
Amanita Section Phalloideae Species in the Mediterranean Basin: Destroying Angels Reviewed

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbarium Collections and Morphological Studies
2.2. Phylogenetic Studies
2.3. Toxicological Studies
2.4. Epidemiological Studies
- Ministry of Science and Innovation–National Center of Epidemiology (CNE)–Sistema de Vigilancia de la Red Nacional de Vigilancia Epidemiológica (RENAVE), SiViEs platform: for the period 1980–2020, outbreaks in which the agent was a fungal toxin and those associated with another agent in which mushrooms were consumed. In this database, only clusters (n ≥ 2 patients) are registered. To target putative phalloidian cases, the criterion “Incubation time” was selected for values > 6 h (when recorded) coupled with “hospitalization”, assuming that phalloidian poisonings required hospitalizations. Cases involving non-phalloidian fungal species were discarded.
- Ministry of Health, Consumer Affairs and Social Welfare-Specialized Health Care Activity Register (CMBD and RAE-CMB data). These databases record individual hospital exits and summarize care pathways (diagnostics, hospitalization times, etc.). For the period 1997–2015 (MBDS-H; ICD 9), clinical criteria 988.1 (toxic effect of mushrooms and mushrooms) and 573.1 (hepatitis unspecified) were selected. For the period 2016–2020 (RAE-CMD; IC10), clinical criteria T62.0X1A (toxic effect of ingested mushrooms, unintentional/accidental) and K71.2 (toxic hepatopathy with acute hepatitis) were retained.
3. Results
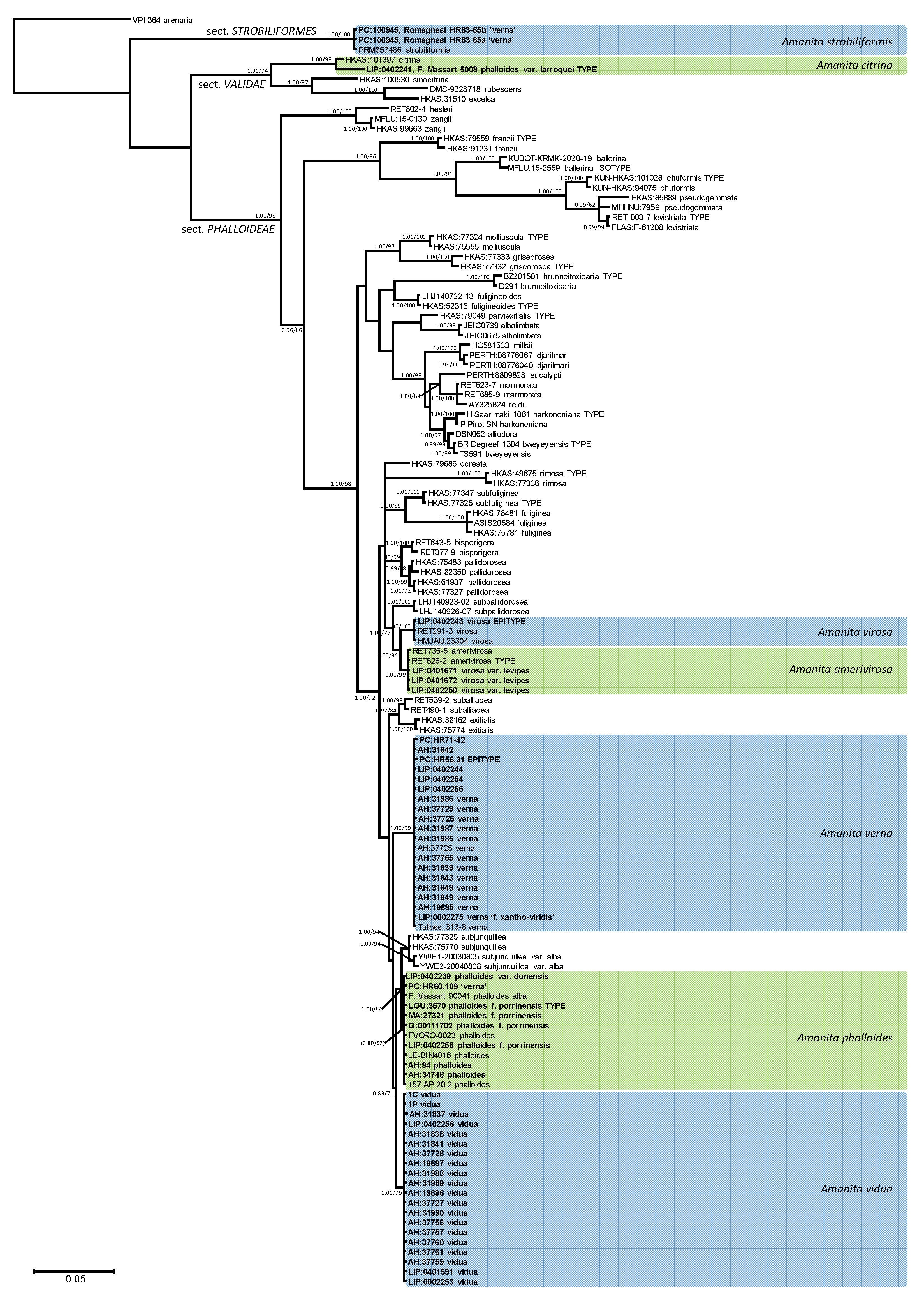
3.1. Phylogeny
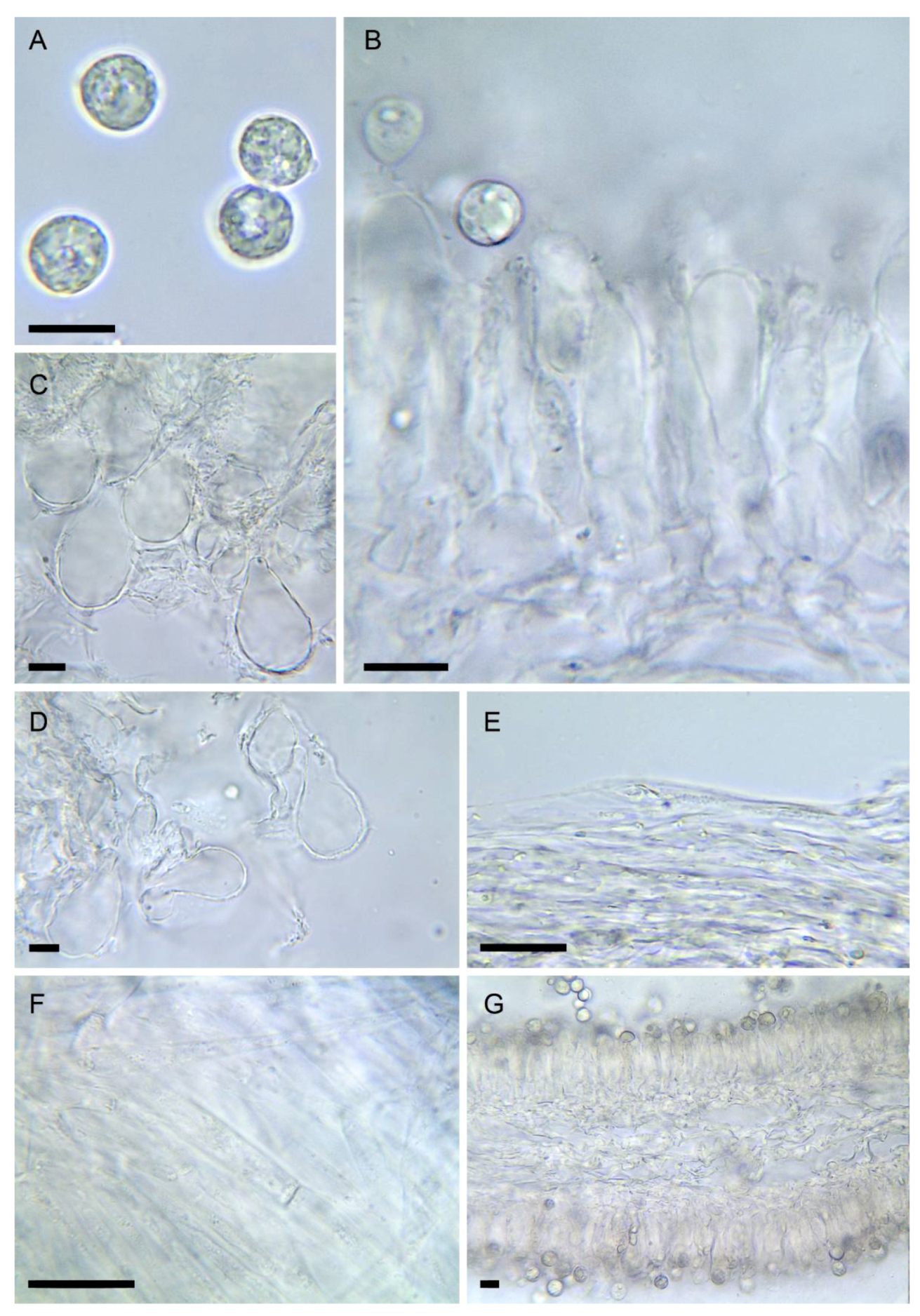
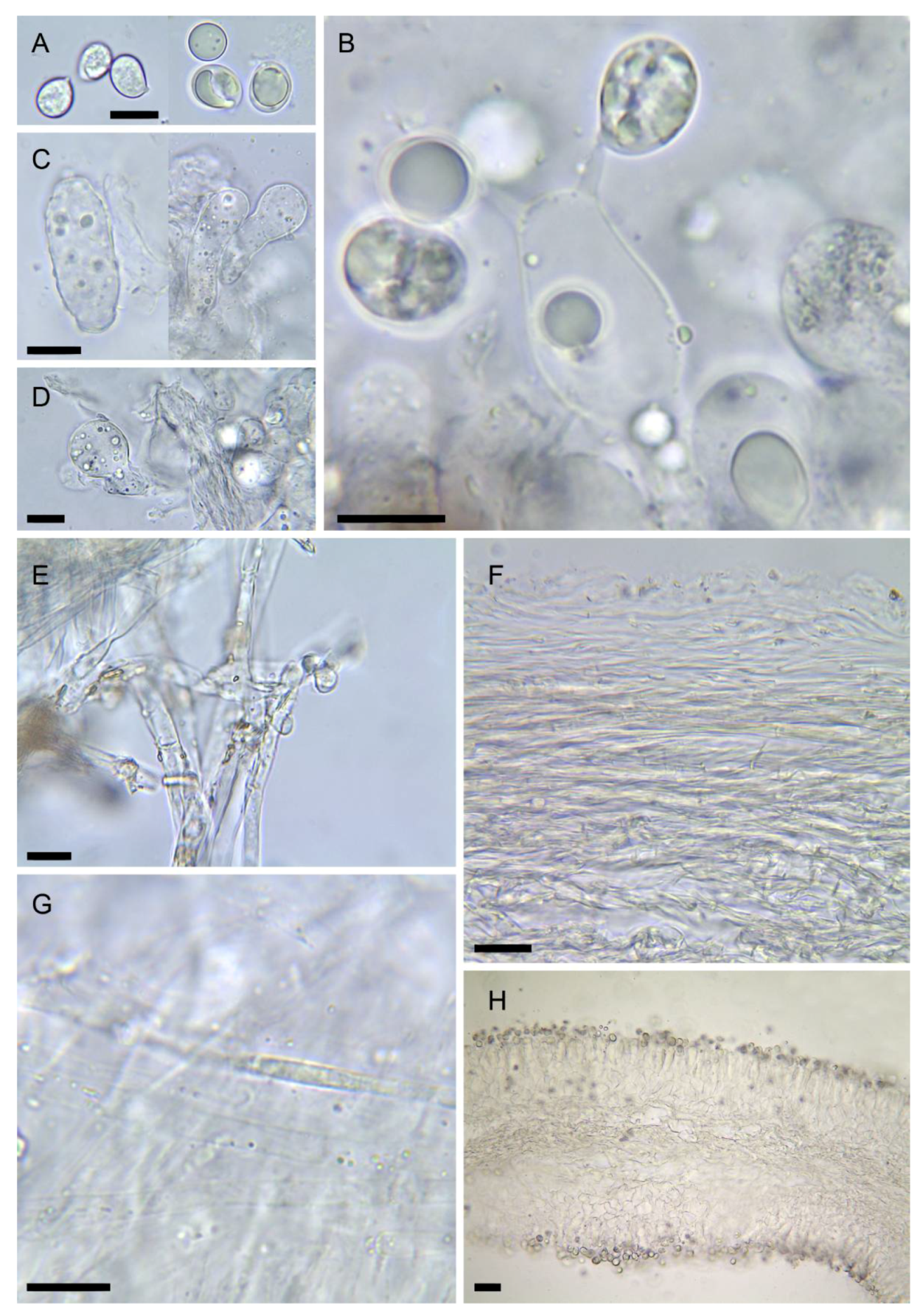
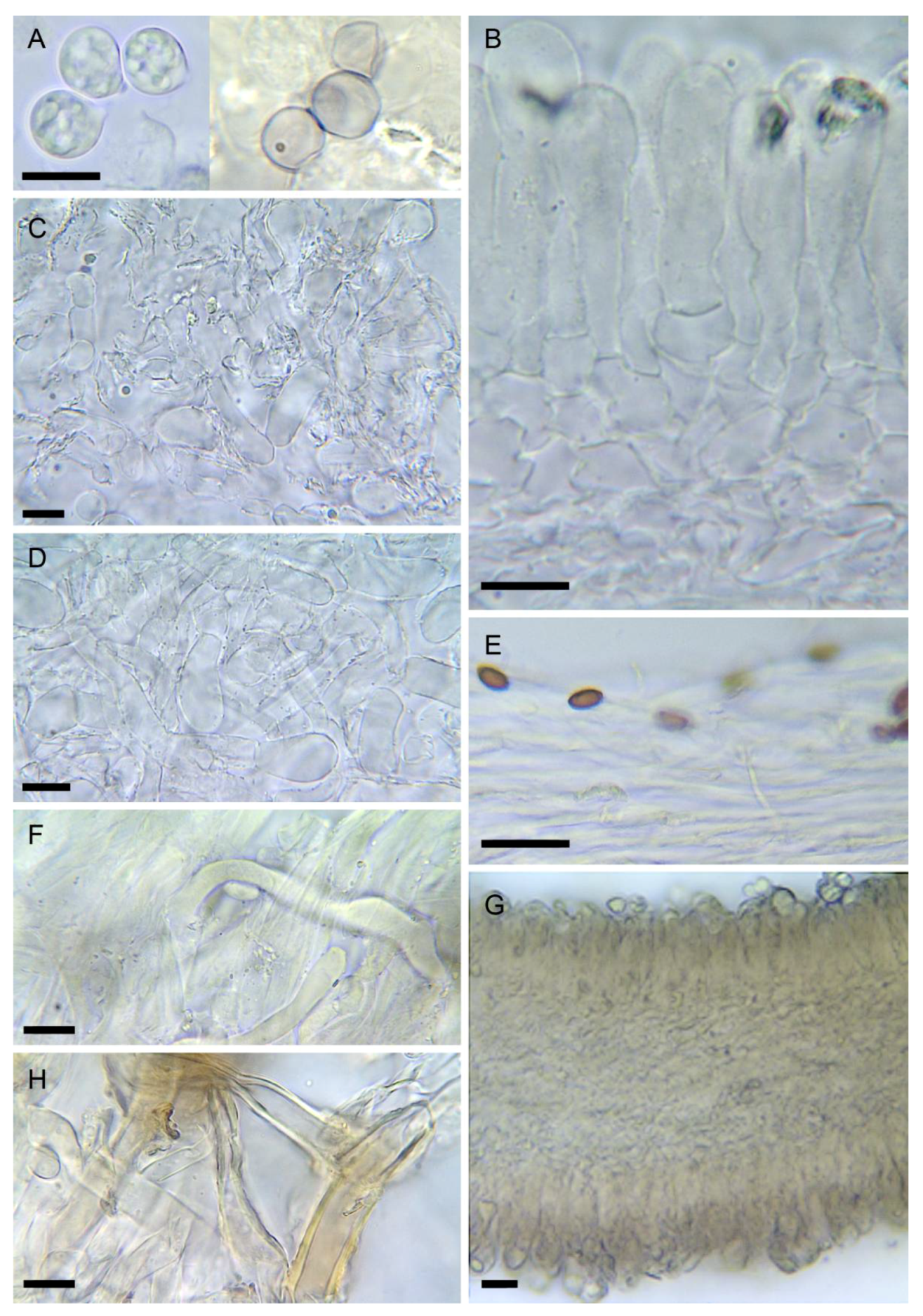
3.2. Taxonomy
- 1.
