
A New Perspective of Pseudomonas—Host Interactions: Distribution and Potential Ecological Functions of the Genus Pseudomonas within the Bark Beetle Holobiont

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
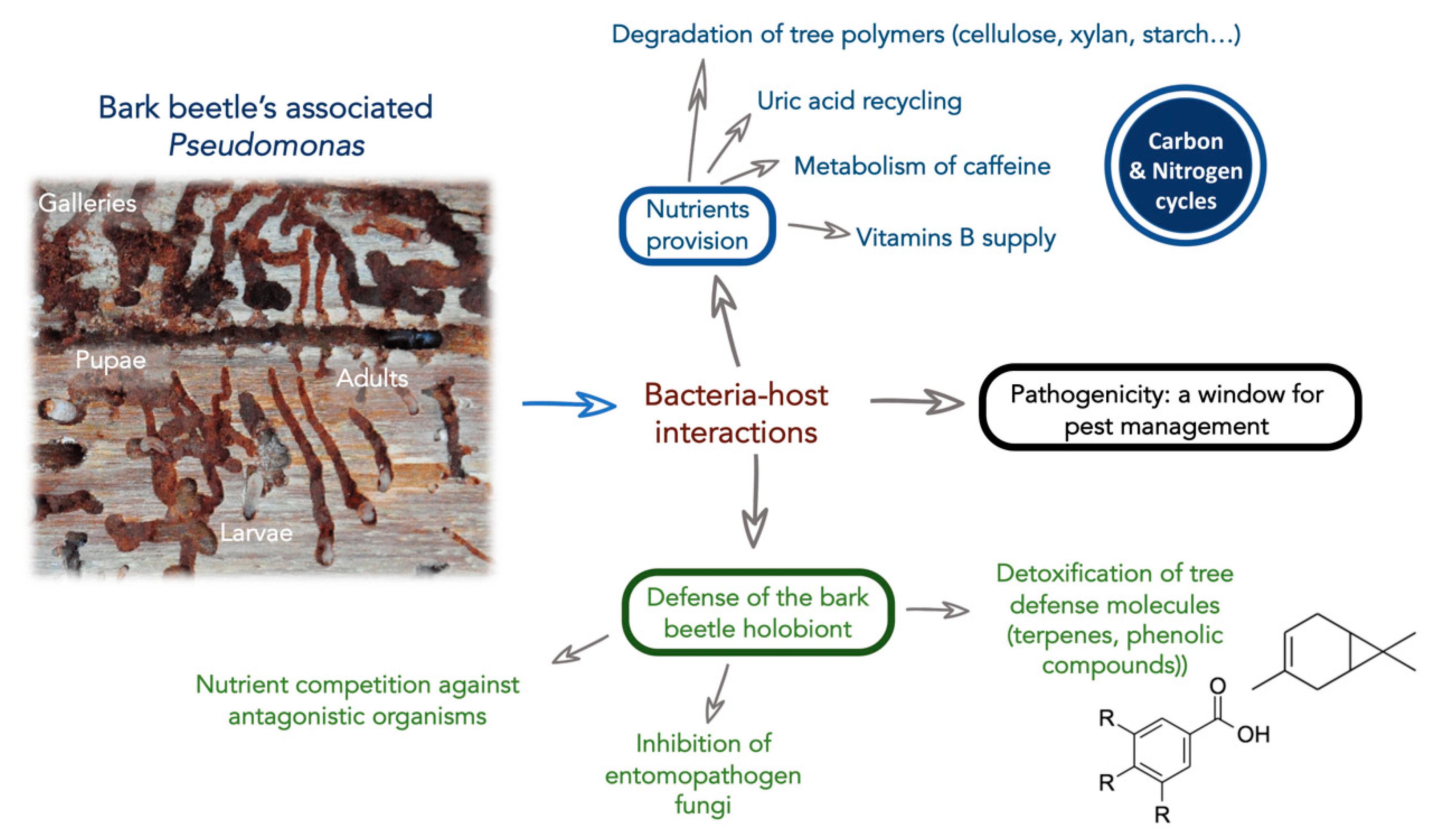
2. Pseudomonas Bacteria Are Broadly Associated to Bark Beetles
3. Role of Pseudomonas in the Nutrition of Bark Beetles
4. Role of Pseudomonas in the Detoxification of the Bark Beetle Environment and in Pheromone Production
5. Role of Pseudomonas in the Defense of the Bark Beetle Holobiont against Pathogens
6. Pathogenic Pseudomonas: Potential as Biocontrol Agents for Bark Beetle Pests
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of climate change for biotic disturbances in North American forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Biedermann, P.H.; Müller, J.; Grégoire, J.C.; Gruppe, A.; Hagge, J.; Hammerbacher, A.; Hofstetter, R.W.; Kandasamy, D.; Kolarik, M.; Kostovcik, M.; et al. Bark beetle population dynamics in the Anthropocene: Challenges and solutions. Trends Ecol. Evol. 2019, 34, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, B.S.; Raffa, K.F. Evolution of tree killing in bark beetles (Coleoptera: Curculionidae): Trade-offs between the maddening crowds and a sticky situation. Can. Entomol. 2013, 145, 471–495. [Google Scholar] [CrossRef] [Green Version]
- Raffa, K.F.; Gregoire, J.C.; Lindgren, B.S. Natural history and ecology of bark beetles. In Bark Beetles; Academic Press: Cambridge, MA, USA, 2015; pp. 1–40. [Google Scholar]
- Byers, J.A. Host-tree chemistry affecting colonization in bark beetles. In Chemical Ecology of Insects 2; Springer: Boston, MA, USA, 1995; pp. 154–213. [Google Scholar]
- Schmidt, K.; Engel, P. Mechanisms underlying gut microbiota–host interactions in insects. J. Exp. Biol. 2021, 224, jeb207696. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Roy, A. Microbial Influence on Plant–Insect Interaction. In Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology; Springer: Singapore, 2021; pp. 337–363. [Google Scholar]
- Weisskopf, L.; Schulz, S.; Garbeva, P. Microbial volatile organic compounds in intra-kingdom and inter-kingdom interactions. Nat. Rev. Microbiol. 2021, 1–14. [Google Scholar] [CrossRef]
- García-Fraile, P. Roles of bacteria in the bark beetle holobiont–how do they shape this forest pest? Ann. Appl. Biol. 2018, 172, 111–125. [Google Scholar] [CrossRef]
- Six, D.L. The bark beetle holobiont: Why microbes matter. J. Chem. Ecol. 2013, 39, 989–1002. [Google Scholar] [CrossRef]
- Fabryová, A.; Kostovčík, M.; Díez-Méndez, A.; Jiménez-Gómez, A.; Celador-Lera, L.; Saati-Santamaría, Z.; Sechovcová, H.; Menéndez, E.; Kolařik, M.; García-Fraile, P. On the bright side of a forest pest-the metabolic potential of bark beetles’ bacterial associates. Sci. Total Environ. 2018, 619, 9–17. [Google Scholar] [CrossRef]
- Boone, C.K.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.S.; Bohlmann, J.; Raffa, K.F. Bacteria associated with a tree-killing insect reduce concentrations of plant defense compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.J.; Oh, D.C.; Yuceer, M.C.; Klepzig, K.D.; Clardy, J.; Currie, C.R. Bacterial protection of beetle-fungus mutualism. Science 2008, 322, 63. [Google Scholar] [CrossRef] [Green Version]
