
Expressional Profiling of TEX11, ESRα and BOLL Genes in Yak under Different Feeding Conditions

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Extraction of RNA and Synthesis of Complementary DNA
2.3. Primer Designing and PCR Amplification
2.4. Exploration through Quantitative Real-Time PCR
2.5. Exploration through Western Blotting
2.6. Immunochemistry of TEX11, ESRα and BOLL Proteins
2.7. Immunofluorescence Analysis
2.8. Statistical Analyses
3. Results
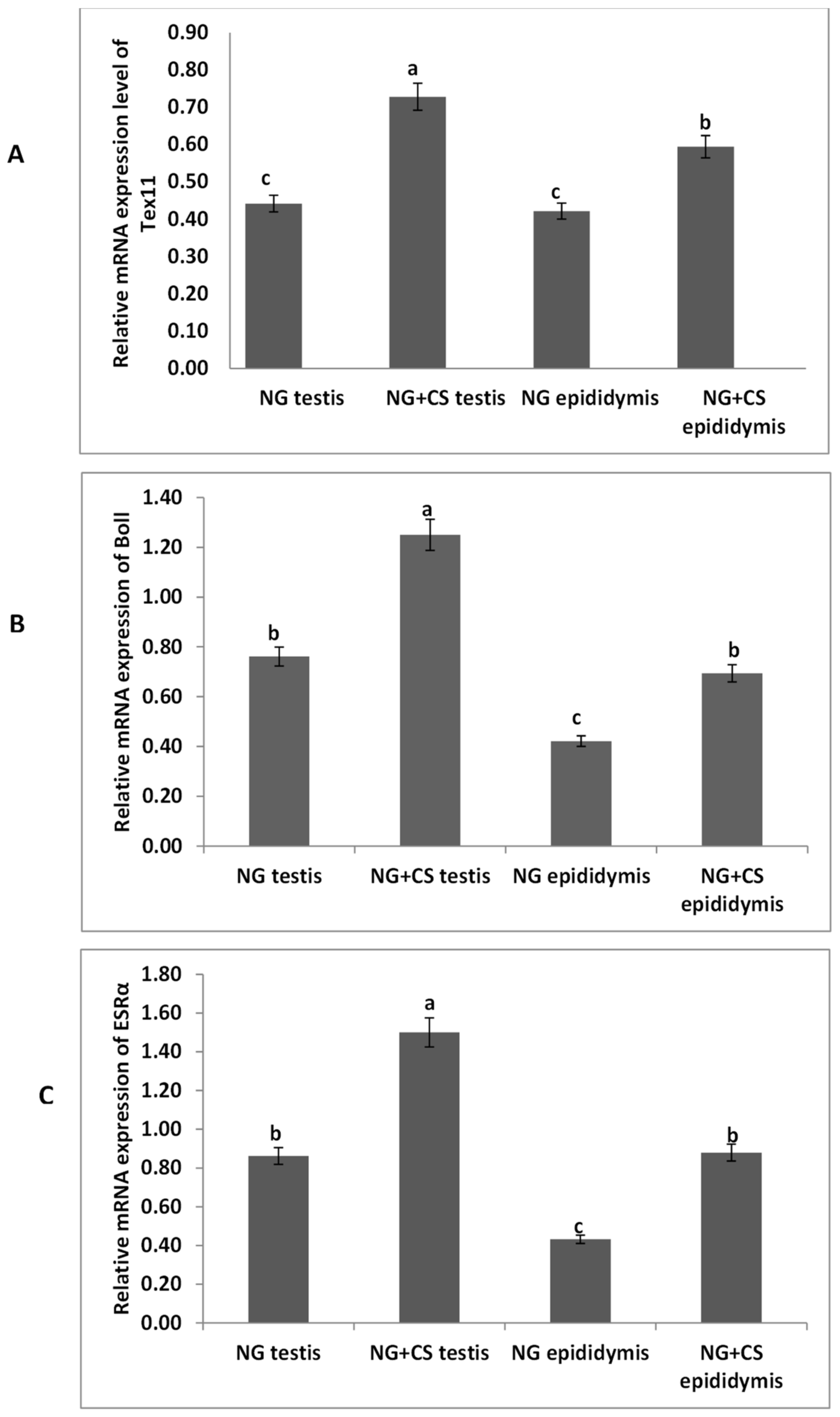
3.1. Expressional Profiling of TEX11, ESRα, and BOLL Genes through Quantitative Real-Time PCR
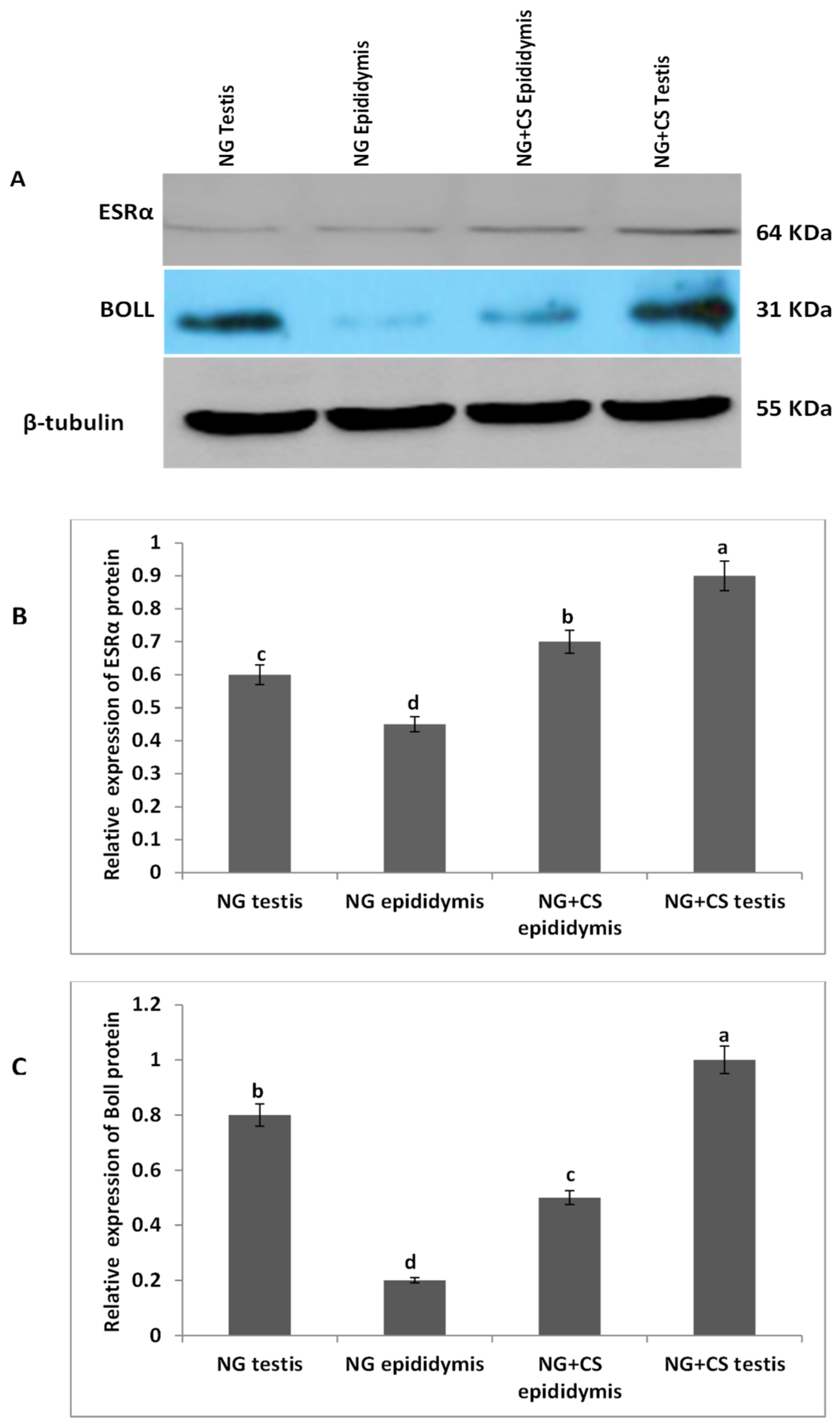
3.2. Western Blotting Analysis of ESRα and BOLL Proteins
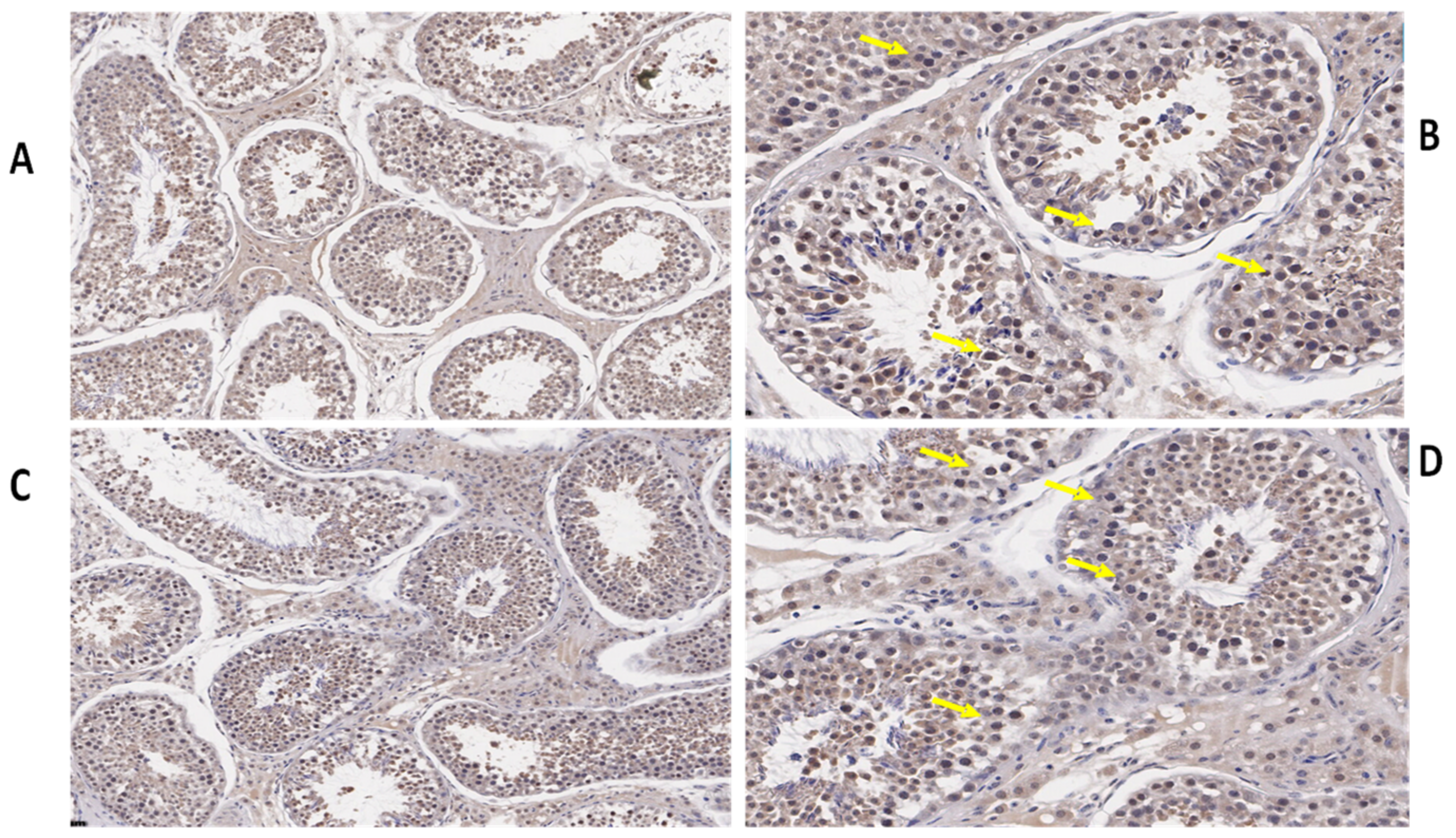
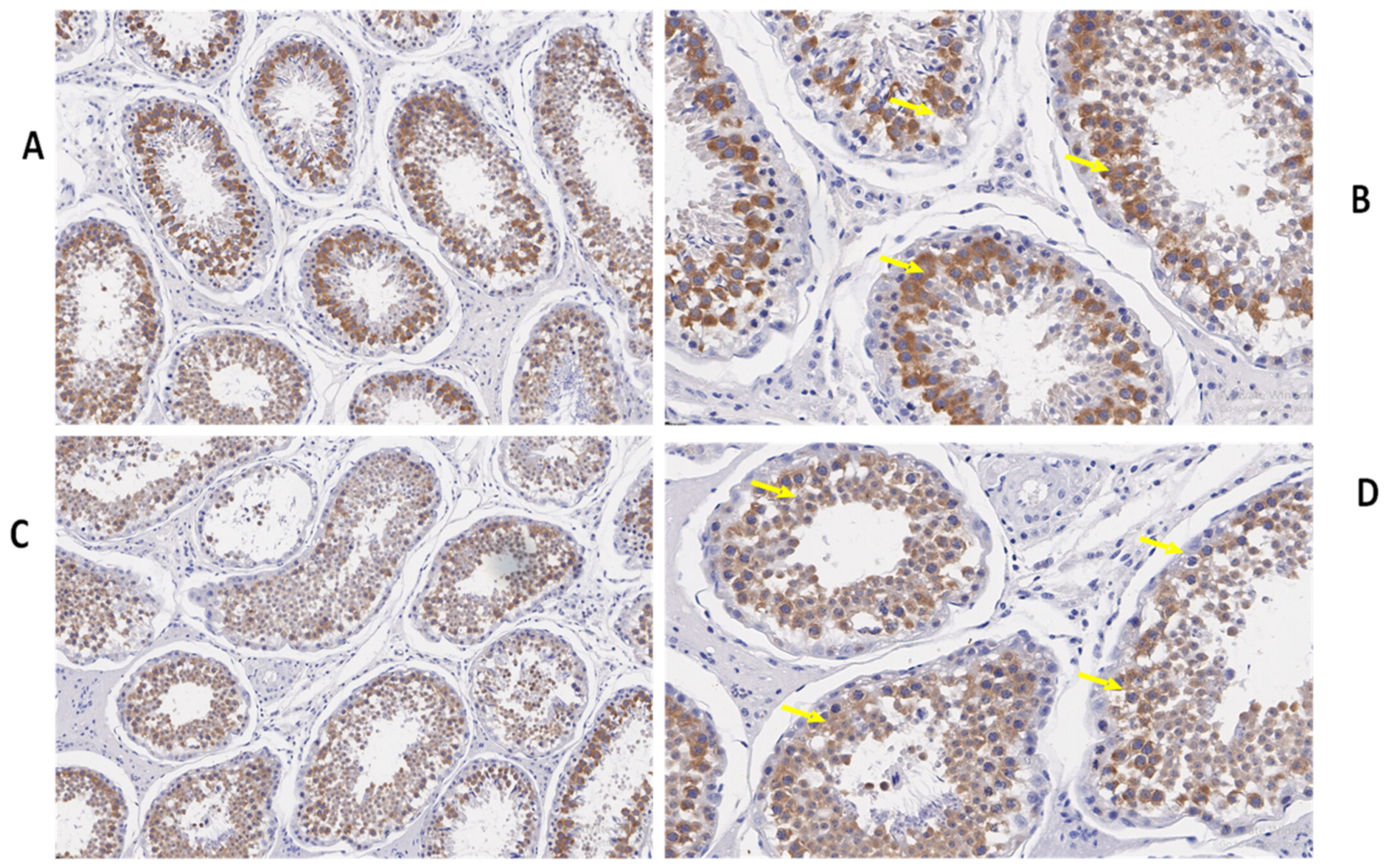
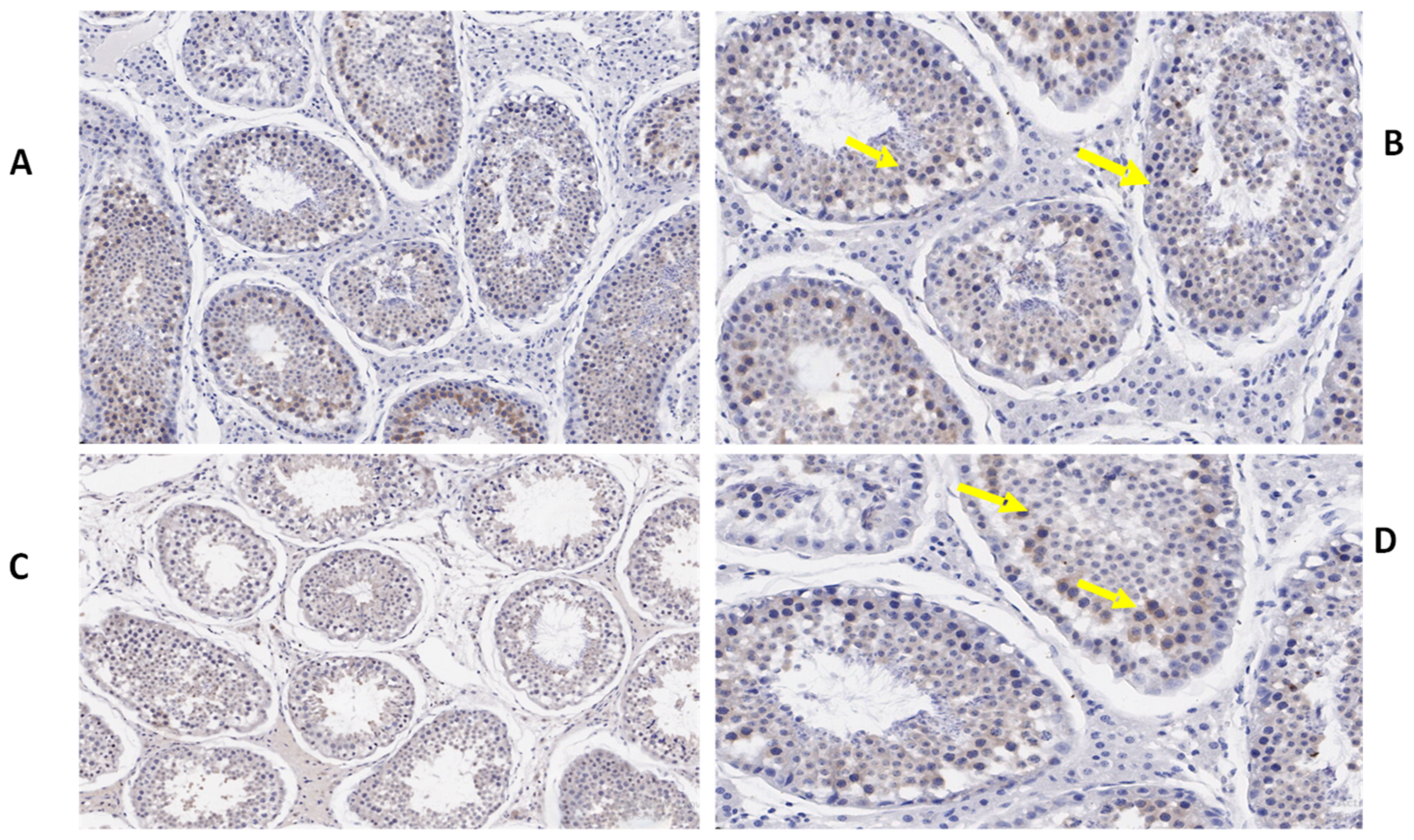
3.3. Immunohistochemistry of TEX11, ESRα, and BOLL Proteins
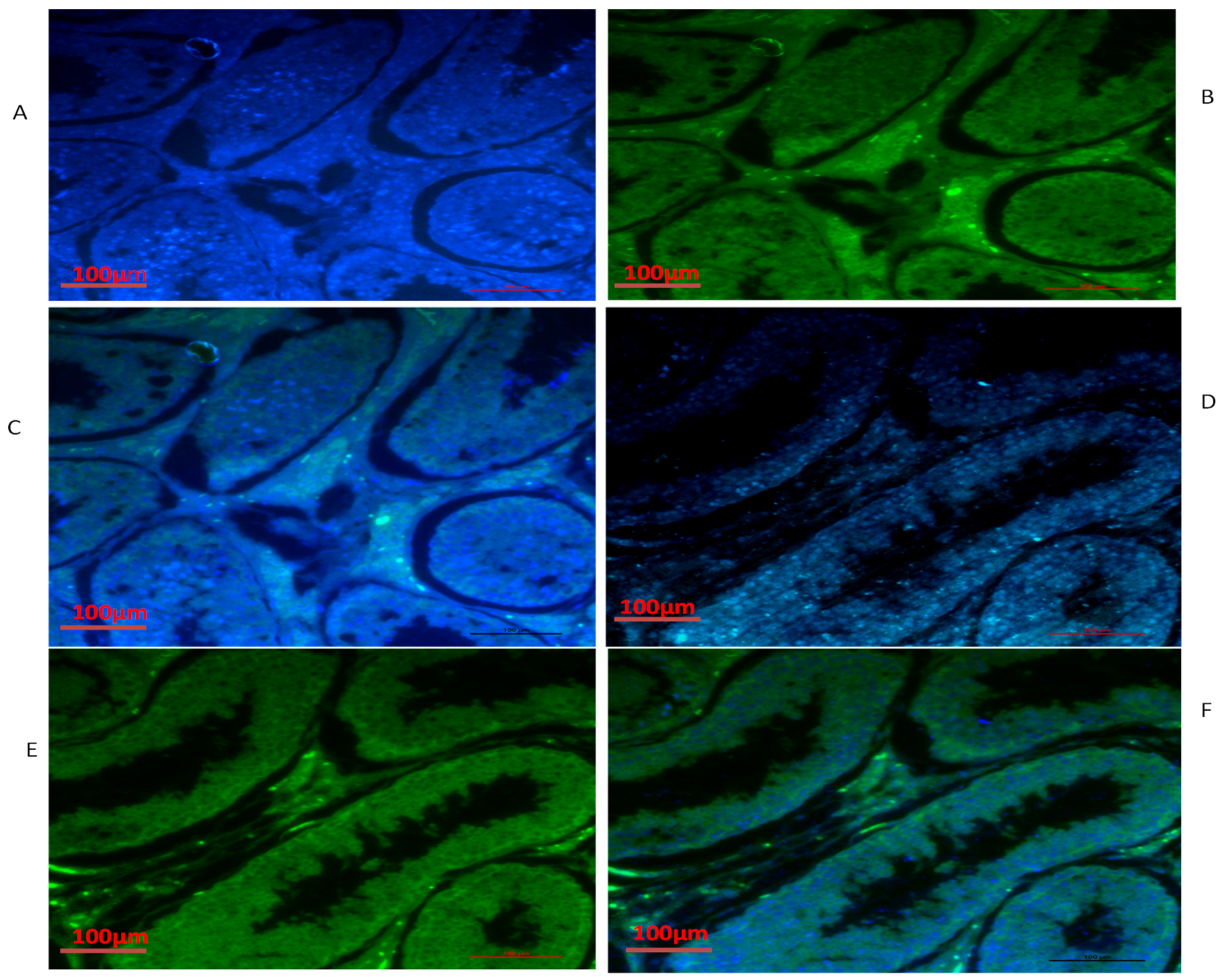
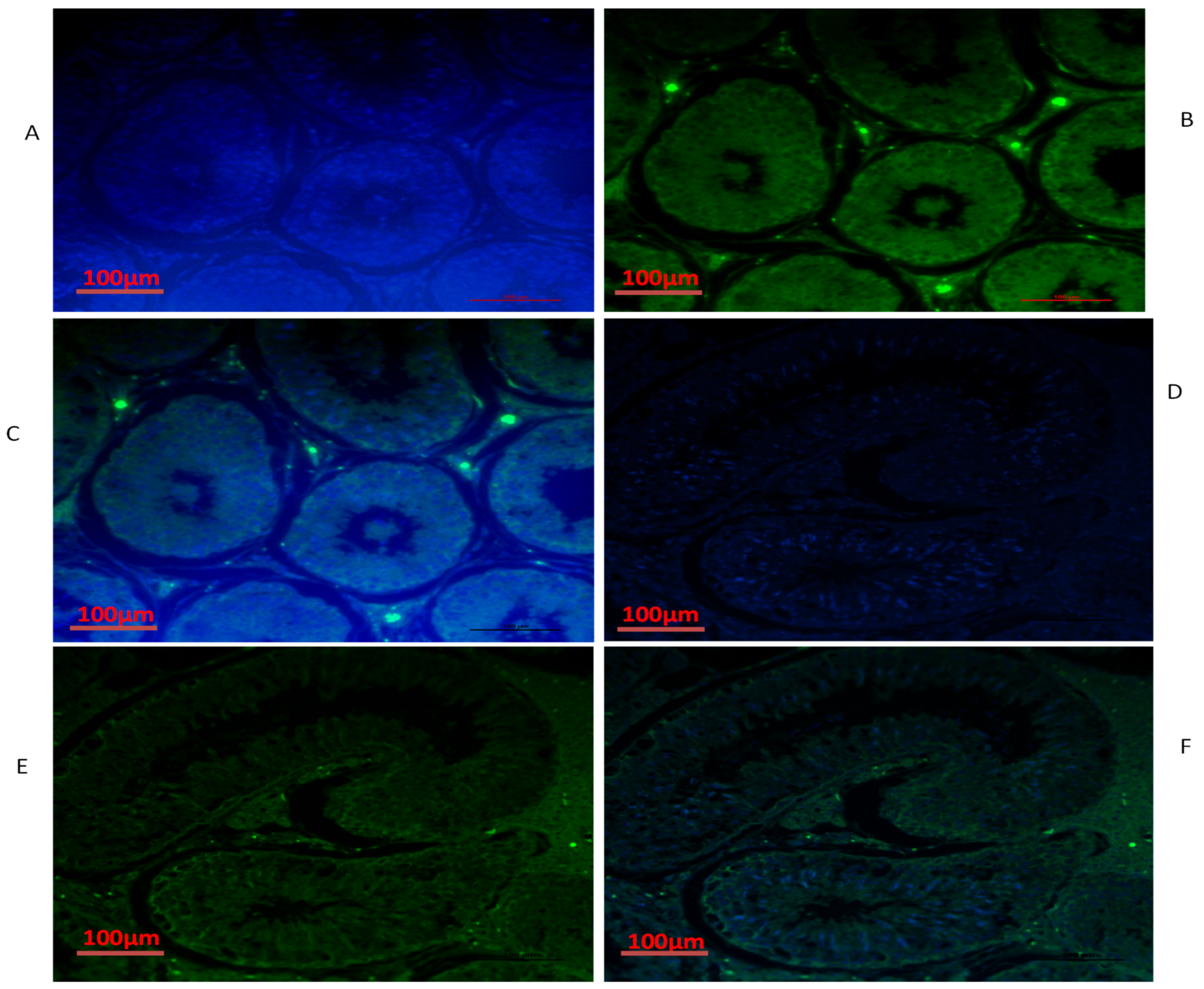
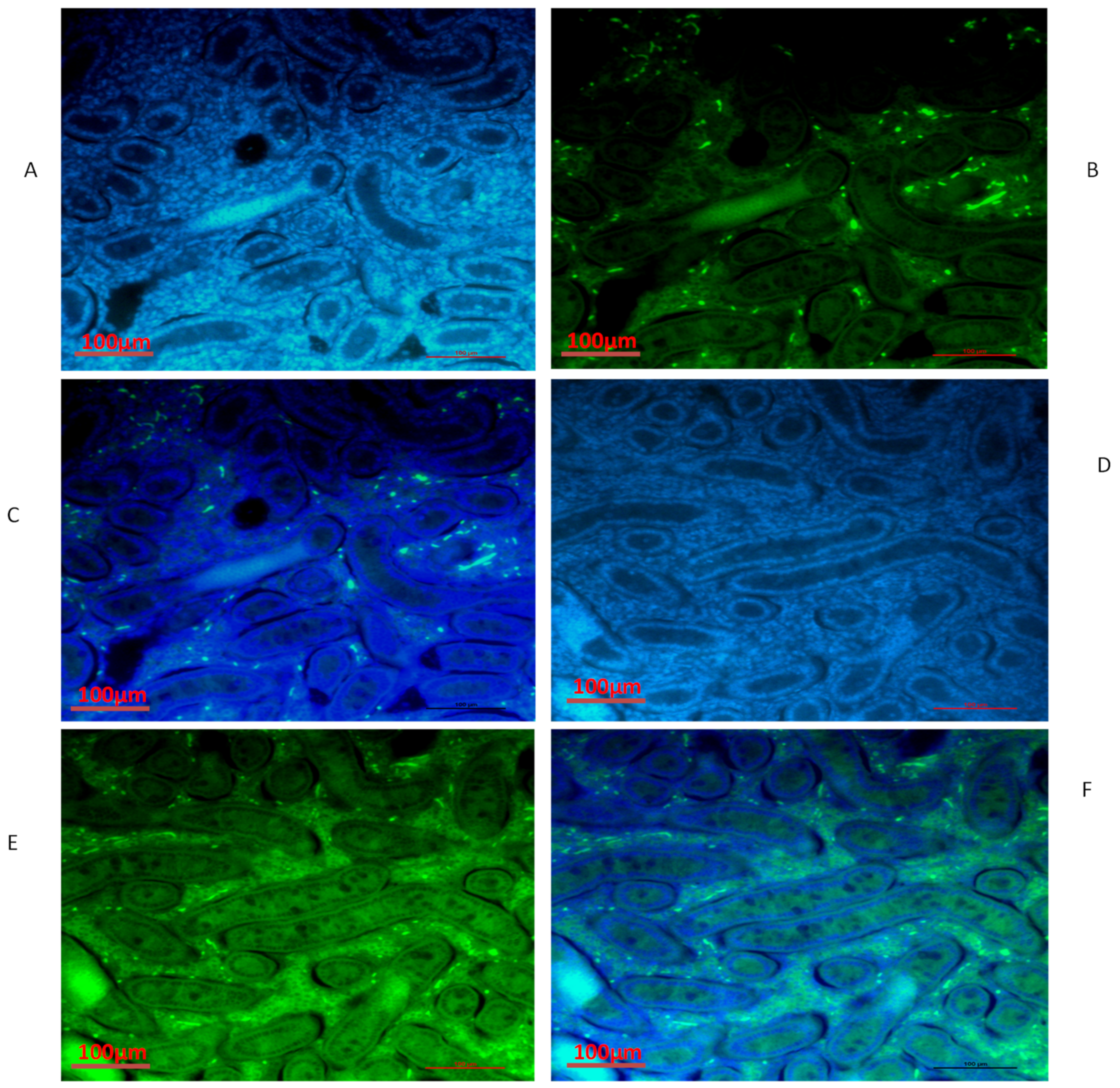
3.4. Immunofluorescence of TEX11, ESRα and BOLL Proteins
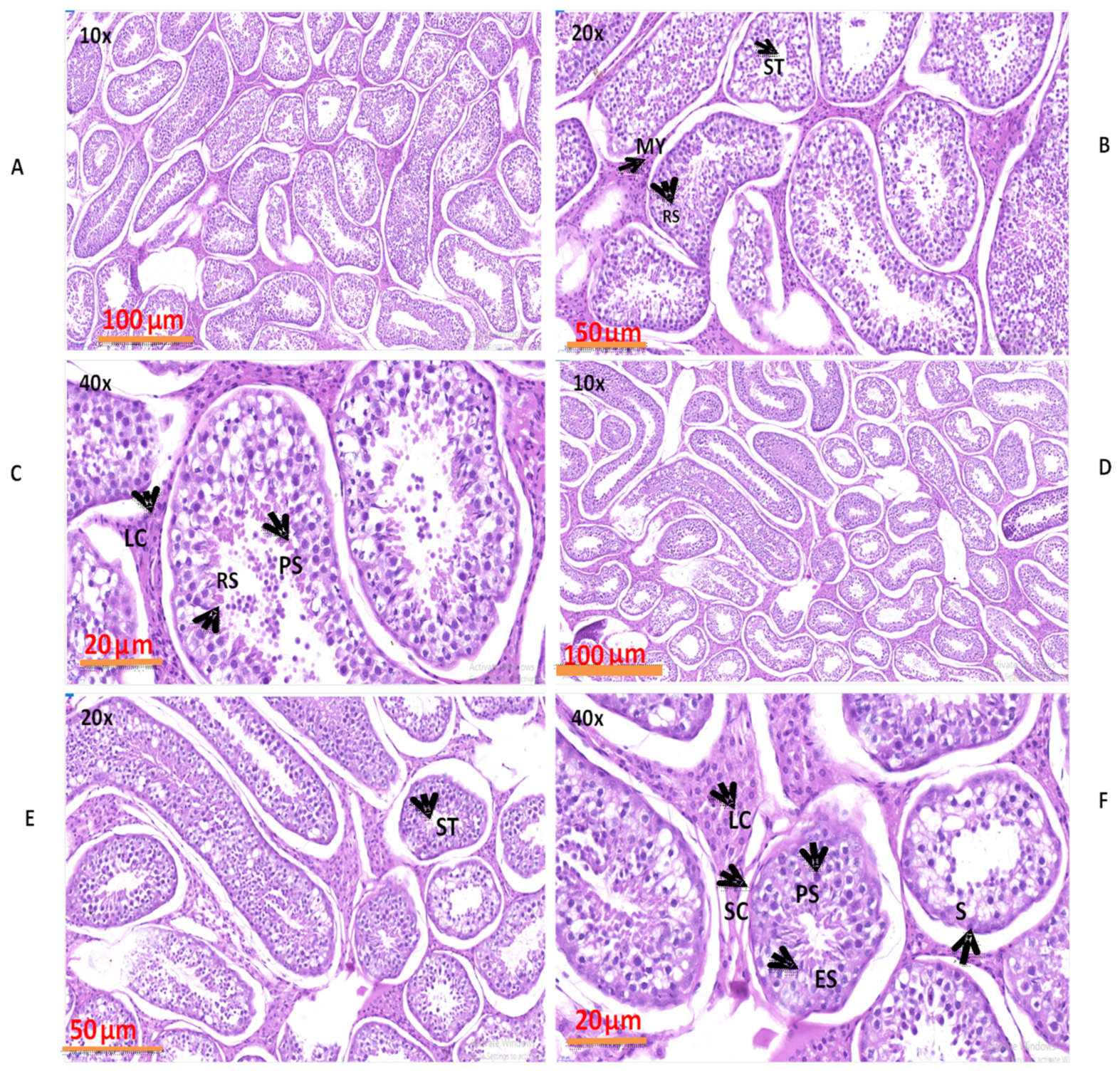
3.5. Evaluation of Numerous Morphological Variations in the Natural Grazing without Concentrate Supplementation and Natural Grazing Plus Concentrate Supplementation Groups
3.6. Spermatogenic Cells and Their Nuclei Diameters (μm) in Yak Testes
3.7. Diameters of Seminiferous Tubules and Numbers of Cells in the Testis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yugal, R.B.; Sulochana, S.; Nabaraj, S.; Tara, N.G. Effects of nutrition on reproduction—A review. Adv. Appl. Sci. Res. 2013, 4, 421–429. [Google Scholar]
- Boland, M.P.; Lonergan, P.; Callaghan, O. Effect of nutrition on endocrine parameters, physiology, and oocyte and embryo development. Theriogenology 2001, 55, 1323–1340. [Google Scholar] [CrossRef]
- Waddad, S.A.; Gaili, E.S. Effect of nutrition on sexual development of Western Sudan Baggara bull calves. Acta Vet. Beogr. 1985, 35, 299–304. [Google Scholar]
- Hansen, M.; Flatt, T.; Aguilaniu, H. Reproduction, Fat Metabolism, and Lifespan—What Is the Connection. Cell Metab. 2013, 8, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Corona, G.; Mannucci, E.; Forti, G.; Maggi, M. Hypogonadism, ED, metabolic syndrome and obesity: A pathological link supporting cardiovascular diseases. Int. J. Androl. 2009, 32, 587–598. [Google Scholar] [CrossRef]
