic extract comparing Soxhlet and microwave-assisted extraction techniques. J. Taibah Univ. Sci. 2019, 13, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Fumi, M.D.; Galli, R.; Lambri, M.; Donadini, G.; De Faveri, D.M. Effect of full-scale brewing process on polyphenols in Italian all-malt and maize adjunct lager beers. J. Food Compos. Anal. 2011, 24, 568–573. [Google Scholar] [CrossRef]
- Zhao, H. Effects of processing stages on the profile of phenolic compounds in beer. In Processing and Impact on Active Components in Food; Elsevier: Amsterdam, The Netherlands, 2015; pp. 533–539. [Google Scholar]
- Pascoe, H.M.; Ames, J.M.; Chandra, S. Critical stages of the brewing process for changes in antioxidant activity and levels of phenolic compounds in ale. J. Am. Soc. Brew. Chem. 2003, 61, 203–209. [Google Scholar] [CrossRef]
- Koren, D.; Kun, S.; Vecseri, B.H.; Kun-Farkas, G. Study of antioxidant activity during the malting and brewing process. J. Food Sci. Technol. 2019, 56, 3801–3809. [Google Scholar] [CrossRef] [Green Version]
- Szwajgier, D. Content of individual phenolic acids in worts and beers and their possible contribution to the antiradical activity of beer. J. Inst. Brew. 2009, 115, 243–252. [Google Scholar] [CrossRef]
- Limón, R.I.; Peñas, E.; Torino, M.I.; Martínez-Villaluenga, C.; Dueñas, M.; Frias, J. Fermentation enhances the content of bioactive compounds in kidney bean extracts. Food Chem. 2015, 172, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Soto, S.A.; Beaufort, S.; Bouajila, J.; Souchard, J.-P.; Renard, T.; Rollan, S.; Taillandier, P. Impact of fermentation conditions on the production of bioactive compounds with anticancer, anti-inflammatory and antioxidant properties in kombucha tea extracts. Process Biochem. 2019, 83, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Verni, M.; Verardo, V.; Rizzello, C.G. How fermentation affects the antioxidant properties of cereals and legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Corona, M.; Sánchez-Martínez, M.; Valderrama, M.; Rodríguez, M.; Cámara, C.; Madrid, Y. Selenium biotransformation by Saccharomyces cerevisiae and Saccharomyces bayanus during white wine manufacture: Laboratory-scale experiments. Food Chem. 2011, 124, 1050–1055. [Google Scholar] [CrossRef]
- Sánchez-Martínez, M.; da Silva, E.G.P.; Pérez-Corona, T.; Cámara, C.; Ferreira, S.L.; Madrid, Y. Selenite biotransformation during brewing. Evaluation by HPLC–ICP-MS. Talanta 2012, 88, 272–276. [Google Scholar]
- Singthong, J.; Oonsivilai, R.; Oonmetta-Aree, J.; Ningsanond, S. Bioactive compounds and encapsulation of Yanang (Tiliacora triandra) leaves. Afr. J. Tradit. Complementary Altern. Med. 2014, 11, 76–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirós-Sauceda, A.E.; Ayala-Zavala, J.F.; Olivas, G.I.; González-Aguilar, G.A. Edible coatings as encapsulating matrices for bioactive compounds: A review. J. Food Sci. Technol. 2014, 51, 1674–1685. [Google Scholar] [CrossRef] [Green Version]
