
A Heterozygous Genotype-Dependent Branched-Spike Wheat and the Potential Genetic Mechanism Revealed by Transcriptome Sequencing


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Evaluation of Agronomic Traits and FHB Resistance of the BSL
2.2. Genetic Pattern Analysis
2.3. Transcriptome Sequencing and DEGs Analysis
2.4. Quantitative Real-Time PCR (qRT-PCR) Analysis
3. Results
3.1. The BSL Plant Is a Naturally Occurring Mutant
3.2. The Genetic Patterns of the BSL Trait
3.3. The BSL Mutant Significantly Differed from the NSL Plants in Agronomic Traits and FHB Resistance
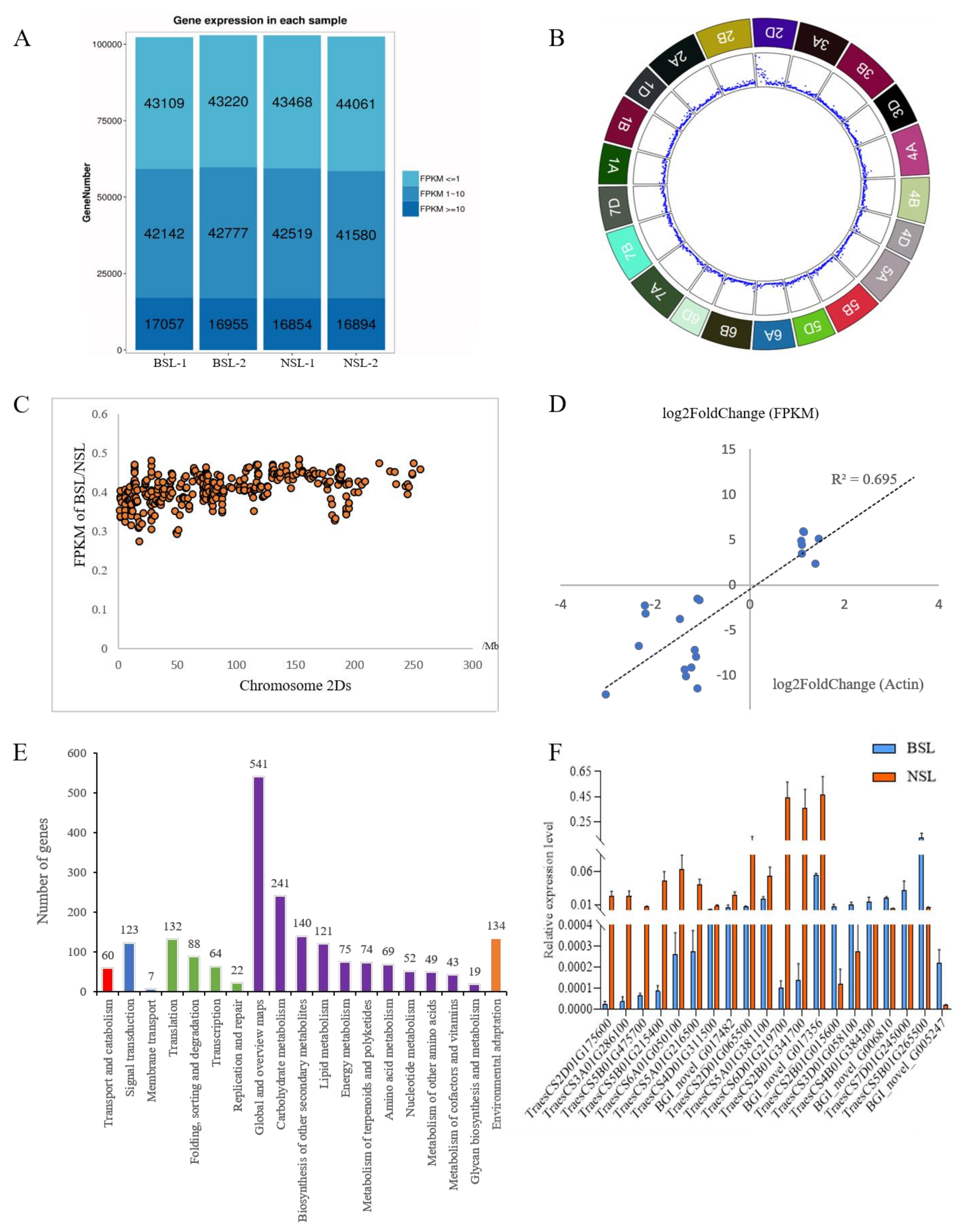
3.4. DEGs between the BSL Plants and the NSL Plants
3.5. Carbohydrate Metabolism Showed a Significant Difference between the BSL and the NSL Plants
4. Discussion
4.1. The Gene(s) Responsible for the BSL Trait May Have Undergone Negatively Artificial Selection
4.2. Gametophytic Male Sterility Putatively Contributed to the Heterozygosity of the BSL Trait
4.3. Wfzp Was Not the Causal Gene for the BSL Mutant
4.4. Carbohydrate Metabolism Showed a Significant Difference between the BSL and the NSL Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BSL | Branched spikelets |
| NSL | Normal spikelets |
| FHB | Fusarium head blight |
| PSS | Proportion of symptomatic spikeletes |
| DEGs | Differentially expressed genes |
| WFZP | Wheat FRIZZY PANICLE |
| CAPS/dCAPS | Derived/cleaved amplified polymorphic sequences |
| CM | Carbohydrate metabolism |
| GO | Gene Ontology |
References
- Eversole, K.; Feuillet, C.; Mayer, K.F.X.; Rogers, J. Slicing the wheat genome. Science 2014, 345, 285–287. [Google Scholar] [CrossRef] [Green Version]
- Poursarebani, N.; Seidensticker, T.; Koppolu, R.; Trautewig, C.; Gawronski, P.; Bini, F.; Govind, G.; Rutten, T.; Sakuma, S.; Tagiri, A. The genetic basis of composite spike form in barley and ‘Miracle-Wheat’. Genetics 2015, 201, 155–165. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Schnurbusch, T. A genetic playground for enhancing grain number in cereals. Trends Plant Sci. 2012, 17, 91–101. [Google Scholar] [CrossRef]
- Tanaka, W.; Pautler, M.; Jackson, D.; Hirano, H.Y. Grass meristems II: Inflorescence architecture, flower development and meristem fate. Plant Cell Physiol. 2013, 54, 313–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Yuan, Z. Molecular control of grass inflorescence development. Annu. Rev. Plant Biol. 2014, 65, 553–578. [Google Scholar] [CrossRef]
- Wei, Y. The regulation of phytohormone in determination of fertile floret number in wheat. J. Sichuan Agric. Univ. 1998, 16, 289–293. [Google Scholar]