- Amanita phalloides (Vaill. ex Fr.) Link, Handb. Erk. Gew. 3: 272 (1833) Figure 2E, Figure 3E and Figure 4E
- ≡
- Agaricus phalloides Vaill. ex Fr., Syst. mycol. (Lundae) 1: 13 (1821), nom. sanct. [protonym Fungus phalloides Vaill., Bot. paris. (Paris): 74, Table 14, Figure 5 (1723), inval.]; Venenarius phalloides (Vaill. ex Fr.) Murrill, Mycologia 4(5): 240 (1912); Amanitina phalloides (Vaill. ex Fr.) E.-J. Gilbert, in Bresadola, Iconogr. mycol., Suppl. I (Milan) 27: 78 (1940)
- =
- Agaricus insidiosus Letell., Annls Sci. Nat., Bot., sér. 2 4: 35 (1835); Amanitopsis insidiosa (Letell.) Sacc., Syll. Fung. 9: 2 (1891); Amanita insidiosa (Letell.) Bigeard & Guillemin, Fl. champ. sup. France Edn 2: 14 (1913); Amanita phalloides f. insidiosa (Letell.) E.-J. Gilbert, Le genre Amanita: 41 (1918)
- =
- Amanita phalloides var. alba Costantin & L.M. Dufour, Nouv. Fl. Champ., Edn 2 (Paris): 256 (1895)
- =
- Amanita phalloides f. citrina J.E. Lange, Dansk bot. Ark. 2(3): 8 (1915)
- =
- Amanita phalloides var. pulla Killerm., Denkschr. Bayer. Botan. Ges. in Regensb. 18: 4 (1930)
- =
- Amanita phalloides var. euphalloides Maire, Mém. Soc. Sci. Nat. Maroc. 45: 103 (1937)
- =
- Amanita phalloides var. moravecii Pilát, Česká Mykol. 20(1): 25 (1966)
- =
- Amanita dunensis R. Heim ex Bon & Andary, Docums Mycol. 13(50): 13 (1983); Amanita phalloides f. dunensis R. Heim, Revue Mycol., Paris 28: 9 (1963) [nom. inval., Art. 36.1] (synonymy proposed here)
- =
- Amanita verna var. tarda Trimbach, Annales du Muséum d’Histoire Naturelle de Nice 91(3): 85 (1999); Amanita tarda (Trimbach) Contu, Boll. Gruppo Micol. ‘G. Bresadola’ (Trento) 43(2): 83 (2000)
- =
- Hypophyllum virosum Paulet Traité champ. Comest. (Paris): 2: 326 (1793) [inval., op. rejic.]
- =
- Amanita viridis Pers., Tent. disp. meth. fung. (Lipsiae): 67 (1797) [illegit., non A. viridis Huds.)]
- =
- Agaricus virosus Vittad., Descr. fung. mang. Italia: 135 (1835) [illegit., non A. virosus Sow.]
- =
- Amanita andaryi Mornand, Docums Mycol. 22(88): 12 (1993) [inval., introduced as nom. nov. based on “A. virosus var. albus Vittad. 1835: 143”, but this name does not exist in the cited book]
- 2.
- Amanita phalloides f. porrinensis (Freire & M.L. Castro) G. Moreno & Olariaga, comb. nov. Figure 2G and Figure 3EMycoBank number: MB 842794
- ≡
- Amanita porrinensis Freire & M.L. Castro, Mykes 1: 59 (1998) [also An. Jard. bot. Madr. 44(2): 533 (1987), inval., art. 39.1]
- 3.
- Amanita amerivirosa Tulloss, L.V. Kudzma & M. Tulloss, Amanitaceae 1(4): 5 (2021) Figure 2A–C, Figure 3A, Figure 4C and Figure 5
- =
- Amanita virosa var. levipes Neville & Poumarat, Fungi europ. 9: 600 (2004)
- 4.