- Veselská, T.; Skelton, J.; Kostovčík, M.; Hulcr, J.; Baldrian, P.; Chudíčková, M.; Cajthamlad, T.; Vojtováa, T.; Garcia-Fraile, P.; Kolařík, M. Adaptive traits of bark and ambrosia beetle-associated fungi. Fungal Ecol. 2019, 41, 165–176. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W. Comparative genomics of plant-associated Pseudomonas spp.: Insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef] [Green Version]
- Barbier, M.; Damron, F.H.; Bielecki, P.; Suárez-Diez, M.; Puchałka, J.; Albertí, S.; dos Santos, V.M.; Goldberg, J.B. From the environment to the host: Re-wiring of the transcriptome of Pseudomonas aeruginosa from 22 °C to 37 °C. PLoS ONE 2014, 9, e89941. [Google Scholar] [CrossRef]
- Crone, S.; Vives-Flórez, M.; Kvich, L.; Saunders, A.M.; Malone, M.; Nicolaisen, M.H.; Martínez-García, E.; Rojas-Acosta, C.; Catalina Gomez-Puerto, M.; Calum, H.; et al. The environmental occurrence of Pseudomonas aeruginosa. Apmis 2020, 128, 220–231. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Bakker, P.A. Interactions between plants and beneficial Pseudomonas spp.: Exploiting bacterial traits for crop protection. Antonie Van Leeuwenhoek 2007, 92, 367–389. [Google Scholar] [CrossRef]
- Xin, X.F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316. [Google Scholar] [CrossRef]
- Huszczynski, S.M.; Lam, J.S.; Khursigara, C.M. The Role of Pseudomonas aeruginosa Lipopolysaccharide in Bacterial Pathogenesis and Physiology. Pathogens 2020, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Dieppois, G.; Opota, O.; Lalucat, J.; Lemaitre, B. Pseudomonas entomophila: A versatile bacterium with entomopathogenic properties. In Pseudomonas; Springer: Dordrecht, The Netherlands, 2015; pp. 25–49. [Google Scholar]
- Kamarajan, B.P.; Muthusamy, A. Survival strategy of Pseudomonas aeruginosa on the nanopillar topography of dragonfly (Pantala flavescens) wing. AMB Express 2020, 10, 85. [Google Scholar] [CrossRef]
- Kim, H.R.; Lee, H.M.; Yu, H.C.; Jeon, E.; Lee, S.; Li, J.; Kim, D.H. Biodegradation of Polystyrene by Pseudomonas sp. Isolated from the Gut of Superworms (Larvae of Zophobas atratus). Environ. Sci. Technol. 2020, 54, 6987–6996. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Klepzig, K.D.; Raffa, K.F. Bacteria in oral secretions of an endophytic insect inhibit antagonistic fungi. Ecol. Entomol. 2006, 31, 636–645. [Google Scholar] [CrossRef]
- Morales-Jiménez, J.; Zúñiga, G.; Ramírez-Saad, H.C.; Hernández-Rodríguez, C. Gut-associated bacteria throughout the life cycle of the bark beetle Dendroctonus rhizophagus Thomas and Bright (Curculionidae: Scolytinae) and their cellulolytic activities. Microb. Ecol. 2012, 64, 268–278. [Google Scholar] [CrossRef]
- Durand, A.A.; Bergeron, A.; Constant, P.; Buffet, J.P.; Déziel, E.; Guertin, C. Surveying the endomicrobiome and ectomicrobiome of bark beetles: The case of Dendroctonus simplex. Sci. Rep. 2015, 5, 17190. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Lou, Q.; Wang, B.; Xu, L.; Cheng, C.; Lu, M.; Sun, J. Altered carbohydrates allocation by associated bacteria-fungi interactions in a bark beetle-microbe symbiosis. Sci. Rep. 2016, 6, 20135. [Google Scholar]
- Saati Santamaría, Z.; López-Mondéjar, R.; Jiménez Gómez, A.; Méndez, A.D.; Větrovský, T.; Igual, J.M.; Velázquez, E.; Kolarik, M.; Rivas, R.; García-Fraile, P. Discovery of phloeophagus beetles as a source of Pseudomonas strains that produce potentially new bioactive substances and description of Pseudomonas bohemica sp. nov. Front. Microbiol. 2018, 9, 913. [Google Scholar] [CrossRef] [Green Version]