- Judd, E.T.; Wessels, F.J.; Drewry, M.D.; Grove, M.; Wright, K.; Hahn, D.A.; Hatle, J.D. Ovariectomy in grasshoppers increases somatic storage, but proportional allocation of ingested nutrients to somatic tissues is unchanged. Aging Cell 2011, 10, 972–979. [Google Scholar] [CrossRef] [Green Version]
- McNamara, J.P.; Shields, S.L. Reproduction during lactation of dairy cattle: Integrating nutritional aspects of reproductive control in a systems research approach. Anim. Front. 2013, 4, 76–83. [Google Scholar] [CrossRef]
- Wang, P.J.; McCarrey, J.R.; Yang, F.; Page, D.C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 2001, 27, 422–426. [Google Scholar] [CrossRef]
- Wang, P.J.; Page, D.C.; McCarrey, J.R. Differential expression of sex-linked and autosomal germ-cell-specific genes during spermatogenesis in the mouse. Hum. Mol. Genet. 2005, 14, 2911–2918. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Gell, K.; Van Der Heijden, G.W.; Eckardt, S.; Leu, N.A.; Page, D.C.; Benavente, R.; Her, C.; Höög, C.; McLaughlin, K.J.; et al. Meiotic failure in male mice lacking an X-linked factor. Genes Dev. 2008, 22, 682–691. [Google Scholar] [CrossRef] [Green Version]
- Adelman, C.A.; Petrini, J.H. ZIP4H (TEX11) deficiency in the mouse impairs meiotic double strand break repair and the regulation of crossing over. PLoS Genet. 2008, 4, 1000042. [Google Scholar] [CrossRef] [Green Version]
- Cooke, P.S.; Nanjappa, M.K.; Ko, C.; Prins, G.S.; Hess, R.A. Estrogens in male physiology. Physiol. Rev. 2017, 97, 995–1043. [Google Scholar] [CrossRef]
- Hess, R.A. Estrogen in the adult male reproductive tract: A review. Reprod. Biol. Endocrinol. 2003, 1, 52. [Google Scholar] [CrossRef] [Green Version]
- Delbes, G.; Levacher, C.; Habert, R. Estrogen effects on fetal and neonatal testicular development. Reproduction 2006, 132, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Lucas, T.F.G.; Siu, E.; Esteves, C.A.; Monteiro, H.P.; Oliveira, C.; Porto, C.S.; Lazari, M.F.M. 17beta-estradiol induces the translocation of the estrogen receptors ESR1 and ESR2 to the cell membrane, MAPK3/1 phosphorylation and proliferation of cultured immature rat Sertoli cells. Biol. Reprod. 2008, 78, 101–114. [Google Scholar] [CrossRef] [Green Version]
