
Molecular Mechanism of Malignant Transformation of Balb/c-3T3 Cells Induced by Long-Term Exposure to 1800 MHz Radiofrequency Electromagnetic Radiation (RF-EMR)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. sXc-1800 Exposure System
2.2. Cell Culture and Exposure Protocol
2.3. Cell Transformation Assay
2.4. Transwell Assay
2.5. Cell Transplantation in SCID Mice
2.6. Soft Agar Clone Formation
2.7. mRNA Microarray Analysis
2.8. Gene Ontology (GO), Reactome, and KEGG (Kyoto Encyclopedia of Genes and Genomes) Pathway Enrichment Analysis
2.9. Protein–Protein Interaction (PPI) Network Analysis
2.10. Statistical Analysis
3. Results
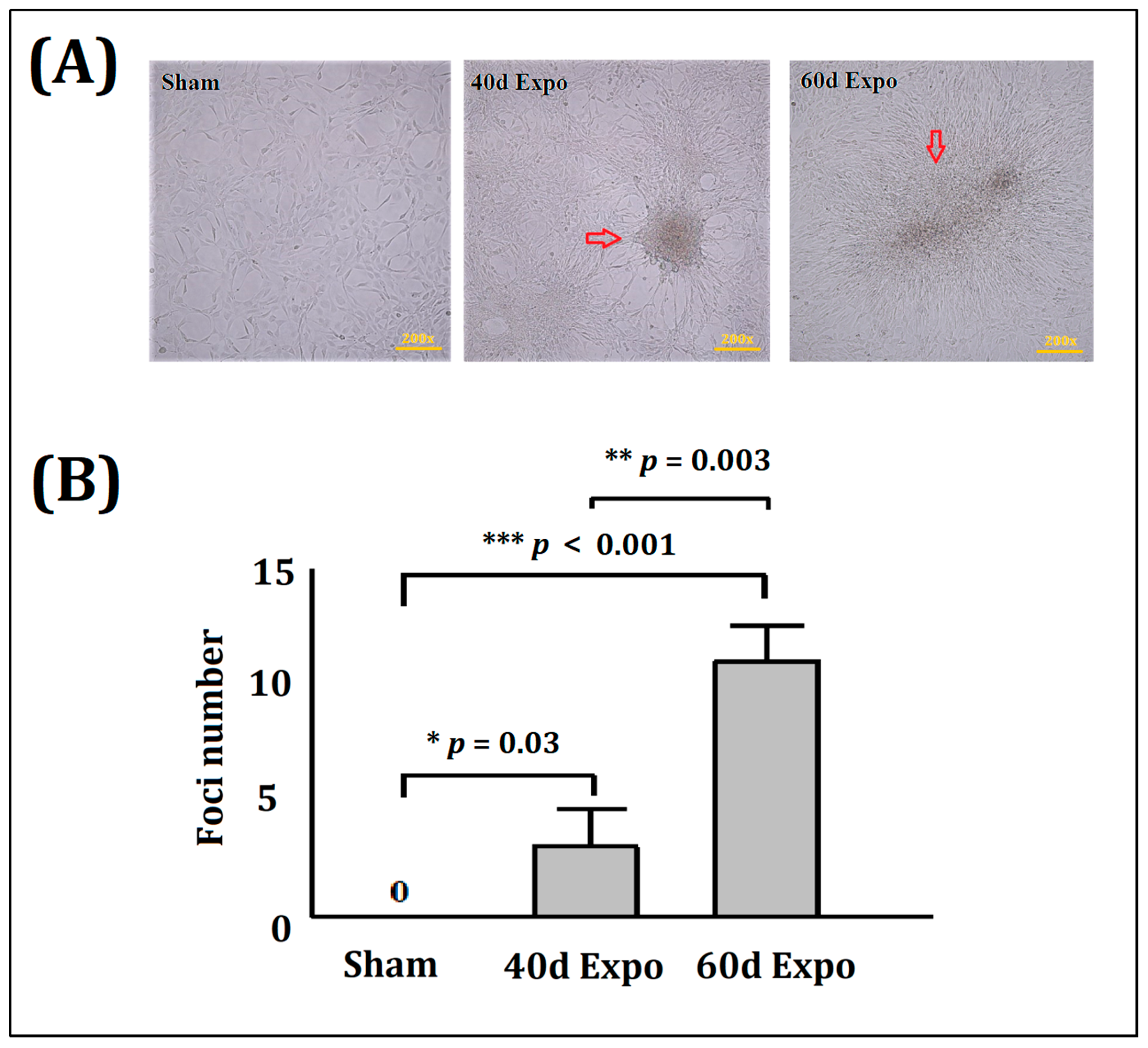
3.1. Malignant Transformation of Balb/c-3T3 Cells Induced by 1800 MHzRF-EMR
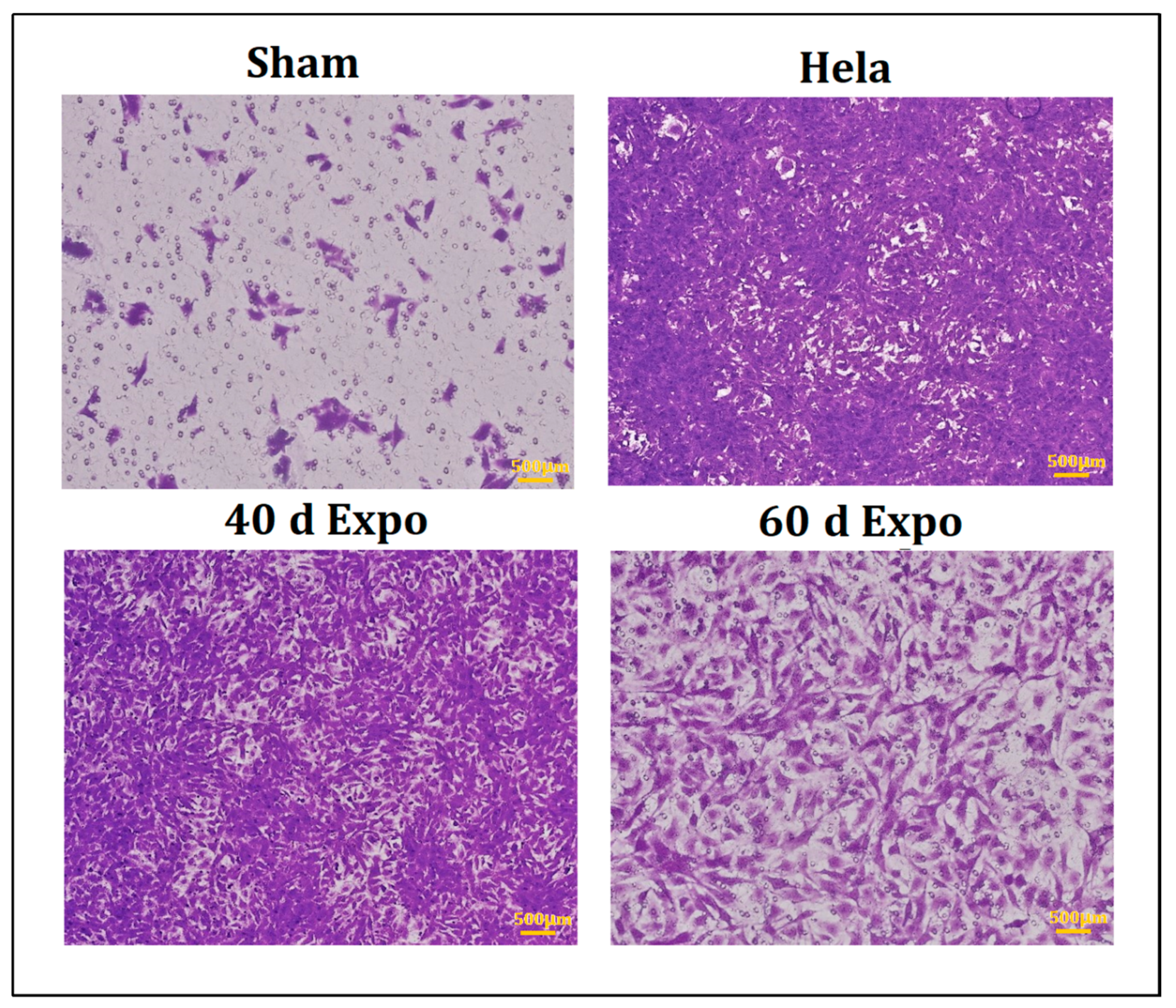
3.2. 1800 MHz EMR Enhances Balb/c-3T3 Cells’ Ability to Migrate
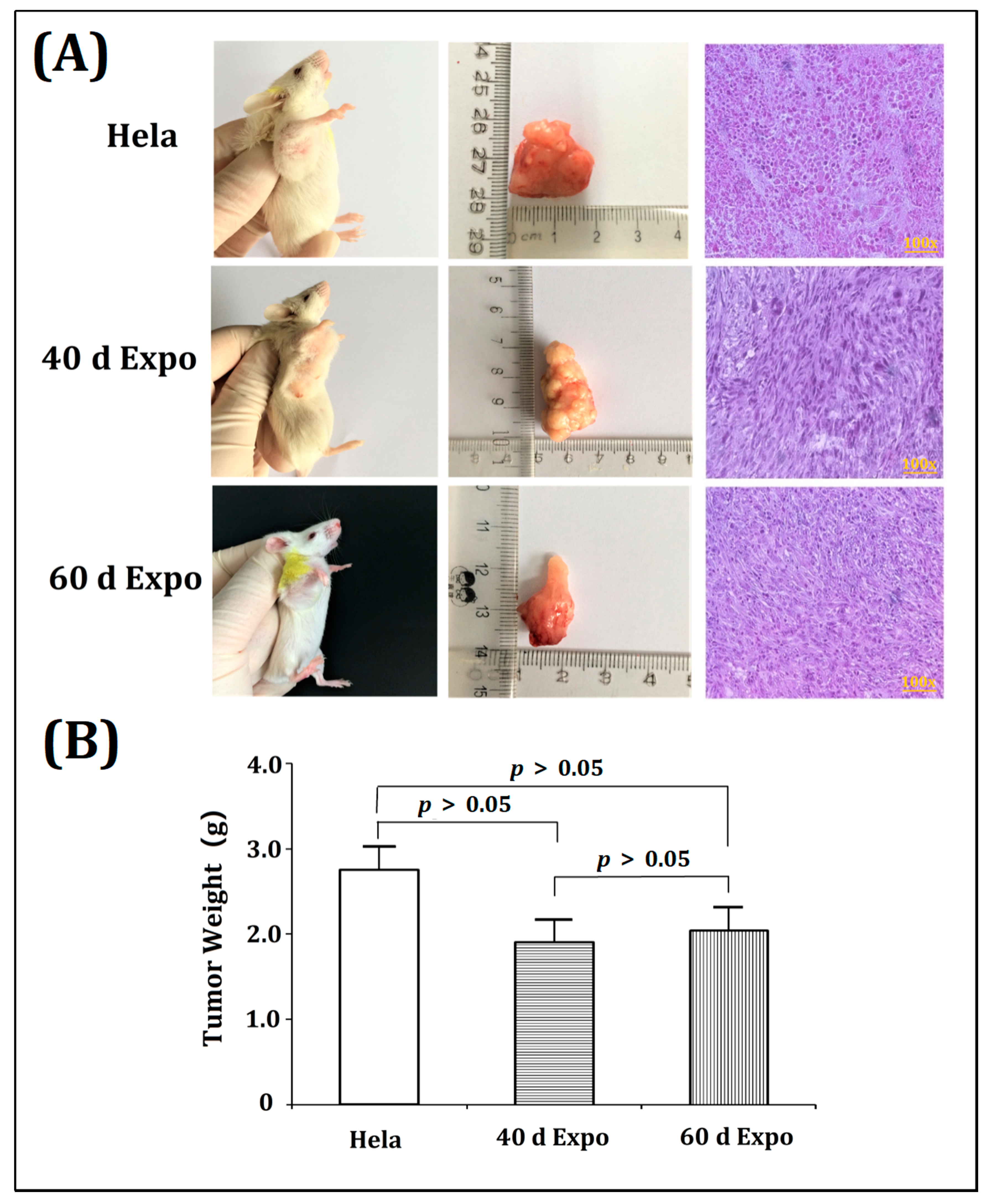
3.3. The 40-Day and 60-Day Exposed Balb/c-3T3 Cells Were Able to Form Tumors in SCID Mice
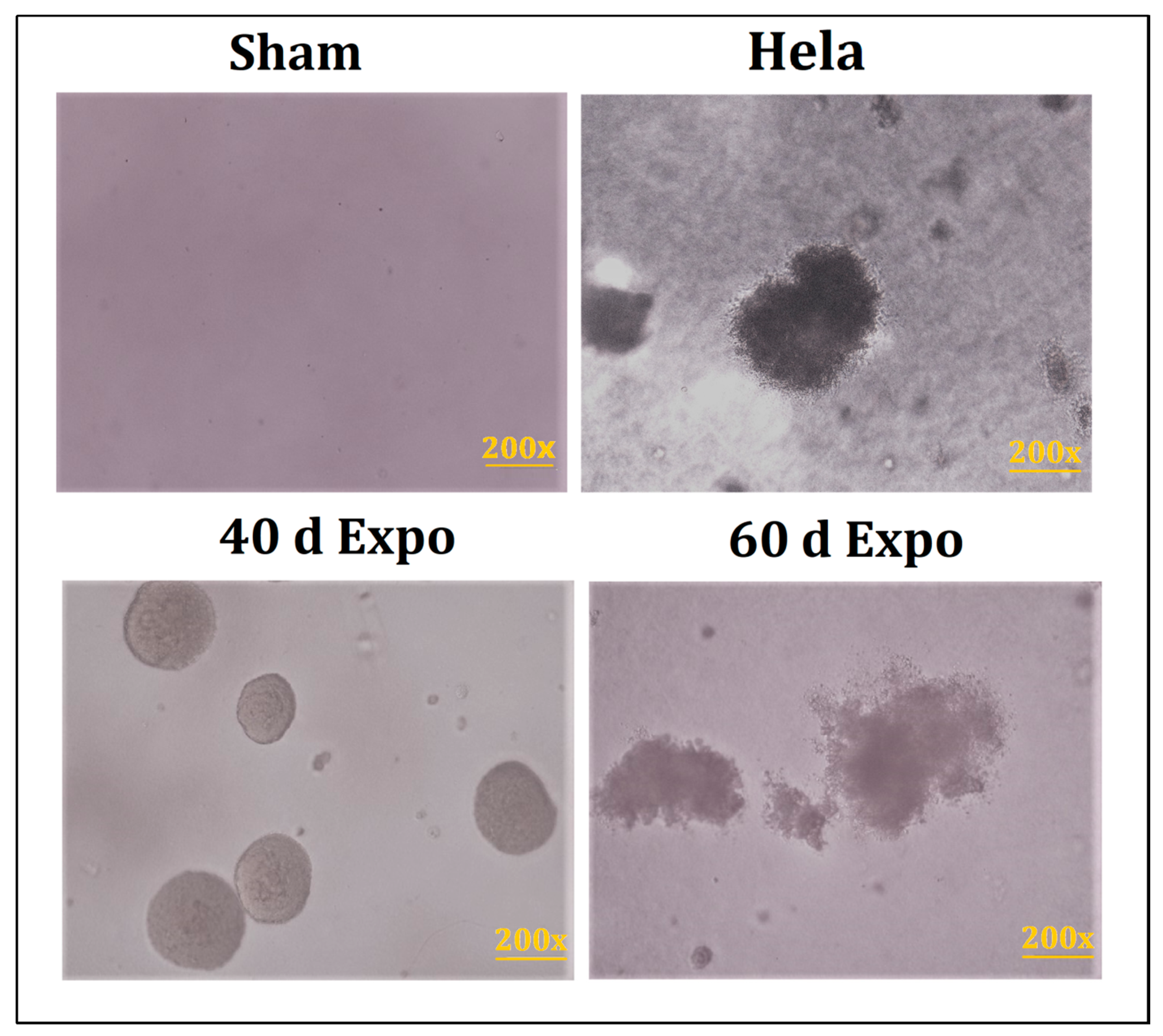
3.4. Clones Were Observed in 40-Day and 60-Day expo Balb/c-3T3 Cells
3.5. Significantly Expressed Genes Were Found by mRNA Microarray Detection
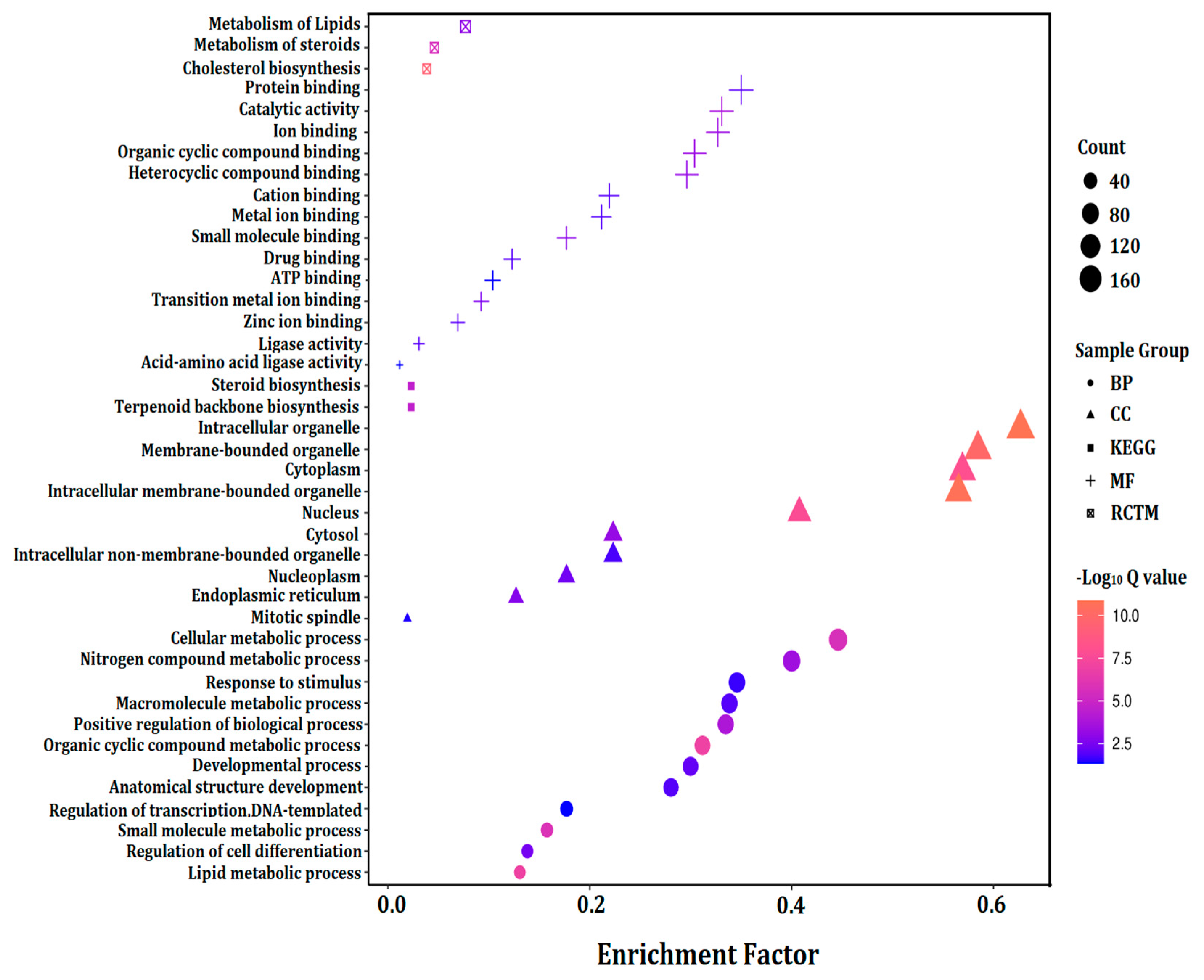
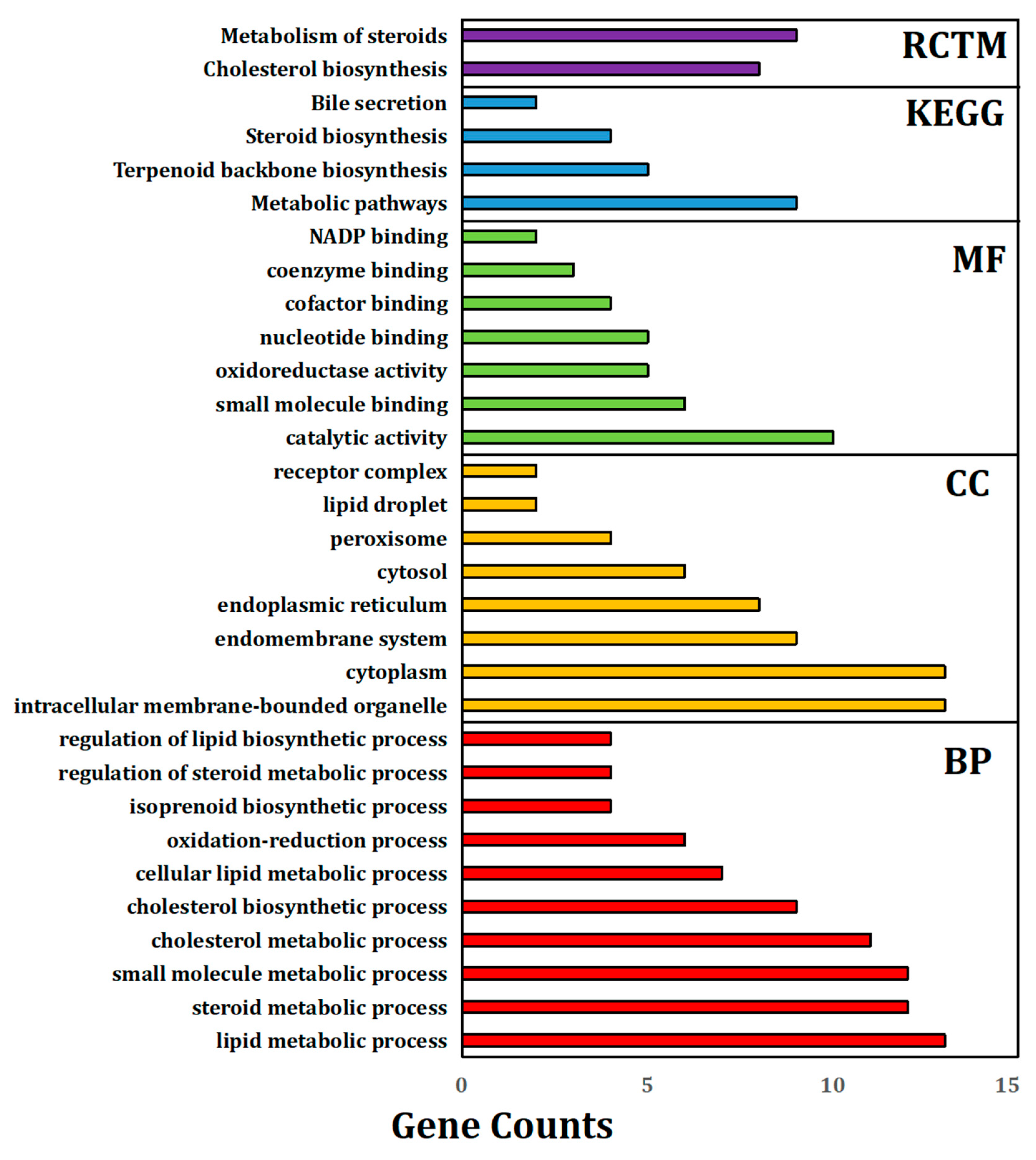
3.6. GO Term Enrichment Analysis of Significantly Expressed Genes (Fold Change ≥ 5)
3.7. Protein–Protein Interaction (PPI) Network Analysis
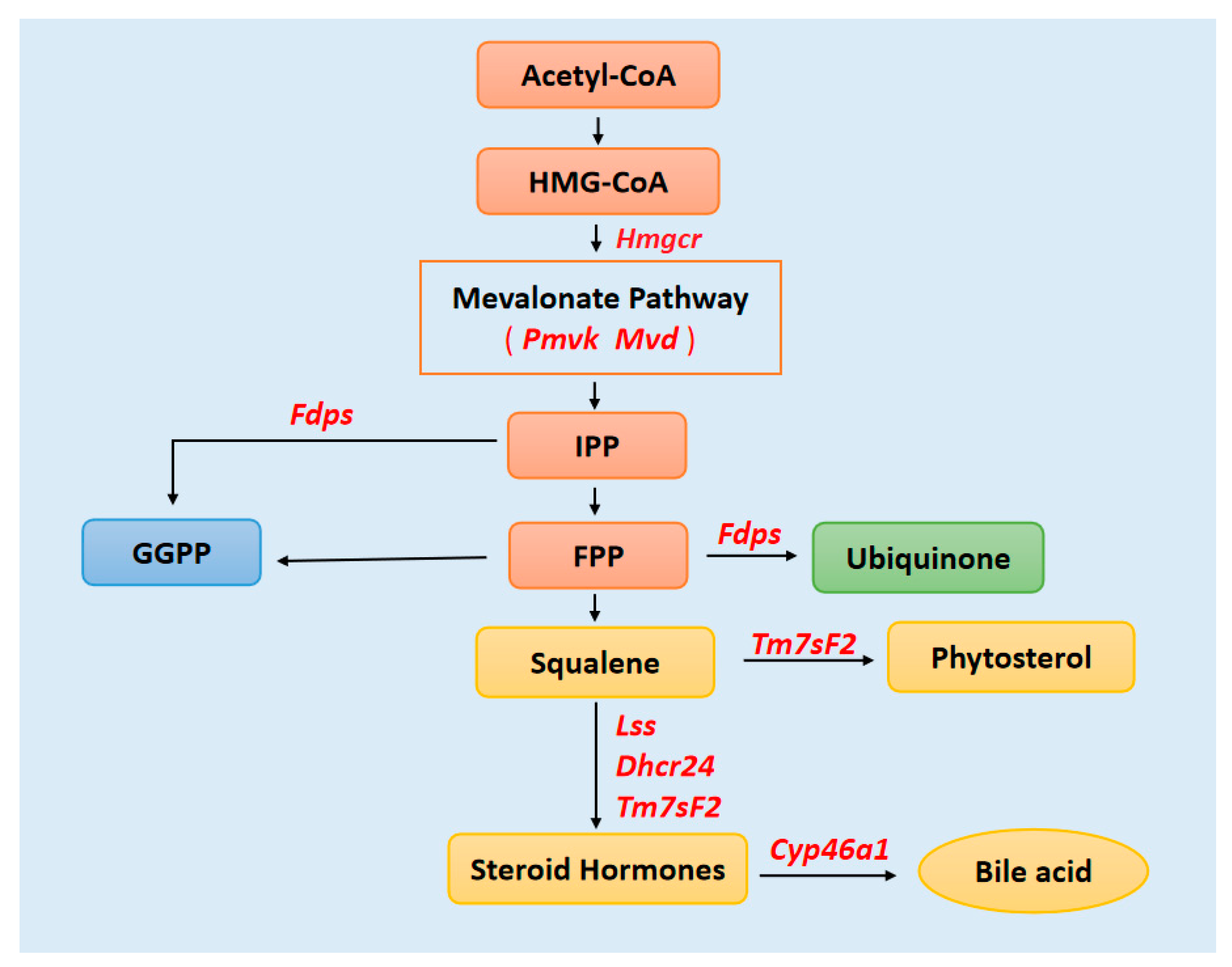
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Di Ciaula, A. Towards 5G communication systems: Are there health implications? Int. J. Hyg. Environ. Health 2018, 221, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, J.; Järås, J.; Henriksson, R.; Holgersson, G.; Bergström, S.; Estenberg, J.; Augustsson, T.; Bergqvist, M. No