- Mason, C.J.; Hanshew, A.S.; Raffa, K.F. Contributions by host trees and insect activity to bacterial communities in Dendroctonus valens (Coleoptera: Curculionidae) galleries, and their high overlap with other microbial assemblages of bark beetles. Environ. Entomol. 2016, 45, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Hulcr, J.; Rountree, N.R.; Diamond, S.E.; Stelinski, L.L.; Fierer, N.; Dunn, R.R. Mycangia of ambrosia beetles host communities of bacteria. Microb. Ecol. 2012, 64, 784–793. [Google Scholar] [CrossRef] [Green Version]
- Hernández-García, J.A.; Briones-Roblero, C.I.; Rivera-Orduña, F.N.; Zúñiga, G. Revealing the gut bacteriome of Dendroctonus bark beetles (Curculionidae: Scolytinae): Diversity, core members and co-evolutionary patterns. Sci. Rep. 2017, 7, 13864. [Google Scholar] [CrossRef]
- Ibarra-Juarez, L.A.; Burton, M.A.J.; Biedermann, P.H.W.; Cruz, L.; Desgarennes, D.; Ibarra-Laclette, E.; Latorre, A.; Alonso-Sánchez, A.; Villafan, E.; Hanako-Rosas, G.; et al. Evidence for Succession and Putative Metabolic Roles of Fungi and Bacteria in the Farming Mutualism of the Ambrosia Beetle Xyleborus affinis. mSystems 2020, 5, e00541-20. [Google Scholar] [CrossRef] [PubMed]
- Briones-Roblero, C.I.; Hernández-García, J.A.; Gonzalez-Escobedo, R.; Soto-Robles, L.V.; Rivera-Orduña, F.N.; Zúñiga, G. Structure and dynamics of the gut bacterial microbiota of the bark beetle, Dendroctonus rhizophagus (Curculionidae: Scolytinae) across their life stages. PLoS ONE 2017, 12, e0175470. [Google Scholar] [CrossRef]
- Xu, L.; Sun, L.; Zhang, S.; Wang, S.; Lu, M. High-Resolution Profiling of Gut Bacterial Communities in an Invasive Beetle using PacBio SMRT Sequencing System. Insects 2019, 10, 248. [Google Scholar] [CrossRef] [Green Version]
- Aylward, F.O.; Suen, G.; Biedermann, P.H.; Adams, A.S.; Scott, J.J.; Malfatti, S.A.; del Rio, T.G.; Tringe, S.G.; Poulsen, M.; Raffa, K.F.; et al. Convergent bacterial microbiotas in the fungal agricultural systems of insects. mBio 2014, 5, e02077-14. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Juarez, L.A.; Desgarennes, D.; Vázquez-Rosas-Landa, M.; Villafan, E.; Alonso-Sánchez, A.; Ferrera-Rodríguez, O.; Moya, A.; Carrillo, D.; Cruz, L.; Carrión, G.; et al. Impact of rearing conditions on the ambrosia beetle’s microbiome. Life 2018, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, A.S.; Aylward, F.O.; Adams, S.M.; Erbilgin, N.; Aukema, B.H.; Currie, C.R.; Suen, G.; Raffa, K.F. Mountain pine beetles colonizing historical and naive host trees are associated with a bacterial community highly enriched in genes contributing to terpene metabolism. Appl. Environ. Microbiol. 2013, 79, 3468–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briones-Roblero, C.I.; Rodríguez-Díaz, R.; Santiago-Cruz, J.A.; Zúñiga, G.; Rivera-Orduña, F.N. Degradation capacities of bacteria and yeasts isolated from the gut of Dendroctonus rhizophagus (Curculionidae: Scolytinae). Folia Microbiol. 2017, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Morales-Jiménez, J.; de León, A.V.P.; García-Domínguez, A.; Martínez-Romero, E.; Zúñiga, G.; Hernández-Rodríguez, C. Nitrogen-fixing and uricolytic bacteria associated with the gut of Dendroctonus rhizophagus and Dendroctonus valens (Curculionidae: Scolytinae). Microb. Ecol. 2013, 66, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Winder, R.S.; Macey, D.E.; Cortese, J. Dominant bacteria associated with broods of mountain pine beetle, Dendroctonus ponderosae (Coleoptera: Curculionidae, Scolytinae). J. Entomol. Soc. Br. Columbia 2010, 107, 43–56. [Google Scholar]