- McKinnell, C.; Atanassova, N.; Williams, K.; Fisher, J.S.; Walker, M.; Turner, K.J.; Saunders, T.K.; Sharpe, R.M. Suppression of androgen action and the induction of gross abnormalities of the reproductive tract in male rats treated neonatally with diethylstilbestrol. J. Androl. 2001, 22, 323–338. [Google Scholar]
- Abney, T.O.; Myers, R.B. 17 beta-estradiol inhibition of Leydig cell regeneration in the ethane dimethylsulfonate-treated mature rat. J. Androl. 1991, 12, 295–304. [Google Scholar] [PubMed]
- Li, T.; Wang, X.; Zhang, H.; Chen, Z.; Zhao, X.; Ma, Y. Histomorphological Comparisons and Expression Patterns of BOLL Gene in Sheep Testes at Different Development Stages. Animals 2019, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Ahmadivand, S.; Farahmand, H.; Teimoori-Toolabi, L.; Mirvaghefi, A.; Eagderi, S.; Geerinckx, T.; Shokrpoor, S.; Rahmati-Holasoo, H. Boule gene expression underpins the meiotic arrest in spermatogenesis in male rainbow trout (Oncorhynchus mykiss) exposed to DEHP and butachlor. Gen. Comp. Endocrinol. 2016, 225, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xue, P.; Gao, L.; Chen, X.; Lin, K.; Yang, X.; Dai, Y.; Xu, E.Y. Highly conserved epigenetic regulation of BOULE and DAZL is associated with human fertility. FASEB J. 2016, 30, 3424–3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luetjens, C.M.; Xu, E.Y.; Reijo Pera, R.A.; Kamischke, A.; Nieschlag, E.; Gromoll, J. Association of meiotic arrest with lack of BOULE protein expression in fertile men. J. Clin. Endocrinol. Metab. 2004, 4, 1926–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, E.Y.; Moore, F.L.; Reijo, R.P. A gene family required for human germ cell development evolved from an ancient meiotic gene conserved in metazoans. Proc. Natl. Acad. Sci. USA 2001, 98, 7414–7419. [Google Scholar] [CrossRef] [Green Version]
- Qiang, Q.; Guojie, Z.; Tao, M.; Wubin, Q.; Junyi, W.; Zhiqiang, Y.; Changchang, C.; Quanjun, H.; Jaebum, K.; Larkin, D.M. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 946–949. [Google Scholar]
- Miao, F.; Guo, Z.; Xue, R.; Wang, X.; Shen, Y. Effects of grazing and precipitation on herbage biomass, herbage nutritive value, and yak performance in an alpine meadow on the qinghai-tibetan plateau. PLoS ONE 2015, 10, e0127275. [Google Scholar] [CrossRef]