Evidence for Increased Brain Tumour Incidence in the Swedish National Cancer Register between Years 1980. Anticancer Res. 2019, 39, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, A.B.; Sears, M.E.; Morgan, L.L.; Davis, D.L.; Hardell, L.; Oremus, M.; Soskolne, C.L. Risks to Health and Well-Being from Ra-dio-Frequency Radiation Emitted by Cell Phones and Other Wireless Devices. Front. Public Health 2019, 7, 223. [Google Scholar] [CrossRef] [Green Version]
- Ostrom, Q.T.; Patil, N.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013–2017. Neuro-Oncology 2020, 22 (Suppl. 1), iv1–iv96. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011. Neuro-Oncology 2018, 20, iv1–iv86. [Google Scholar] [CrossRef] [Green Version]
- Philips, A.; Henshaw, D.L.; Lamburn, G.; O’Carroll, M.J. Brain Tumours: Rise in Glioblastoma Multiforme Incidence in England 1995–2015 Suggests an Adverse Environmental or Lifestyle Factor. J. Environ. Public Health 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hardell, L.; Nilsson, M.; Koppel, T.; Carlberg, M. Aspects on the International Commission on Non-Ionizing Radiation Protec-tion (ICNIRP) 2020 Guidelines on Radiofrequency Radiation. J. Cancer Sci. Clin. Ther. 2021, 5, 250–285. [Google Scholar] [CrossRef]
- Belpomme, D.; Hardell, L.; Belyaev, I.; Burgio, E.; Carpenter, D.O. Thermal and non-thermal health effects of low intensity non-ionizing radiation: An international perspective. Environ. Pollut. 2018, 242, 643–658. [Google Scholar] [CrossRef]