- =
- Agaricus vernalis Bolton, Hist. fung. Halifax (Huddersfield) 2: 48, Table XLVIII (1788)
- =
- Amanita verna var. ochroleuca Forq. ex Quél., 1888, Fl. mycol. Fr.: 309; Amanita phalloides var. ochroleuca (Forq. ex Quél.) Quél. & Bataille, 1902, Fl. Amanites et lépiotes: 32; Amanita ochroleuca (Forq. ex Quél.) Bigeard & Guillemin, 1913, Fl. champ. sup. France 2nd ed.: 14
- =
- Amanita verna var. decipiens Trimbach, Riviéra Scientifique, 1970(1): 18 (1970); Amanita decipiens (Trimbach) Jacquet., Docums Mycol. 22(86): 30 (1992)
- =
- Amanita verna f. ellipticospora E.-J. Gilbert, in Bresadola, Iconogr. Mycol., Suppl. II (Milan) 27: 320 (1941) [inval., art. 38.1 and art. 39.1]
- 5.
- 6.
- =
- Amanita virosa var. aculeata Voglino, Boll. Soc. bot. ital., 1894: 120 (1894)
- Key to European species of sect. Phalloideae
- 1.
- KOH reaction bright yellow on surfaces…………………………………………………………2
- 1.
- KOH reaction none or only pale cream yellow…………………………………………………5
- 2.
- Stipe smooth. Ring complete, not lacerated at maturity, membranous, persistent on stipe, made up of mostly filamentous hyphae…………………………………………………………3
- 2.
- Stipe woolly-scaly. Ring thin, usually lacerate, hanging at margin or disappearing at maturity, mostly made up of short cylindrical elements. Smell none till late maturity, finally slightly nitrous. Spores mostly subglobose (Qav < 1.15). Moist, acidic, often peaty forests under conifers (Pinus, Picea) or broadleaved (Betula, Fagus)…………………A. virosa
- 3.
- Pileus soon flattened to depressed. Smell not perceptible in the bulb. Basidiomata white to ochraceous when dry [pileus yellowish green in ‘f. xanthoviridis’]. Subhymenium 30–45 µm thick, of puzzle-like structure. Spores mostly broadly elliptical (Qav > 1.15). Calcareous or mineral-rich soils, mostly under Quercus spp., usually in spring……………………A. verna
- 3.
- Pileus conical to campanulate……………………………………………………………………4
- 4.
- Strong smell of iodine in the bulb. Basidiomata turning uniformly lemon yellow when dry. Subhymenium poorly developed, ramose, less than 20 µm thick. Spores mostly subglobose (Qav < 1.15). Acidic, mesophilic forests, mostly under Quercus…………A. amerivirosa
- 4.
- Smell not of iodine. Basidiomata turning brownish when dry. Subhymenium well-developed, with puzzle-like elements. Spores mostly broadly ellipsoid (Qav > 1.15). Mediterranean forests…………………………………………………A. phalloides f. porrinensis
- 5.
- Fruiting in spring or early summer. KOH reaction dirty cream-yellow. Pileus white turning ochraceous at disc when old or dry, often gibbose at maturity. Under oaks on mineral-rich soils, in spring, Mediterranean…………………………………………A. vidua
- 5.