- Guo, X.; Long, R.; Kreuzer, M.; Ding, L.; Shang, Z.; Zhang, Y.; Yang, Y.; Cui, G. Importance of Functional ingredients in yak milk-derived food on health of Tibetan nomads living under high-altitude stress: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 292–302. [Google Scholar] [CrossRef]
- Nichi, M.; Bols, P.E.; Zuge, R.M.; Barnabe, V.H.; Goovaerts, I.G.; Barnabe, R.C.; Cortada, C.N. Seasonal variation in semen quality in Bos indicus and Bos taurus bulls raised under tropical conditions. Theriogenology 2006, 66, 822–828. [Google Scholar] [CrossRef]
- Kowalowka, M.; Wysocki, P.; Fraser, L.; Strzezek, J. Extracellular superoxide dismutase of boar seminal plasma. Reprod. Domest. Anim. 2008, 43, 490–496. [Google Scholar] [CrossRef]
- Casao, A.; Cebrian, I.; Asumpcao, M.E.; Perez-Pe, R.; Abecia, J.A.; Forcada, F.; Cebrian-Perez, J.A.; Muino-Blanco, T. Seasonal variations of melatonin in ram seminal plasma are correlated to those of testosterone and antioxidant enzymes. Reprod. Biol. Endocrinol. 2010, 8, 59. [Google Scholar] [CrossRef] [Green Version]
- Marti, E.; Mara, L.; Marti, J.; Muiño-Blanco, T.; Pérez, J.; Álvaro, C. Seasonal variations in antioxidant enzyme activity in ram seminal plasma. Theriogenology 2007, 67, 1446–1454. [Google Scholar] [CrossRef]
- Long, R.; Ding, L.; Shang, Z.; Guo, X. The yak grazing system on the Qinghai-Tibetan plateau and its status. Rangel. J. 2008, 30, 241–246. [Google Scholar]
- Rhynes, W.E.; Ewing, L.L. Testicular endocrine function in Hereford bulls exposed to high ambient temperature. Endocrinology 1973, 92, 509–515. [Google Scholar] [CrossRef]
- Minton, J.E.; Wettemann, R.P.; Meyerhoeffer, D.C.; Hintz, R.L.; Turman, E.J. Serum luteinizing hormone and testosterone in bulls during exposure to elevated ambient temperature. J. Anim. Sci. 1981, 53, 1551–1558. [Google Scholar] [CrossRef]
- De Amicis, F.; Santoro, M.; Gervasi, S.; Aquila, S. Nutrition and Male Reproduction: Nutrients Directly Affect Sperm Performance. Int. J. Nutr. Sci. 2018, 3, 1025. [Google Scholar]
- Qudratullah, K.; Xuezhi, D.; Anum, A.A.; Min, C.; Xiaoyun, W.; Pengjia, B.; Ping, Y. Expression Analysis of IZUMO1 Gene during Testicular Development of Datong Yak (Bos Grunniens). Animals 2019, 9, 292. [Google Scholar]