- Narayanan, S.N.; Mohapatra, N.; John, P. Radiofrequency electromagnetic radiation exposure effects on amygdala mor-phology, place preference behavior and brain caspase-3 activity in rats. Environ. Toxicol. Pharmacol. 2018, 58, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Eker, E.D.; Arslan, B.; Yildirim, M.; Akar, A.; Aras, N. The effect of exposure to 1800 MHz radiofrequency radiation on epidermal growth factor, caspase-3, Hsp27 and p38MAPK gene expressions in the rat eye. Bratisl. Med. J. 2018, 119, 588–592. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Mesharam, M.K.; Krishnamurthy, S. Electromagnetic radiation 2450 MHz exposure causes cognition deficit with mitochondrial dysfunction and activation of intrinsic pathway of apoptosis in rats. J. Biosci. 2018, 43, 263–276. [Google Scholar] [CrossRef]
- Li, Y.; Qu, X.; Wang, X.; Liu, M.; Wang, C.; Lv, Z.; Li, W.; Tao, T.; Song, D.; Liu, X. Microwave Radiation Injures Microvasculature Through Inducing Endoplasmic Reticulum Stress. Microcirculation 2014, 21, 490–498. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Bai, L.; Uchida, K.; Bai, W.; Wu, B.; Xu, W.; Zhu, H.; Huang, H. Effects of low frequency electromagnetic field on proliferation of human epidermal stem cells: An in vitro study. Bioelectromagnetic 2013, 34, 74–80. [Google Scholar] [CrossRef]
- Kumar, S.; Behari, J.; Sisodia, R. Influence of electromagnetic fields on reproductive system of male rats. Int. J. Radiat. Biol. 2013, 89, 147–154. [Google Scholar] [CrossRef]
- Chang, H.Y.; Shih, M.H.; Huang, H.C.; Tsai, S.R.; Juan, H.F.; Lee, S.C. Middle infrared radiation induces G2/M cell cycle arrest in A549 lung cancer cells. PLoS ONE 2013, 8, e54117. [Google Scholar] [CrossRef] [Green Version]
- Ahmadianpour, M.R.; Abdolmaleki, P.; Mowla, S.J.; Hosseinkhani, S. Static magnetic field of 6 mT induces apoptosis and alters cell cycle in p53 mutant Jurkat cells. Electromagn. Biol. Med. 2012, 32, 9–19. [Google Scholar] [CrossRef]