- Hu, X.; Yu, J.; Wang, C.; Chen, H. Cellulolytic bacteria associated with the gut of Dendroctonus armandi larvae (Coleoptera: Curculionidae: Scolytinae). Forests 2014, 5, 455–465. [Google Scholar] [CrossRef]
- Xu, L.; Lou, Q.; Cheng, C.; Lu, M.; Sun, J. Gut-associated bacteria of Dendroctonus valens and their involvement in verbenone production. Microb. Ecol. 2015, 70, 1012–1023. [Google Scholar] [CrossRef]
- Harrington, T.C. Ecology and evolution of mycophagous bark beetles and their fungal partners. In Insect-Fungal Associations. Ecology and Evolution; Oxford University Press: New York, NY, USA, 2005; pp. 257–291. [Google Scholar]
- Yilmax, H.; Sezen, K.; Kati, H.; Demirbağ, Z. The first study on the bacterial flora of the European spruce bark beetle, Dendroctonus micans (Coleoptera: Scolytidae). Biologia 2006, 61, 679–686. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Vasanthakumar, A.; Suazo, A.; Raffa, K.F. Survey and phylogenetic analysis of culturable microbes in the oral secretions of three bark beetle species. Entomol. Exp. Appl. 2009, 131, 138–147. [Google Scholar] [CrossRef]
- Kati, A.; Kati, H. Isolation and identification of bacteria from Xylosandrus germanus (Blandford) (Coleoptera: Curculionidae). Afr. J. Microbiol. Res. 2013, 7, 5288–5289. [Google Scholar]
- Adams, A.S.; Boone, C.K.; Bohlmann, J.; Raffa, K.F. Responses of bark beetle-associated bacteria to host monoterpenes and their relationship to insect life histories. J. Chem. Ecol. 2011, 37, 808–817. [Google Scholar] [CrossRef]
- Menéndez, E.; Ramírez-Bahena, M.H.; Fabryová, A.; Igual, J.M.; Benada, O.; Mateos, P.F.; Peix, A.; Kolarik, M.; García-Fraile, P. Pseudomonas coleopterorum sp. nov.; a cellulase-producing bacterium isolated from the bark beetle Hylesinus fraxini. Int. J. Syst. Evol. Microbiol. 2015, 65, 2852–2858. [Google Scholar] [CrossRef]
- Adams, A.S.; Currie, C.R.; Cardoza, Y.; Klepzig, K.D.; Raffa, K.F. Effects of symbiotic bacteria and tree chemistry on the growth and reproduction of bark beetle fungal symbionts. Can. J. For. Res. 2009, 39, 1133–1147. [Google Scholar] [CrossRef]
- Canganella, F.; Paparatti, B.; Natali, V. Microbial species isolated from the bark beetle Anisandrus dispar F. Microbiol. Res. 1994, 149, 123–128. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, F.; Wang, B.; Xu, D.; Cao, Q.; Lu, M.; Sun, J. Volatiles produced by bacteria alleviate antagonistic effects of one associated fungus on Dendroctonus valens larvae. Sci. China Life Sci. 2017, 60, 924–926. [Google Scholar] [CrossRef]
- Zhou, F.; Xu, L.; Wang, S.; Wang, B.; Lou, Q.; Lu, M.; Sun, J. Bacterial volatile ammonia regulates the consumption sequence of D-pinitol and D-glucose in a fungus associated with an invasive bark beetle. ISME J. 2017, 11, 2809–2820. [Google Scholar] [CrossRef]
- Therrien, J.; Mason, C.J.; Cale, J.A.; Adams, A.; Aukema, B.H.; Currie, C.R.; Raffa, K.F.; Erbilgin, N. Bacteria influence mountain pine beetle brood development through interactions with symbiotic and antagonistic fungi: Implications for climate-driven host range expansion. Oecologia 2015, 179, 467–485. [Google Scholar] [CrossRef]