- Fruiting in autumn. KOH reaction nil or pale yellowish. Pileus usually greenish with dark fibrils (typical form), but also yellowish, ochraceous, greenish brown to pure white or with only faint greenish shade, convex to flattened but only exceptionally gibbose (A. phalloides f. porrinensis). Under various deciduous (especially Fagaceae) or coniferous (Abies, Cedrus, Pinus, Picea) trees on acidic to moderately calcareous soils, summer to early winter, Mediterranean to cold-temperate…………………………………………A. phalloides
3.3. Toxicology
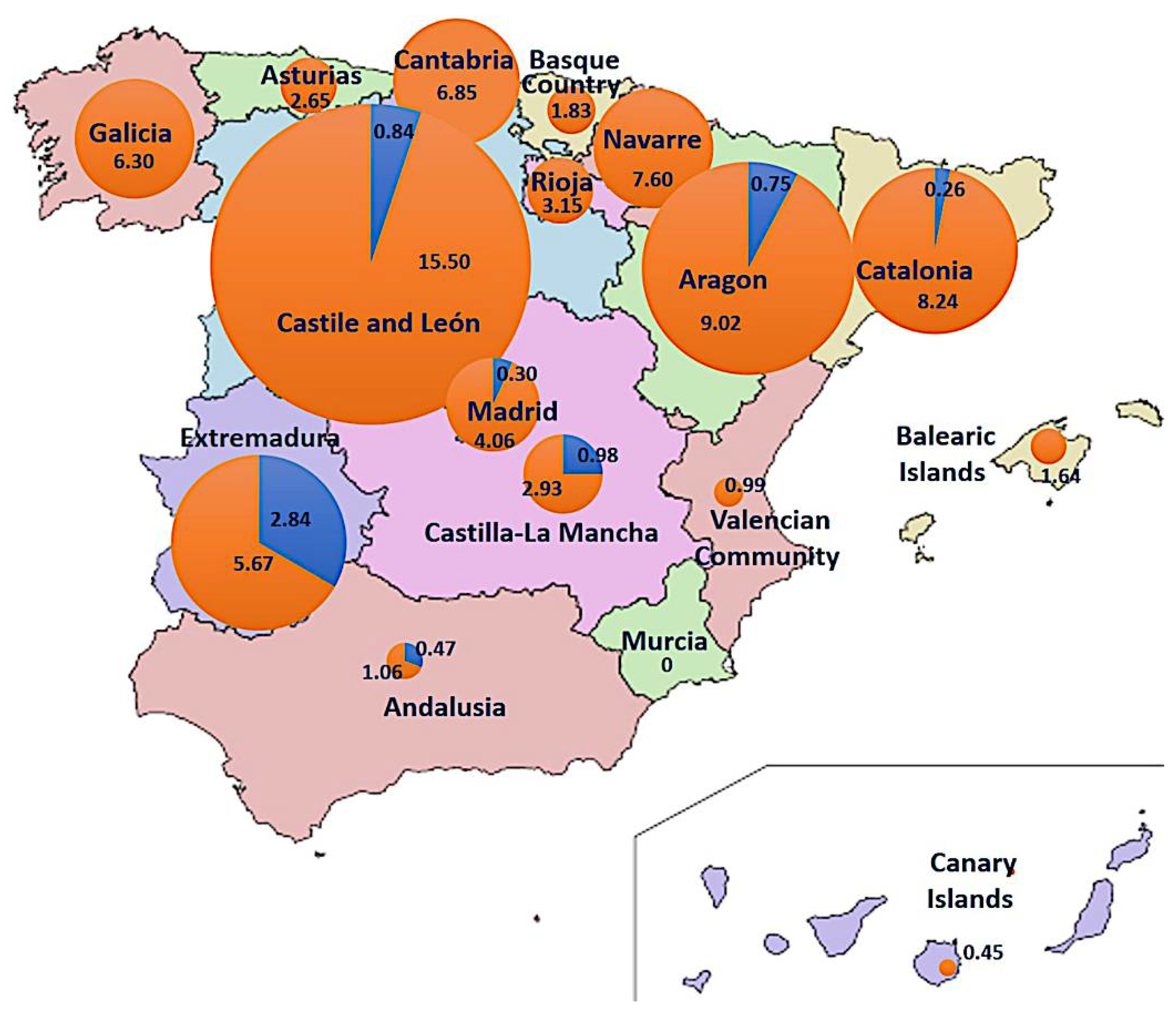
3.4. Epidemiology
4. Discussion
5. Conclusions
- (a)
- Five distinct species of Amanita section Phalloideae can be found in Mediterranean Europe: A. phalloides, A. virosa, A. verna, A. amerivirosa, and A. vidua sp. nov., a new name proposed for the KOH-negative Mediterranean species previously described as A. verna or A. decipiens by various authors.
- (b)
- Three taxa, Amanita decipiens, A. porrinensis, and A. virosa var. levipes are here considered later heterotypic synonyms of A. verna, A. phalloides, and A. amerivirosa, respectively.
- (c)
- Samples identified with the provisional name ‘Amanita verna f. xanthoviridis’ are not genetically different from A. verna f. verna, suggesting that this species can produce distinctly colored basidiomata.