- Hou, Y.; Zhou, X.; Liu, J.; Yuan, J.; Cheng, H.; Zhou, R. Nuclear factor-Y (NF-Y) regulates transcription of mouse Dmrt7 gene by binding to tandem CCAAT boxes in its proximal promoter. Int. J. Biol. Sci. 2010, 6, 655. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO: Geneva, Switzerland, 2010. [Google Scholar]
- Jungwirth, A.; Giwercman, A.; Tournaye, H.; Diemer, T.; Kopa, Z.; Dohle, G.; Krausz, C.G. European Association of Urology guidelines on Male Infertility: The 2012 update. Eur. Urol. 2012, 62, 324. [Google Scholar] [CrossRef]
- Abbasi, A.; Prasad, A.; Rabbani, P. Experimental zinc deficiency in man: Effect on spermatogenesis. Trans. Assoc. Am. Physicians 1979, 92, 292–302. [Google Scholar]
- Hoedemaker, M.; Prange, D.; Gundelach, Y. Body condition change ante- and postpartum, health and reproductive performance in German Holstein cows. Reprod. Domest. Anim. 2009, 44, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Smorag, L.; Xu, X.; Engel, W.; Pantakani, D. The roles of DAZL in RNA biology and development. Wiley Interdiscip. Rev. RNA 2014, 5, 527–535. [Google Scholar] [CrossRef]
- Fu, X.F.; Cheng, S.F.; Wang, L.Q.; Yin, S.; Felici, M.D.; Shen, W. DAZ family proteins, key players for germ cell development. Int. J. Biol. Sci. 2015, 11, 1226–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Liu, C.; Zhu, H.; Sun, J.; Yu, M.; Niu, Z.; Liu, W.; Peng, S.; Hua, J. Expression pattern of Boule in dairy goat testis and its function in promoting the meiosis in male germline stem cells (mGSCs). J. Cell. Biochem. 2013, 114, 294–302. [Google Scholar] [CrossRef]
- Shah, C.; Vangompel, M.J.; Naeem, V.; Chen, Y.; Lee, T.; Angeloni, N.; Wang, Y.; Xu, E.Y. Widespread presence of human BOULE homologs among animals and conservation of their ancient reproductive function. PLoS Genet. 2010, 6, e1001022. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, C.R.; Dorfman, V.B.; Vitullo, A.D. IGF1 regulation of BOULE and CDC25A transcripts via a testosterone-independent pathway in spermatogenesis of adult mice. Reprod. Biol. 2015, 15, 48–55. [Google Scholar] [CrossRef]