- Delalibera, I.; Vasanthakumar, A.; Klepzig, K.D.; Raffa, K.F. Composition of the bacterial community in the gut of the pine engraver, Ips pini (Say)(Coleoptera) colonizing red pine. Symbiosis 2007, 43, 97–104. [Google Scholar]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.; Ashraf, M.Z.; Modlinger, R.; Synek, J.; Schlyter, F.; Roy, A. Unravelling the gut bacteriome of Ips (Coleoptera: Curculionidae: Scolytinae): Identifying core bacterial assemblage and their ecological relevance. Sci. Rep. 2020, 10, 18572. [Google Scholar] [CrossRef]
- Vasanthakumar, A.; Delalibera, I., Jr.; Handelsman, J.; Klepzig, K.D.; Schloss, P.D.; Raffa, K.F. Characterization of gut-associated bacteria in larvae and adults of the southern pine beetle, Dendroctonus frontalis Zimmermann. Environ. Entomol. 2006, 35, 1710–1717. [Google Scholar] [CrossRef]
- Dohet, L.; Grégoire, J.C.; Berasategui, A.; Kaltenpoth, M.; Biedermann, P.H. Bacterial and fungal symbionts of parasitic Dendroctonus bark beetles. FEMS Microbiol. Ecol. 2016, 92, fiw129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevim, A.; Gökçe, C.; Erbaş, Z.; Özkan, F. Bacteria from Ips sexdentatus (Coleoptera: Curculionidae) and their biocontrol potential. J. Basic Microbiol. 2012, 52, 695–704. [Google Scholar] [CrossRef]
- Sezen, K.; Kati, H.; Nalcacioĝlu, R.; Muratoĝlu, H.; Demirbaĝ, Z. Identification and pathogenicity of bacteria from European shot-hole borer, Xyleborus dispar Fabricius (Coleoptera: Scolytidae). Ann. Microbiol. 2008, 58, 173–179. [Google Scholar] [CrossRef]
- Peral-Aranega, E.; Saati-Santamaría, Z.; Kolařik, M.; Rivas, R.; García-Fraile, P. Bacteria Belonging to Pseudomonas typographi sp. nov. from the Bark Beetle Ips typographus Have Genomic Potential to Aid in the Host Ecology. Insects 2020, 11, 593. [Google Scholar] [CrossRef] [PubMed]
- Quince, C.; Walker, A.W.; Simpson, J.T.; Loman, N.J.; Segata, N. Shotgun metagenomics, from sampling to analysis. Nat. Biotechnol. 2017, 35, 833–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Mondéjar, R.; Kostovčík, M.; Lladó, S.; Carro, L.; García-Fraile, P. Exploring the plant microbiome through multi-omics approaches. In Probiotics in Agroecosystem; Springer: Singapore, 2017; pp. 233–268. [Google Scholar]
- Poveda, J.; Jiménez-Gómez, A.; Saati-Santamaría, Z.; Usategui-Martín, R.; Rivas, R.; García-Fraile, P. Mealworm frass as a potential biofertilizer and abiotic stress tolerance-inductor in plants. Appl. Soil Ecol. 2019, 142, 110–122. [Google Scholar] [CrossRef]
- Douglas, G.M.; Beiko, R.G.; Langille, M.G. Predicting the functional potential of the microbiome from marker genes using PICRUSt. In Microbiome Analysis; Humana Press: New York, NY, USA, 2018; pp. 169–177. [Google Scholar]
- Winand, R.; Bogaerts, B.; Hoffman, S.; Lefevre, L.; Delvoye, M.; Van Braekel, J.; Fu, Q.; Roosens, N.H.; De Keersmaecker, S.C.; Vanneste, K. Targeting the 16s rRNA gene for bacterial identification in complex mixed samples: Comparative evaluation of second (illumina) and third (oxford nanopore technologies) generation sequencing technologies. Int. J. Mol. Sci. 2020, 21, 298. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Gómez, A.; Saati-Santamaría, Z.; Kostovcik, M.; Rivas, R.; Velázquez, E.; Mateos, P.F.; Menéndez, E.; García-Fraile, P. Selection of the Root Endophyte Pseudomonas brassicacearum CDVBN10 as Plant Growth Promoter for Brassica napus L. Crops. Agronomy 2020, 10, 1788. [Google Scholar] [CrossRef]
- Sauvard, D. General biology of bark beetles. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Springer: Dordrecht, The Netherlands, 2007; pp. 63–88. [Google Scholar]