- Santini, S.J.; Cordone, V.; Falone, S.; Mijit, M.; Tatone, C.; Amicarelli, F.; Di Emidio, G. Role of Mitochondria in the Oxidative Stress Induced by Electromagnetic Fields: Focus on Reproductive Systems. Oxidative Med. Cell. Longev. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Hanc, H.; Kerimoğlu, G.; Mercantepe, T.; Odac, E. Changes in testicular morphology and oxidative stress biomarkers in 60-day-old Sprague Dawley rats following exposure to continuous 900-MHz electromagnetic field for 1 h a day throughout adolescence. Reprod Toxicol. 2018, 81, 71–78. [Google Scholar] [CrossRef]
- Fasshauer, M.; Krüwel, T.; Zapf, A.; Stahnke, V.C.; Rave-Fränk, M.; Staab, W.; Sohns, J.S.; Steinmetz, M.; Unterberg-Buchwald, C.; Schuster, A.; et al. Absence of DNA double-strand breaks in human peripheral blood mononuclear cells after 3 Tesla magnetic resonance imaging assessed by γH2AX flow cytometry. Eur. Radiol. 2017, 28, 1149–1156. [Google Scholar] [CrossRef]
- Pandey, N.; Giri, S. Melatonin attenuates radiofrequency radiation (900 MHz)-induced oxidative stress, DNA damage and cell cycle arrest in germ cells of male Swiss albino mice. Toxicol. Ind. Health 2018, 34, 315–327. [Google Scholar] [CrossRef]
- Zosangzuali, M.; Lalramdinpuii, M.; Jagetia, G.C. Impact of radiofrequency radiation on DNA damage and anti-oxidants in peripheral blood lymphocytes of humans residing in the vicinity of mobile phone base stations. Electromagn. Biol. Med. 2017, 36, 295–305. [Google Scholar]
- Duan, W.; Liu, C.; Zhang, L. Comparison of the genotoxic effects induced by 50 Hz extremely low-frequency electromag-netic fields and 1800 MHz radiofrequency electromagnetic fields in GC-2 cells. Radiat Res. 2015, 183, 305–314. [Google Scholar] [CrossRef]