- Kim, B.; Rhee, K. BOULE, a deleted in azoospermia homolog, is recruited to stress granules in the mouse male germ cells. PLoS ONE 2016, 11, e0163015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.Z.; Yan, G.Y.; Han, L.; Pang, J.; Zhong, B.S.; Zhang, G.M.; Wang, F.; Zhang, Y.L. Overexpression of STRA8, BOULE, and DAZL genes promotes goat bone marrow-derived mesenchymal stem cells in vitro transdifferentiation toward putative male germ cells. Reprod. Sci. 2017, 24, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Dwarakanath, M.; Lim, M.; Xu, H.; Hong, Y. Differential expression of boule and dazl in adult germ cells of the Asian seabass. Gene 2014, 549, 237–242. [Google Scholar] [CrossRef]
- Xu, H.; Li, Z.; Li, M.; Wang, L.; Hong, Y. Boule is present in fish and bisexually expressed in adult and embryonic germ cells of medaka. PLoS ONE 2009, 4, e6097. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.; Mäkelä, S.; Treuter, E.; Tujague, M.; Thomsen, J.; Andersson, G.; Enmark, E.; Pettersson, K.; Warner, M.; Gustafsson, J.Å. Mechanisms of estrogen action. Physiol. Rev. 2001, 81, 1535–1565. [Google Scholar] [CrossRef] [PubMed]
- Hess, R.A.; Carnes, K. The role of estrogen in testis and the male reproductive tract: A review and species comparison. Anim. Reprod. 2004, 1, 5–30. [Google Scholar]
- Carreau, S.; de Vienne, C.; Galeraud-Denis, I. Aromatase and estrogens in man reproduction: A review and latest advances. Adv. Med. Sci. 2008, 53, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, G.; El-Alfy, M. Immunocytochemical localization of estrogen receptors alpha and beta in the human reproductive organs. J. Clin. Endocrinol. Metab. 2000, 85, 4835–4840. [Google Scholar] [PubMed] [Green Version]
- Carpino, A.; Rago, V.; Pezzi, V.; Carani, C.; Andò, S. Detection of aromatase and estrogen receptors (ERalpha, ERbeta1, ERbeta2) in human Leydig cell tumor. Eur. J. Endocrinol. 2007, 157, 239–244. [Google Scholar] [CrossRef]
- Saunders, P.T.; Sharpe, R.M.; Williams, K.; MacPherson, S.; Urquart, H.; Irvine, D.; Millar, M.R. Differential expression of oestrogen receptor alpha and beta proteins in the testes and male reproductive system of human and non-human primates. Mol. Hum. Reprod. 2001, 7, 227–236. [Google Scholar] [CrossRef]
- O’Donnell, L.; Robertson, K.M.; Jones, M.E.; Simpson, E.R. Estrogen and spermatogenesis. Endocr. Rev. 2001, 22, 289–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.H.; Makela, S.; Andersson, L.C.; Salmi, S.; Saji, S.; Webster, J.I.; Jensen, E.V.; Nilsson, S.; Warner, M.; Gus-Tafsson, J.A. A role for estrogen receptor as in the regulation of growth of the ventral prostate. Proc. Natl. Acad. Sci. USA 2001, 98, 6330–6335. [Google Scholar]