- Vega, F.E.; Hofstetter, R.W. (Eds.) Bark Beetles: Biology and Ecology of Native and Invasive Species; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Hofstetter, R.W.; Dinkins-Bookwalter, J.; Davis, T.S.; Klepzig, K.D. Symbiotic associations of bark beetles. In Bark Beetles; Academic Press: Cambridge, MA, USA, 2015; pp. 209–245. [Google Scholar]
- Saha, B.C. Hemicellulose bioconversion. J. Ind. Microbiol. 2003, 30, 279–291. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [Green Version]
- Carro, L.; Menéndez, E. Knock, knock-let the bacteria. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Academic Press: Cambridge, MA, USA, 2020; pp. 169–178. [Google Scholar]
- Deryło, M.; Skorupska, A. Enhancement of symbiotic nitrogen fixation by vitamin-secreting fluorescent Pseudomonas. Plant Soil 1993, 154, 211–217. [Google Scholar] [CrossRef]
- Marek-Kozaczuk, M.; Skorupska, A. Production of B-group vitamins by plant growth-promoting Pseudomonas fluorescens strain 267 and the importance of vitamins in the colonization and nodulation of red clover. Biol. Fertil. Soils 2001, 33, 146–151. [Google Scholar] [CrossRef]
- Martens, J.H.; Barg, H.; Warren, M.A.; Jahn, D. Microbial production of vitamin B 12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Ansari, Z.A.; Iqbal, F.; Akram, M. Microbial production of vitamin B12 by methanol utilizing strain of Pseudomonas specie. Pak. Biochem. Mol. Biol. 2007, 40, 5–10. [Google Scholar]
- Xia, W.; Chen, W.; Peng, W.F.; Li, K.T. Industrial vitamin B 12 production by Pseudomonas denitrificans using maltose syrup and corn steep liquor as the cost-effective fermentation substrates. Bioprocess Biosyst. Eng. 2015, 38, 1065–1073. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Figueroa-Teran, R.; Aw, M.; Song, M.; Gorzalski, A.; Abbott, N.L.; Chang, E.; Tittiger, C. Pheromone production in bark beetles. Insect Biochem. Mol. Biol. 2010, 40, 699–712. [Google Scholar] [CrossRef]
- Xu, L.T.; Lu, M.; Sun, J.H. Invasive bark beetle-associated microbes degrade a host defensive monoterpene. Insect Sci. 2016, 23, 183–190. [Google Scholar] [CrossRef]
- Martin, V.J.; Yu, Z.; Mohn, W.W. Recent advances in understanding resin acid biodegradation: Microbial diversity and metabolism. Arch. Microbiol. 1999, 172, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Kolařík, M.; Kostovčík, M.; Pažoutová, S. Host range and diversity of the genus Geosmithia (Ascomycota: Hypocreales) living in association with bark beetles in the Mediterranean area. Mycol. Res. 2007, 111, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Kolařík, M.; Kubátová, A.; Hulcr, J.; Pažoutová, S. Geosmithia fungi are highly diverse and consistent bark beetle associates: Evidence from their community structure in temperate Europe. Microb. Ecol. 2008, 55, 65–80. [Google Scholar] [CrossRef] [PubMed]
- González-Dominici, L.I.; Saati-Santamaría, Z.; García-Fraile, P. Genome Analysis and Genomic Comparison of the Novel Species Arthrobacter ipsi Reveal Its Potential Protective Role in Its Bark Beetle Host. Microb. Ecol. 2020, 1–12. [Google Scholar] [CrossRef]
- Lee, J.H.; Ma, K.C.; Ko, S.J.; Kang, B.R.; Kim, I.S.; Kim, Y.C. Nematicidal activity of a nonpathogenic biocontrol bacterium, Pseudomonas chlororaphis O6. Curr. Microbiol. 2011, 62, 746–751. [Google Scholar] [CrossRef]