- Djameh, C.; Ellis, W.; Oduro, I.; Saalia, F.; Haslbeck, K.; Komlaga, G. West African sorghum beer fermented with Lactobacillus delbrueckii and Saccharomyces cerevisiae: Fermentation by-products. J. Inst. Brew. 2019, 125, 326–332. [Google Scholar] [CrossRef]
- Wang, M.-L.; Choong, Y.-M.; Su, N.-W.; Lee, M.-H. A rapid method for determination of ethanol in alcoholic beverages using capillary gas chromatography. J. Food Drug Anal. 2003, 11, 133–140. [Google Scholar]
- Demuyakor, B.; Ohta, Y. Characteristics of single and mixed culture fermentation of pito beer. J. Sci. Food Agric. 1993, 62, 401–408. [Google Scholar] [CrossRef]
- Yeboah-Awudzi, M. Microbial Hazard Analysis and Development of Control Measures for Bissap Drink (sobolo). Master’s Thesis, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana, 2017. [Google Scholar]
- Ayirezang, F.A.; Saba, C.K.S.; Amagloh, F.K.; Gonu, H. Shelf life improvement of sorghum beer (pito) through the addition of moringa oleifera and pasteurization. Afr. J. Biotechnol. 2016, 15, 2627–2636. [Google Scholar]
- Avicor, M.; Saalia, F.; Djameh, C.; Sinayobye, E.; Mensah-Brown, H.; Essilfie, G. The fermentation characteristics of single and mixed yeast cultures during pito wort fermentation. Int. Food Res. J. 2015, 22, 102–109. [Google Scholar]
- Lyumugabe, F.; Gros, J.; Songa, E.B.; Thonart, P. Sorghum beer brewing using eleusine coracana “Finger Millet” to improve the saccharification. Am. J. Food Technol. 2015, 10, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Bamforth, C.W. pH in brewing: An overview. Tech. Q. -Master Brew. Assoc. Am. 2001, 38, 1–9. [Google Scholar]
- Attchelouwa, C.K.; Aka-Gbézo, S.; N’guessan, F.K.; Kouakou, C.A.; Djè, M.K. Biochemical and microbiological changes during the Ivorian sorghum beer deterioration at different storage temperatures. Beverages 2017, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Djameh, C.; Ellis, W.; Oduro, I.; Saalia, F.; Blay, Y.; Komlaga, G. West African sorghum beer fermented with Lactobacillus delbrueckii and Saccharomyces cerevisiae: Shelf-life and consumer acceptance. J. Inst. Brew. 2019, 125, 333–341. [Google Scholar] [CrossRef]
- Onyenekwe, P.C.; Erhabor, G.O.; Akande, S.A. Characterisation of aroma volatiles of indigenous alcoholic beverages: Burukutu and pito. Nat. Prod. Res. 2016, 30, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Lyumugabe, F.; Gros, J.; Thonart, P.; Collin, S. Occurrence of polyfunctional thiols in sorghum beer ‘ikigage’made with Vernonia amygdalina ‘umubirizi’. Flavour Fragr. J. 2012, 27, 372–377. [Google Scholar] [CrossRef]
- Klug, L.; Daum, G. Yeast lipid metabolism at a glance. FEMS Yeast Res. 2014, 14, 369–388. [Google Scholar] [CrossRef] [Green Version]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Dufour, J.P.; Malcorps, P.; Silcock, P. Control of ester synthesis during brewery fermentation. Brew. Yeast Ferment. Perform. 2003, 213–233. [Google Scholar]