- Tang, L.; Zeng, W.; Clark, R.K.; Dobrinski, I. Characterization of the porcine testis-expressed gene 11 (Tex11). Spermatogenesis 2011, 1, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Tamowski, S.; Aston, K.I.; Carrell, D.T. The use of transgenic mouse models in the study of male infertility. Syst. Biol. Reprod. Med. 2010, 56, 260–273. [Google Scholar] [CrossRef]
- Cooke, H.J.; Saunders, P.T. Mouse models of male infertility. Nat. Rev. 2002, 3, 790–801. [Google Scholar] [CrossRef]
- Ahmad, N.; Umair, S.; Shahab, M.; Arslan, M. Testicular development and establishment of spermatogenesis in Nili-Ravi buffalo bulls. Theriogenology 2010, 73, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Akosman, M.S.; Lenger, O.F.; Demirel, H.H. Morphological, stereological and histometrical assessment of the testicular parameters between Holstein and Simmental bulls. Int. J. Morphol. 2013, 31, 1076–1080. [Google Scholar] [CrossRef]
- Paulo, A.T.A.; Deiler, S.C.; Fábio, J.C.F.; Carlos, A.C.F.; Marcelo, D.S.; Juliana, C.B.S. Testicular Histomorphometric Evaluation of Zebu Bull Breeds. Braz. Arch. Biol. Technol. 2014, 57, 900–907. [Google Scholar]
- Franca, L.R.; Russell, L.D. The testis of domestic animals in: Male reproduction. In A Multidisciplinary Overview; Regadera, J., Martinez-Garcia, F., Eds.; Churchill Livingstone: Madrid, Spain, 1998; pp. 197–219. [Google Scholar]
- Russell, L.; Ettlin, R.; Sinha Hikim, A.; Clegg, E. Histological and Histopathological Evaluation of the Testis; Cache River Press: Clearwater, FL, USA, 1990; ISBN 0-9627422-0-1. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Grazing Group (Forage) | Concentrated Group |
|---|---|---|
| Protein | 8.86 ± 0.58 | 28.39 ± 0.86 |
| Calcium | 3.73 ± 0.07 | 4.80 ± 0.05 |
| Phosphorus | 0.14 ± 0.03 | 0.17 ± 0.03 |
| Neutral detergent fiber | 60.45 ± 0.91 | 52.52 ± 0.09 |
| Fat | 0.89 ± 0.89 | 4.34 ± 0.15 |
| Ash | 8.97 ± 0.57 | 8.75 ± 0.07 |
| Accession No. | Gene | Primers Sequence (5′ → 3′) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| AY656813.1 | ESRα | F: GCCATAGCCCATGCCTTTT R: TCATGGGGCTGTATGTGGTTT | 748 | 60.33 59.64 |