- Wei, S.H.; Peng, Z.S.; Zhou, Y.H.; Yang, Z.J.; Wu, K.; Ouyang, Z.M. Nucleotide diversity and molecular evolution of the WAG-2 gene in common wheat (Triticum aestivum L) and its relatives. Genet. Mol. Biol. 2011, 34, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Shaman, B.C. Branched heads in wheat and wheat hybrids. Nature 1944, 153, 497–498. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, F.; Yan, L. Branching shoots and spikes from lateral meristems in bread wheat. PLoS ONE 2016, 11, e0151656. [Google Scholar] [CrossRef]
- Ritter, M.K.; Padilla, C.M.; Schmidt, R.J. The maize mutant barren stalk1 is defective in axillary meristem development. Am. J. Bot. 2002, 89, 203–210. [Google Scholar] [CrossRef]
- Gallavotti, A.; Zhao, Q.; Kyozuka, J.; Meeley, R.B.; Ritter, M.K.; Doebley, J.F.; Pe, M.E.; Schmidt, R.J. The role of barren stalk1 in the architecture of maize. Nature 2004, 432, 630–635. [Google Scholar] [CrossRef]
- Hirano, H.Y.; Tanaka, W.; Toriba, T. Grass flower development. Methods Mol. Biol. 2014, 1110, 57–84. [Google Scholar]
- Jiang, Y.; Cai, Z.; Xie, W.; Long, T.; Yu, H.; Zhang, Q. Rice functional genomics research: Progress and implications for crop genetic improvement. Biotechnol. Adv. 2012, 30, 1059–1070. [Google Scholar] [CrossRef]
- Pennell, A.L.; Halloran, G.M. Inheritance of supernumerary spikelets in wheat. Euphytica 1983, 32, 767–776. [Google Scholar] [CrossRef]
- Peng, Z.S.; Yen, C.; Yang, J.L. Chromosomal location of genes for supernumerary spikelet in bread wheat. Euphytica 1998, 103, 109–114. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audic, S.; Claverie, J.M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]
- Jaina, M.; Finn, R.D.; Eddy, S.R.; Alex, B. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, J.; Xue, X.; Li, C.; Yang, Z.; Li, T. CAPS/dCAPS designer: A web-based high-throughput dCAPS marker design tool. Sci. China Life Sci. 2018, 61, 992–995. [Google Scholar] [CrossRef]
- Li, L.; Shi, X.; Zheng, F.; Li, C.; Wu, D.; Bai, G.; Gao, D.; Wu, J.; Li, T. A novel nitrogen-dependent gene associates with the lesion mimic trait in wheat. Theor. Appl. Genet. 2016, 129, 2075–2084. [Google Scholar] [CrossRef]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Bai, G. A deletion mutation in TaHRC confers Fhb1 resistance to fusarium head blight in wheat. Nat. Genet. 2019, 51, 1–7. [Google Scholar] [CrossRef]
- Li, G.Q.; Zhou, J.Y.; Jia, H.Y.; Gao, Z.X.; Fan, M.; Luo, Y.J.; Zhao, P.T.; Xue, S.L.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Kong, L. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.X.; Liu, Y.G. Molecular control of male reproductive development and pollen fertility in rice. J. Integr. Plant Biol. 2012, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.; Condon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Aliyeva, A.J.; Aminov, N.K. Inheritance of the branching in hybrid populations among tetraploid wheat species and the new branched spike line 166-schakheli. Genet. Resour. Crop Evol. 2011, 58, 621–628. [Google Scholar] [CrossRef]