- Popa, V.; Déziel, E.; Lavallée, R.; Bauce, E.; Guertin, C. The complex symbiotic relationships of bark beetles with microorganisms: A potential practical approach for biological control in forestry. Pest Manag. Sci. 2012, 68, 963–975. [Google Scholar] [CrossRef]
- Vodovar, N.; Vinals, M.; Liehl, P.; Basset, A.; Degrouard, J.; Spellman, P.; Boccard, F.; Lemaitre, B. Drosophila host defense after oral infection by an entomopathogenic Pseudomonas species. Proc. Natl. Acad. Sci. USA 2005, 102, 11414–11419. [Google Scholar] [CrossRef] [Green Version]
- Otsu, Y.; Matsuda, Y.; Mori, H.; Ueki, H.; Nakajima, T.; Fujiwara, K.; Matsumoto, M.; Azuma, N.; Kakutani, K.; Nonomura, T.; et al. Stable phylloplane colonization by entomopathogenic bacterium Pseudomonas fluorescens KPM-018P and biological control of phytophagous ladybird beetles Epilachna vigintioctopunctata (Coleoptera: Coccinellidae). Biocontrol Sci. Technol. 2004, 14, 427–439. [Google Scholar] [CrossRef]


{kind=link}
| Beetle (Tree-Host) | Sample | Life Stage | MD | Functions of Pseudomonas | Reference |
|---|---|---|---|---|---|
| Dendroctonus adjunctus (Pinus hartwegii), D. approximatus (P. teocote), D. jeffreyi (P. jeffreyi), D. mesoamericanus (P. teocote), D. mexicanus (P. patula), D. parallelocollis (P. hartwegii), D. ponderosae (P. strobiformis), D. pseudotsugae (Pseudotsuga menziesii var. glauca), D. rhizophagus (P. arizonica), D. valens (P. leiophylla), D. vitei (P. pseudostrobus) | G | - | CIA | - | [32] |
| D. rhizophagus (P. arizonica) | G | L, Pu, TA, PA, EA | CIA | - | [34] |
| D. simplex (Larix × eurolepis) | Cuticle, Ga | A | CIA | - | [27] |
| D. valens (P. resinosa) | Ga | - | CIA | - | [27] |
| D. valens (P. tabuliformis) | G | A | CIA | Prediction of metabolic activities using the PICRUSt method. | [35] |
| Xyleborinus saxeseni, D. ponderosae, D. frontalis | B, Ga | L, A | CIACIS | They found that Pseudomonas genome bins reconstructed from different insects are very similar (based on Average Nucleotide Identity (ANI) comparison). | [36] |
| Xyleborus affinis, Xyleborus bispinatus, and Xyleborus volvulus (artificially reared) | B | A | CIA | Prediction of metabolic activities using the PICRUSt method. | [37] |
| Xyleborus affinis (artificially reared) | B | A | CIA | Prediction of metabolic activities using the PICRUSt method. | [33] |
| D. pondersoae (P. contorta and P. contorta—P. banksiana hybrid) | B, Ga | A | CIS | Genetic potential for terpene and diterpene degradation. | [38] |
| D. rhizophagus (P. arizonica) | G | L, A | CD | Lipolytic, amylolytic, esterase, cellulolytic and xylanolytic activities. | [39] |
| D. ponderosae (Pinus sp.) | B, Ga, P | A | CD | In vitro degradation of monoterpenes. | [12] |
| D. valens (P. montezumae or P. leiophylla) | G | - | CD | Uricolytic activity. Ability to use uric acid as sole nitrogen, carbon and energy source. | [40] |
| D. ponderosae (P. contorta) | S | L, A | CD | - | [41] |
| D. armandi (P. armandii) | G | L | CD | Cellulolytic activity. | [42] |
| D. valens (Pinus sp.) | G, F | A | CD | Verbenone production, α-pinene degradation. | [43,44] |
| D. micans (Picea orientalis) | B | L, A | CD | - | [45] |
| D. rufipennis (Picea sp.), D. ponderosae (P. contorta) | OS | A | CD | - | [46] |
| Anisandrus (=Xyleborus) dispar (Corylus sp.) | B | L | CD | - | [47] |
| D. valens (P. contorta and P. contorta—P. banksiana hybrids) | B | A | CD | Ability to grow in presence of monoterpenes. | [48] |
| Hylesinus fraxini (Fraxinus excelsior) | B | A | CD | Cellulolytic activity. | [49] |