- Gülçin, İ.; Huyut, Z.; Elmastaş, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Shewakena, S.; Chandravanshi, B.; Debebe, A. Levels of total polyphenol, flavonoid, tannin and antioxidant activity of selected Ethiopian fermented traditional beverages. Int. Food Res. J. 2017, 24, 2033–2040. [Google Scholar]
- Marova, I.; Parilova, K.; Friedl, Z.; Obruca, S.; Duronova, K. Analysis of phenolic compounds in lager beers of different origin: A contribution to potential determination of the authenticity of Czech beer. Chromatographia 2011, 73, 83–95. [Google Scholar] [CrossRef]
- Kao, F.-J.; Chiu, Y.-S.; Chiang, W.-D. Effect of water cooking on antioxidant capacity of carotenoid-rich vegetables in Taiwan. J. Food Drug Anal. 2014, 22, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Demuyakor, B. Exploitation of Ghanaian raw materials in tropical beer brewing. Ph.D. Thesis, Hiroshima University, Hiroshima, Japan, 1994. [Google Scholar]
- Sanni, A.; Onilude, A.; Fadahunsi, I.; Afolabi, R. Microbial deterioration of traditional alcoholic beverages in Nigeria. Food Res. Int. 1999, 32, 163–167. [Google Scholar] [CrossRef]
- Council, N.R. Applications of Biotechnology in Traditional Fermented Foods; National Academies Press: Washington, DC, USA, 1992. [Google Scholar]
- Ellis, W.; Oduro, I.; Terkuu, D. Preliminary studies on extension of the shelflife of pito. J. Sci. Technol. (Ghana) 2005, 25, 11–15. [Google Scholar] [CrossRef]
- Evera, E.; Abedin Abdallah, S.H.; Shuang, Z.; Sainan, W.; Yu, H. Shelf life and nutritional quality of sorghum beer: Potentials of phytogenic-based extracts. J. Agric. Food. Tech. 2019, 2, 1–14. [Google Scholar]
- Cabrera, C.; Lloris, F.; Gimenez, R.; Olalla, M.; Lopez, M.C. Mineral content in legumes and nuts: Contribution to the Spanish dietary intake. Sci. Total Environ. 2003, 308, 1–14. [Google Scholar] [CrossRef]
- Anderson, R.A.; Bryden, N.A.; Polansky, M.M. Dietary chromium intake. Biol. Trace Elem. Res. 1992, 32, 117–121. [Google Scholar] [CrossRef]
- Ebong, A.; Chapel, A.; Matinus, N.; Alexander, D. Foliar Fertilization Stuttgar; Fischer-verlag: Frankfurt, Germany, 2006. [Google Scholar]
- Dan, E.U.; Ebong, G.A. Impact of cooking utensils on trace metal levels of processed food items. Ann. Food Sci. Technol 2013, 14, 350–355. [Google Scholar]
- Dabonne, S.; Koffi, B.; Kouadio, E.; Koffi, A.; Due, E.; Kouame, L. Traditional utensils: Potential sources of poisoning by heavy metals. Br. J. Pharmacol. Toxicol. 2010, 1, 90–92. [Google Scholar]
- Oz, S.; Teke, G.; Anli, R.E.; Atakol, O. Heavy Metal Content of Selected Some Turkish Beers. Int. J. Sci. Technol. Res. 2015, 1, 46–56. [Google Scholar]
- Debebe, A.; Chandravanshi, B.S.; Abshiro, M. Assessment of essential and non-essential metals in Ethiopian traditional fermented alcoholic beverages. Bull. Chem. Soc. Ethiop. 2017, 31, 17–30. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients | [40] | [44] c | [45] c | [46] c |