- National Toxicology Program. NTP Technical Report on the Toxicology and Carcinogenesis Studies in Hsd: Sprague Dawley Sd Rats Exposed to Whole-Body Radio Frequency Radiation at a Frequency (900 MHz) and Modulations (GSM and CDMA) Used by Cell Phones. NTP TR. Available online: https://ntp.niehs.nih.gov/ntp/about_ntp/trpanel/2018/march/tr595peerdraft.pdf (accessed on 11 September 2021).
- Lönn, S.; Ahlbom, A.; Hall, P.; Feychting, M.; the Swedish Interphone Study Group. Long-Term Mobile Phone Use and Brain Tumor Risk. Am. J. Epidemiol. 2005, 161, 526–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Roe, S.L.; Wyde, M.E.; Stout, M.D.; Winters, J.W.; Hobbs, C.A.; Shepard, K.G.; Green, A.S.; Kissling, G.E.; Shockley, K.R.; Tice, R.R.; et al. Evaluation of the genotoxicity of cell phone radiofrequency radiation in male and female rats and mice following subchronic exposure. Environ. Mol. Mutagen. 2019, 61, 276–290. [Google Scholar] [CrossRef] [Green Version]
- Falcioni, L.; Bua, L.; Tibaldi, E.; Lauriola, M.; De Angelis, L.; Gnudi, F.; Mandrioli, D.; Manservigi, M.; Manservisi, F.; Manzoli, I.; et al. Report of final results regarding brain and heart tumors in Sprague-Dawley rats exposed from prenatal life until natural death to mobile phone radiofrequency field representative of a 1.8 GHz GSM base station environmental emission. Environ. Res. 2018, 165, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Xiao, S.; Chen, H.; Zhang, M.; Chen, Z.; Long, Y.; Gao, L.; He, J.; Ge, Y.; Yi, W.; et al. The receptor for activated protein kinase C promotes cell growth, invasion and migration in cervical cancer. Int. J. Oncol. 2017, 51, 1497–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.-H.; Kim, H.S.; Kim, R.N.; Jung, S.-Y.; Hong, B.S.; Kang, E.J.; Lee, H.-B.; Moon, H.-G.; Noh, D.