- Millet, E. Genetic control of heading date and spikelet number in common wheat (T. aestivum L.) line ‘Noa’. Theor. Appl. Genet. 1986, 72, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Y.; Lu, B.R.; Hu, X.R.; Yu, Y.; Zhang, Y. Inheritance of the triple-spikelet character in a Tibetan landrace of common wheat. Genet. Resour. Crop Evol. 2005, 52, 847–851. [Google Scholar] [CrossRef]
- Klindworth, D.L.; Williams, N.D.; Joppa, L.R. Chromosomal location of genes for supernumerary spikelet in tetraploid wheat. Genome 1990, 33, 515–520. [Google Scholar] [CrossRef]
- Zhang, R.Q.; Wang, X.E.; Chen, P.D. Inheritance and mapping of gene controlling four-rowed spike in tetraploid wheat (Triticum turgidum L.). Acta Agron. Sin. 2013, 39, 29–33. [Google Scholar] [CrossRef]
- Yang, W.Y. SSR mapping locus conferring on the triple-spikelet trait of the Tibetan triple-spikelet wheat (Triticum aestivum L. Concv. Tripletum). Triticeae Genom. Genet. 2011, 2, 1–6. [Google Scholar]
- Zhang, Z.; Belcramb, H.; Gornickic, P.; Charlesb, M.; Justb, J.; Huneaub, C. Duplication and partitioning in evolution and function of homoeologous Q loci governing domestication characters in polyploid wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 18737–18742. [Google Scholar] [CrossRef] [Green Version]
- Dobrovolskaya, O.; Martinek, P.; Voylokov, A.V.; Korzun, V.; Marion, S.R.; Andreas, B. Microsatellite mapping of genes that determine supernumerary spikelets in wheat (T. aestivum) and rye (S. cereale). Theor. Appl. Genet. 2009, 119, 867–874. [Google Scholar] [CrossRef]
- Sears, E.R. The Aneuploids of Common Wheat; Research Bulletin 572; University of Missouri: Columbia, MO, USA, 1954. [Google Scholar]
- Komatsu, M.; Chujo, A.; Nagato, Y.; Shimamoto, K.; Kyozuka, J. Frizzy panicle is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development 2003, 130, 3841–3850. [Google Scholar] [CrossRef] [Green Version]
- Chuck, G.; Muszynski, M.; Kellogg, E.; Hake, S.; Schmidt, R.J. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science 2002, 298, 1238–1241. [Google Scholar] [CrossRef]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N.; et al. Frizzy panicle drives supernumerary spikelets in bread wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, L.; Zhao, M.; Guo, L.; Guo, X.; Zhao, D.; Batool, A.; Dong, B.; Xu, H.; Cui, S.; et al. Wheat Frizzy Panicle activates VERNALIZATION1-A and HOMEOBOX4-A to regulate spike development in wheat. Plant Biotechnol. J. 2020. [Google Scholar] [CrossRef]