|---|---|---|---|---|
| Crude protein | 1.47 ± 0.01 a% 1.95 ± 0.01 b% 2.74 ± 0.01 c% | 3.70 ± 0.02% | 8.7 g | 3.9 g |
| Dry matter | × | × | × | 7.9 g |
| Crude fat | 0.52 ± 0.01 a% 0.80 ± 0.01 b% 1.07 ± 0.01 c% | 0.31 ± 0.01% | 0.3 g | × |
| Ash | 0.92 ± 0.01 a% 1.25 ± 0.01 b% 1.74 ± 0.01 c% | 1.50 ± 0.01% | 4.1 g | × |
| Available carbohydrate | × | 5.60 ± 0.31 g | × | 4.8 |
| Crude fibre | × | 0.20 ± 0.01% | × | |
| Moisture | 91.05 ± 0.00 a% 90.41 ± 0.01 b% 89.71 ± 0.01 c% | 88.69 ± 0.48 | × | × |
| Vitamin B1 | 0.0515 ± 0.0005 a IU/mL 0.0575 ± 0.0005 b IU/mL 0.0645 ± 0.0005 c IU/mL | × | × | 0.11 mg |
| Vitamin B2 | 0.0420 ± 0.0010 a mg/100 mL 0.0515 ± 0.0005 b mg/100 mL 0.0625 ± 0.0005 c mg/100 mL | × | × | 0.05 |
| Vitamin B6 | 0.035 ± 0.0005 a mg/100 mL 0.046 ± 0.0010 b mg/100 mL 0.052 ± 0.0010 c mg/100 mL | × | × | × |
| Vitamin C | × | × | × | 0.04 |
| Calcium | 24.625 ± 0.005 a mg 29.80 ± 0.010 b mg 31.57 ± 0.010 c mg | × | 20.7 mg | 2.2 mg |
| Iron | 1.865 ± 0.005 a mg 2.055 ± 0.015 b mg 2.460 ± 0.010 c mg | × | × | 2.55 mg |
| Zinc | 1.040 ± 0.010 a mg 1.680 ± 0.010 b mg 2.485 ± 0.015 c mg | × | × | × |
| Magnesium | 48.675 ± 0.015 a mg 61.965 ± 0.015 b mg 68.350 ± 0.010 c mg | × | × | × |
| Potassium | × | × | 1101 mg | 84 mg |
| Sodium | × | × | 26.9 mg | 1.1 |
| Niacin | × | × | 8 µg | 0.43 |
| Riboflavine | × | × | 760 µg | |
| Thiamine | × | × | 3441 µg | |
| Phosphorus | × | × | × | 39 |
| Pantothenic acid | × | × | × | 0.09 |
| Energy value (kJ/g) | × | 164.00 ± 3.12 | 394 | 164 |
| Fruit Part | Total Polyphenols | Flavonoids | Saponins | Tannin | Phytate |
|---|---|---|---|---|---|
| Seeds | 38.05 ± 0.21 C | 10.30 ± 0.42 C | 60.80 ± 11.88 C | 675.50 ± 152.03 B | 3545.00 ± 77.78 B |
| Pulp | 1866.88 ± 1.02 B | 410.75 ± 1.06 A | 953.40 ± 9.33 A | 1097.50 ± 26.16 A | 5170.00 ± 42.43 A |
| Woody shell | 2907.15 ± 2.19 A | 354.60 ± 0.85 B | 641.50 ± 18.81 B | 135.50 ± 20.51 C | 1021.00 ± 15.56 C |
| Mean | 1604.03 | 258.55 | 551.90 | 636.17 | 3245.33 |
| Phenolics | Concentration (mg/g) |
|---|---|
| Gallic acid 1 | 2.95 ± 0.01 |
| Catechin 1 | 0.43 ± 0.03 |
| Chlorogenic acid 1 | 0.21 ± 0.01 |
| Caffeic acid 1 | 3.72 ± 0.02 |
| Ellagic acid 1 | 3.69 ± 0.04 |
| Epicatechin 1 | 1.38 ± 0.01 |
| Rutin 1 | 1.74 ± 0.01 |
| Quercitin 1 | 3.65 ± 0.03 |
| Luteolin 1 | 0.45 ± 0.02 |
| Apigenin 1 | 3.73 ± 0.01 |
| Total phenol 2 | 27.48 ± 0.16 α 42.18 ± 0.16 β |
| Tannin 2 | 32.54 ± 0.22 α 46.99 ± 0.17 β |
| Total flavonoid 2 | 0.18 ± 0.01 α 0.44 ± 0.02 β |
| Phenolics | Concentration (ppm) |
|---|---|
| Hydroxycitric acid 1 | 8288.03 ± 397.63 |
| Hibiscicus acid 1 | 31122.02 ± 1128.39 |
| Chlorogenic acid isomer I 1 | 2755.15 ± 62.42 |
| Chlorogenic acid 1 | 1923.72 ± 38.69 |
| Chlorogenic acid isomer II 1 | 1041.19 ± 16.96 |