-Y.; Han, W. NAD(P)-dependent steroid dehydrogenase-like is involved in breast cancer cell growth and metastasis. BMC Cancer 2020, 20, 375. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xie, J.; Liu, L.; Huang, W.; Han, Q.; Qin, J.; Liu, S.; Jiang, Z. NAD(P)-dependent steroid dehydrogenase-like protein and neutral cholesterol ester hydrolase 1 serve as novel markers for early detection of gastric cancer identified using quantitative proteomics. J. Clin. Lab. Anal. 2021, 35, e23652. [Google Scholar] [CrossRef]
- Fagone, P.; Jackowski, S. Membrane phospholipid synthesis and endoplasmic reticulum function. J. Lipid Res. 2009, 50, S311–S316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.S.; Nakano, A. Membrane Traffic Within the Golgi Apparatus. Annu. Rev. Cell Dev. Biol. 2009, 25, 113–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Bell, E.H.; Chakravarti, A. Lipid metabolism emerges as a promising target for malignant glioma therapy. CNS Oncol. 2013, 2, 289–299. [Google Scholar] [CrossRef]
- Röhrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Cancer 2016, 16, 732–749. [Google Scholar] [CrossRef]
- Zhao, J.; Zhi, Z.; Wang, C.; Xing, H.; Song, G.; Yu, X.; Zhu, Y.; Wang, X.; Zhang, X.; Di, Y. Exogenous lipids promote the growth of breast cancer cells via CD. Oncol. Rep. 2017, 38, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, G.; Avgustinova, A.; Mejetta, S.; Martín, M.; Castellanos, A.; Attolini, C.S.-O.; Berenguer, A.; Prats, N.; Toll, A.; Hueto, J.A.; et al. Targeting metastasis-initiating cells through the fatty acid receptor CD. Nature 2017, 541, 41–45. [Google Scholar] [CrossRef]
- Cheng, C.; Geng, F.; Cheng, X.; Guo, D. Lipid metabolism reprogramming and its potential targets in cancer. Cancer Commun. 2018, 38, 27. [Google Scholar] [CrossRef]