| NM_001168704.1 | BOLL | F: AATAACCCAACGAGTGC R: AGTGACGAAGCCATACC | 147 | 59.97 59.67 |
| XM_024988612.1 | TEX11 | F: TCACGTCGGAGGAGCTTGAT R: AATTGCCGCAGGTGTGGTAG | 145 | 55.00 55.00 |
| NM_001034034.2: | GAPDH | F: AATGAAAGGGCCATCACCATC R: CACCACCCTGTTGCTGTAGCCA | 204 | 55.85 60.00 |
| Germ Cells | NG | NG + CS |
|---|---|---|
| Spermatogonium | 5.10 ± 0.60 a | 6.20 ± 1.00 b |
| Spermatogonium nuclei | 3.20 ± 1.10 a | 4.50 ± 1.20 b |
| Spermatogonium nuclei | 5.60 ± 0.70 a | 6.20 ± 0.90 b |
| Primary spermatocyte | 3.00 ± 1.00 a | 4.60 ± 1.18 b |
| Primary spermatocyte nuclei | 6.50 ± 1.69 a | 6.90 ± 1.30 a |
| Round spermatid | 6.89 ± 1.72 a | 7.01 ± 1.21 a |
| Sertoli cells | 4.50 ± 0.86 a | 5.10 ± 0.69 b |
| Leydig cells | 6.00 ± 1.19 a | 7.10 ± 1.55 b |
| Germ Cells | NG | NG + CS |
|---|---|---|
| Tubular diameter (μm) | 225.30 ± 0.70 a | 250.31 ± 0.44 b |
| Epithelial height (μm) | 65.70 ± 0.43 a | 73.79 ± 0.60 b |
| Luminal diameter (µm2) | 74.90 ± 0.40 a | 85.62 ± 0.90 b |
| Luminal area (µm2) | 60.56 ± 4.60 a | 70.10 ± 6.38 b |
| Leydig cell area (µm2) | 134.43 ± 20.23 a | 140.76 ± 27.30 b |
| Width of tunica | 13.90 ± 1.00 a | 14.60 ± 0.50 a |
| albuginea (µm) | ||
| ST volume density (%) | 72.10 ± 0.70 a | 79.84 ± 0.40 b |
| Leydig cells (%) | 27.20 ± 1.90 a | 29.50 ± 2.40 a |
| Sertoli cells (%) | 30.40 ± 1.63 a | 38.10 ± 2.70 b |
| Spermatogonium (%) | 175 ± 15.80 a | 190 ± 15.55 b |
| Spermatocyte (%) | 16.60 ± 1.40 a | 21.00 ± 1.60 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalwar, Q.; Chu, M.; Ahmad, A.A.; Xiong, L.; Zhang, Y.; Ding, X.; Yan, P. Expressional Profiling of TEX11, ESRα and BOLL Genes in Yak under Different Feeding Conditions. Biology 2021, 10, 731. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10080731
Kalwar Q, Chu M, Ahmad AA, Xiong L, Zhang Y, Ding X, Yan P. Expressional Profiling of TEX11, ESRα and BOLL Genes in Yak under Different Feeding Conditions. Biology. 2021; 10(8):731. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10080731
Chicago/Turabian StyleKalwar, Qudratullah, Min Chu, Anum Ali Ahmad, Lin Xiong, Yongfeng Zhang, Xuezhi Ding, and Ping Yan. 2021. "Expressional Profiling of TEX11, ESRα and BOLL Genes in Yak under Different Feeding Conditions" Biology 10, no. 8: 731. https://0-doi-org.brum.beds.ac.uk/10.3390/biology10080731