| Myricetin 3-arabinogalactose 1 | 57.32 ± 2.51 |
| Quercetin 3-sambubioside 1 | 304.02 ± 5.90 |
| 5-O-Caffeoylshikimic acid 1 | 171.47 ± 6.92 |
| Quercetin 3-rutinoside 1 | 495.70 ± 4.34 |
| Quercetin 3-glucoside 1 | 143.74 ± 2.16 |
| Kaempferol 3-O-rutinoside 1 | 91.86 ± 2.28 |
| N-Feruloyltyramine 1 | 98.97 ± 1.80 |
| Kaempferol 3-(p-coumarylglucoside) 1 | 28.37 ± 0.48 |
| Quercetin 1 | 121.24 ± 2.01 |
| 7-Hydroxycoumarin 1 | 1839.20 ± 25.34 |
| Delphinidin 3-sambubioside 1 | 2701.21 ± 165.55 |
| Cyanidin 3-sambubioside 1 | 1939.15 ± 39.27 |
| Hisbiscus acid hydroxyethylesther 2 | 235.05 |
| Hisbiscus acid dimethylesther 2 | 217.04 |
| 2-O-trans-cafeoyl-hydroxicitric acid 2 | 369.05 |
| Methylepigallocatechin 2 | 319.08 |
| Ethylchlorogenate 2 | 381.12 |
| 2-O-trans-feruloyl-hydroxicitric acid 2 | 263.08 |
| Hibiscus acid hydroxyethyldimethylesther 2 | 263.08 |
| Coumaroylquinic acid 2 | 337.09 |
| Cryptochlorogenic acid 2 | 353.09 |
| Methyl digallate 2 | 335.04 |
| Parameters | Extract | Must | HS wine |
|---|---|---|---|
| oBrix | 2.00 ± 0.02 a 2.50 ± 0.02 b | 1600 ± 0.02 a 22.00 ± 0.20 b | 4.90 ± 0.02 a 5.10 ± 0.01 b |
| pH | 3.78 ± 0.03 a 2.34 ± 0.14 b | 3.72 ± 0.04 a 3.76 ± 0.08 b | 3.43 ± 0.04 a 3.57 ± 0.02 b |
| Colour (visual) | Deep red a,b | Deep red a Dull red b | Brilliant rose a,b |
| Alcohol (% w/v) | 0.00 a,b | 0.00 a,b | 10.80 ± 0.08 a 10.50 ± 0.01 b |
| Ash (%) | 0.70 ± 0.05 a | 0.68 ± 0.02 a | 0.43 ± 0.04 a |
| Total anthocyanins (abs/mL) | 28.43 ± 0.04 a | 28.30 ± 0.10 a | 22.65 ± 0.08 a |
| Crude protein (%) | 2.63 ± 0.23 a | 4.21 ± 0.26 a | 1.75 ± 0.09 a |
| Ascorbic acid (mg/100 mL) | 1.12 ± 0.06 a | 1.06 ± 0.14 a | 0.6 ± 90.13 a |
| Sulphur dioxide (%) | 0.00 a | 13.40 ± 0.12 a | 10.50 ± 0.06 a |
| Ethanol Content | Author |
|---|---|
| 4.54 ± 0.04 £%ABV (SSC) 4.5 ± 0.03 †%ABV (MSC) | [1] |
| 3.93%ABV | [159] |
| 2.2 ± 0.46% (v/v) | [5] |
| 2.78 ± 0.02%ABV (SSC) 2.73 ± 0.01%ABV (MSC) | [160] |
| 2.30 ×% (v/v) 3.10 α% (v/v) | [161] |
| 4.2 ± 0.3 †%, v/v 3.6 ± 0.1 £ | [155] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adadi, P.; Kanwugu, O.N. Potential Application of Tetrapleura tetraptera and Hibiscus sabdariffa (Malvaceae) in Designing Highly Flavoured and Bioactive Pito with Functional Properties. Beverages 2020, 6, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6020022
Adadi P, Kanwugu ON. Potential Application of Tetrapleura tetraptera and Hibiscus sabdariffa (Malvaceae) in Designing Highly Flavoured and Bioactive Pito with Functional Properties. Beverages. 2020; 6(2):22. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6020022
Chicago/Turabian StyleAdadi, Parise, and Osman N. Kanwugu. 2020. "Potential Application of Tetrapleura tetraptera and Hibiscus sabdariffa (Malvaceae) in Designing Highly Flavoured and Bioactive Pito with Functional Properties" Beverages 6, no. 2: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/beverages6020022