- Sharma, B.; Agnihotri, N. Role of cholesterol homeostasis and its efflux pathways in cancer progression. J. Steroid Biochem. Mol. Biol. 2019, 191, 105377. [Google Scholar] [CrossRef] [PubMed]
- Krycer, J.R.; Brown, A.J. Cholesterol accumulation in prostate cancer: A classic observation from a modern perspective. Biochim. Biophys. Acta 2013, 1835, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Freter, C. Lipid Metabolism, Apoptosis and Cancer Therapy. Int. J. Mol. Sci. 2015, 16, 924–949. [Google Scholar] [CrossRef] [Green Version]
- Chushi, L.; Wei, W.; Kangkang, X.; Yongzeng, F.; Ning, X.; Xiaolei, C. HMGCR is up-regulated in gastric cancer and promotes the growth and migration of the cancer cells. Gene 2016, 587, 42–47. [Google Scholar] [CrossRef]
- Chen, C.L.; Paul, L.N.; Mermoud, J.C.; Steussy, C.N.; Stauffacher, C.V. Visualizing the enzyme mechanism of mevalonate diphos-phate decarboxylase. Nat. Commun. 2020, 11, 3969. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Rachagani, S.; Muniyan, S.; Siddiqui, J.A.; Cruz, E.; Sharma, S.; Krishnan, R.; Killips, B.J.; Sheinin, Y.; Lele, S.M.; et al. FDPS cooperates with PTEN loss to promote prostate cancer progression through modulation of small GTPases/AKT axis. Oncogene 2019, 38, 5265–5280. [Google Scholar] [CrossRef]
- Wu, J.; Guo, L.; Qiu, X.; Ren, Y.; Li, F.; Cui, W.; Song, S. Genkwadaphnin inhibits growth and invasion in hepatocellular carcinoma by blocking DHCR24-mediated cholesterol biosynthesis and lipid rafts formation. Br. J. Cancer 2020, 123, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.-Z.; Yin, J.-B.; Yang, J.