- Oliver, S.N.; Dennis, E.S.; Dolferus, R. ABA regulates apoplastic sugar transport and is a potential signal for cold-induced pollen sterility in rice. Plant Cell Physiol. 2007, 48, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Datta, R.; Chamusco, K.C.; Chourey, P.S. Starch biosynthesis during pollen maturation is associated with altered patterns of gene expression in maize. Plant Physiol. 2002, 130, 1645–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahir, A.; Kang, J.; Choulet, F.; Ravel, C.; Romeuf, I.; Rasouli, F.; Nosheen, A.; Branlard, G. Deciphering carbohydrate metabolism during wheat grain development via integrated transcriptome and proteome dynamics. Mol. Biol. Rep. 2020, 47, 5439–5449. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Xu, G.; Zhu, Q. Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling. Plant Physiol. 2004, 135, 1621–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | Entry | Number of BSL Plants | Number of NSL Plants | χ2 |
|---|---|---|---|---|
| 2018 | 1 | 72 | 68 | 0.064 |
| 2 | 74 | 75 | 0.000 | |
| 3 | 72 | 69 | 0.028 | |
| 4 | 77 | 76 | 0.000 | |
| 5 | 62 | 71 | 0.481 | |
| 6 | 73 | 53 | 2.865 | |
| 7 | 74 | 73 | 0.000 | |
| Total | 504 | 485 | 0.328 | |
| 2019 | 1 | 74 | 67 | 0.255 |
| 2 | 77 | 59 | 2.125 | |
| 3 | 78 | 77 | 0.000 | |
| 4 | 71 | 73 | 0.007 | |
| 5 | 51 | 55 | 0.085 | |
| 6 | 54 | 52 | 0.009 | |
| 7 | 69 | 67 | 0.007 | |
| Total | 474 | 450 | 0.573 |
| Days from Sowing to Heading | Days from Sowing to Anthesis | Grains Number per Spike | 1000-Grain Weight/g | |
|---|---|---|---|---|
| NSL | 173.4 ± 1.35 | 177.2 ± 2.02 | 44.0 ± 12.98 | 47.5 ± 14.91 |
| BSL | 175.6 ± 1.56 ** | 180.5 ± 1.72 ** | 55.9 ± 14.70 ** | 35.6 ± 5.74 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, T.; Li, L.; Zhao, Y.; Hua, C.; Sun, Z.; Li, T. A Heterozygous Genotype-Dependent Branched-Spike Wheat and the Potential Genetic Mechanism Revealed by Transcriptome Sequencing. Biology 2021, 10, 437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050437
Ma T, Li L, Zhao Y, Hua C, Sun Z, Li T. A Heterozygous Genotype-Dependent Branched-Spike Wheat and the Potential Genetic Mechanism Revealed by Transcriptome Sequencing. Biology. 2021; 10(5):437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050437
Chicago/Turabian StyleMa, Tian, Lei Li, Yang Zhao, Chen Hua, Zhengxi Sun, and Tao Li. 2021. "A Heterozygous Genotype-Dependent Branched-Spike Wheat and the Potential Genetic Mechanism Revealed by Transcriptome Sequencing" Biology 10, no. 5: 437. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10050437