| Cryphalus piceae (Abies alba), Ips typographus (Picea abies) | B | A, L | CD | Degradation of cellullose, xylan, starch, and diverse chemical dyes. | [13] |
| D. rufipennis | OS | A | CD | Inhibit fungi associated with mouthparts of the beetle. | [25] |
| D. ponderosae (P. contorta) | OS | L, A | CD | Promote the growth of some beetle-associated fungi and suppress the growth of some other beetle-associated fungi. | [50] |
| D. valens (P. tabuliformis) | S, F, G | L | CD | Inoculation of different strains of Pseudomonas alleviate the antagonistic effects of fungi L. procerum and O. minus on larva growth. They show a direct inhibition of O. minus growth. They show that the D-pinitol and D-glucose consumption of O. minus is decreased, while the D-pinitol consumption of L. procerum is increased when bacteria is inoculated. | [28] |
| I. sexdentatus (P. orientalis) | B | A | CD | One isolate increase significatively the mortality when is inoculated into larvae.Another one do not increase significatively the mortality. | [51] |
| D. valens | - | - | CD | Pseudomonas volatiles decrease the growth of O. minus and, therefore, to increase the growth of larvae when O. minus is present. | [52] |
| D. valens | - | - | CD | Pseudomonas volatiles decrease the growth of L. procerum and, therefore, to increase the growth of larvae when L. procerum is present. | [53] |
| D. ponderosae | B, Ga | - | CD | Inoculation of different strains of Pseudomonas in P. contorta and in P. banksiana, reduced antagonistic effects by Aspergillus and Trichoderma resulting in more larvae and longer ovipositional and larval Ga. | [54] |
| Ips acuminatus (P. sylvestris), Pityogenes bidentatus (P. sylvestris) | B | A | CD | Ability to inhibit other microorganisms and genetic potential to produce antimicrobial compounds. | [29] |
| Ips pini (P. resinosa) | G | P, A | CD | - | [55] |
| Hypothenemus hampei (Coffea arabica, Coffea arabica var limani, C. congensis × C. canephora), H. eruditus (Cecropia sp.), Scolytodes maurus (Cecropia sp.) | B | - | CD CIA | In vitro and in vivo caffeine-degradation (use as sole source of C and N). | [56] |
| Ips duplicatus, I. typographus and Polygraphus poligraphus (Picea abies), I. acuminatus and I. sexdentatus (Pinus sylvestris), I. cembrae (Larix decidua). | G | A | CIA | Prediction of metabolic activities using the PICRUSt method. | [57] |
| D. frontalis (P. taeda) | G | A | CD CIA | - | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saati-Santamaría, Z.; Rivas, R.; Kolařik, M.; García-Fraile, P. A New Perspective of Pseudomonas—Host Interactions: Distribution and Potential Ecological Functions of the Genus Pseudomonas within the Bark Beetle Holobiont. Biology 2021, 10, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10020164
Saati-Santamaría Z, Rivas R, Kolařik M, García-Fraile P. A New Perspective of Pseudomonas—Host Interactions: Distribution and Potential Ecological Functions of the Genus Pseudomonas within the Bark Beetle Holobiont. Biology. 2021; 10(2):164. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10020164
Chicago/Turabian StyleSaati-Santamaría, Zaki, Raúl Rivas, Miroslav Kolařik, and Paula García-Fraile. 2021. "A New Perspective of Pseudomonas—Host Interactions: Distribution and Potential Ecological Functions of the Genus Pseudomonas within the Bark Beetle Holobiont" Biology 10, no. 2: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10020164