-J.; Cao, L. Regulatory factor X1 depresses ApoE-dependent Aβ uptake by miRNA-124 in microglial response to oxidative stress. Neuroscience 2017, 344, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, F. MicroRNA-758 inhibits tumorous behavior in tongue squamous cell carcinoma by directly targeting metadherin. Mol. Med. Rep. 2019, 19, 1883–1890. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; You, X. MicroRNA-758 inhibits malignant progression of retinoblastoma by directly targeting PAX. Oncol. Rep. 2018, 40, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhai, R.; Zhang, L.; Zhao, S. MicroRNA-149 suppresses the proliferation and increases the sensitivity of ovarian cancer cells to cisplatin by targeting X-linked inhibitor of apoptosis. Oncol. Lett. 2018, 15, 7328–7334. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Wang, H. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol. Cancer 2019, 18, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Z.; Xiang, X.; Li, J.; Wu, S. Molecular Mechanism of Malignant Transformation of Balb/c-3T3 Cells Induced by Long-Term Exposure to 1800 MHz Radiofrequency Electromagnetic Radiation (RF-EMR). Bioengineering 2022, 9, 43. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9020043
Ding Z, Xiang X, Li J, Wu S. Molecular Mechanism of Malignant Transformation of Balb/c-3T3 Cells Induced by Long-Term Exposure to 1800 MHz Radiofrequency Electromagnetic Radiation (RF-EMR). Bioengineering. 2022; 9(2):43. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9020043
Chicago/Turabian StyleDing, Zhen, Xiaoyong Xiang, Jintao Li, and Shuicai Wu. 2022. "Molecular Mechanism of Malignant Transformation of Balb/c-3T3 Cells Induced by Long-Term Exposure to 1800 MHz Radiofrequency Electromagnetic Radiation (RF-EMR)" Bioengineering 9, no. 2: 43. https://0-doi-org.brum.beds.ac.uk/10.3390/bioengineering9020043