- (d)
- All Mediterranean species of Amanita sect. Phalloideae contain amatoxins and phallotoxins, although their identity and quantity differ between species. The newly described A. vidua is the most amanitin-rich European species of the section. No amatoxin was detected in the analyzed samples of Amanita amerivirosa from western France.
- (e)
- In Spain, the incidence of mushroom poisonings in spring attributed to species of Amanita sect. Phalloideae is about 10 times higher in regions where other similar edible species are collected in the same season, representing up to 1/3 of all mushroom poisoning during the year in these regions.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brandenburg, W.E.; Ward, K.J. Mushroom poisoning epidemiology in the United States. Mycologia 2018, 110, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Sinno-Tellier, S.; Bruneau, C.; Daoudi, J.; Greillet, C.; Verrier, A.; Bloch, J. Surveillance nationale des intoxications alimentaires par des champignons: Bilan des cas rapportés au réseau des centres antipoison de 2010 à 2017 en France métropolitaine. Bull Epidemiol. Hebd. 2019, 33, 666–678. [Google Scholar]
- Horowitz, B.Z.; Moss, M.J. Amatoxin Mushroom Toxicity; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Vaillant, S. Botanicon Parisiense, ou Dénombrement par Ordre Alphabétique des Plantes qui se Trouvent aux Environs de Paris; Verbeek, J., Verbeek, H., Lakeman, B., Eds.; A.-C. Briasson: Leiden/Amsterdam, The Netherlands, 1727; pp. 1–205. [Google Scholar]
- Bulliard, J.B.F. Herbier de la France; Bulliard, J.B.G., Ed.; “Chez l’auteur, Didot, Debure et Belin”: Paris, France, 1782–1783; Volume 3, pp. 97–128. [Google Scholar]
- Fries, E.M. Epicrisis Systematis Mycologici; Gleerup, C.W.K., Ed.; “Typographia Academica”: Uppsala, Sweden, 1838; pp. 1–610. [Google Scholar]
- Ye, Y.; Liu, Z.; Zhao, M. CLIF-OF > 9 predicts poor outcome in patients with Amanita phalloides poisoning. Am. J. Emerg. Med. 2021, 39, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Marinov, P.; Bonchev, G.; Ivanov, D.; Zlateva, S.; Dimitrova, T.; Georgiev, K. Mushrooms intoxications. J. IMAB 2018, 24, 1887–1890. [Google Scholar] [CrossRef] [Green Version]
- Piqueras, J. Actualidades en intoxicaciones por setas. Butl. Soc. Catalana Micol. 1983, 7, 19–23. [Google Scholar]
- Pelclová, D.; Rakovcová, H. Amanita phalloides poisoning queries recorded at the toxicology information center in Prague. Cas. Lek. Ces. 1993, 132, 470–472. [Google Scholar]
- Lecot, J.; Bruneau, C.; Courtois, A.; Vodovar, D.; Landreau, A.; Le Roux, G.; D’Escatha, A. Syndrome phalloïdien: Série rétrospective de 204 cas. Toxicol. Anal. Clin. 2021, 33, S13–S39. [Google Scholar] [CrossRef]
- Cervellin, G.; Comelli, I.; Rastelli, G.; Sanchis-Gomar, F.; Negri, F.; De Luca, C.; Lippi, G. Epidemiology and clinics of mushroom poisoning in Northern Italy: A 21-year retrospective analysis. Hum. Exp. Toxicol. 2018, 37, 697–703. [Google Scholar] [CrossRef]
- Giusti, A.; Ricci, E.; Gasperetti, L.; Galgani, M.; Polidori, L.; Verdigi, F.; Narducci, R.; Armani, A. Building of an Internal Transcribed Spacer (ITS) gene dataset to support the Italian Health Service in mushroom identification. Foods 2021, 10, 1193. [Google Scholar] [CrossRef]
- Wennig, R.; Eyer, F.; Schaper, A.; Zilker, T.; Andresen-Streichert, H. Mushroom poisoning. Dtsch. Ärztebl. 2020, 117, 701–708. [Google Scholar] [CrossRef]
- Ahn, C.; Kang, H.; Lim, T.H.; Oh, J. Poisoning due to ingestion of amatoxin-containing mushrooms in South Korea: A systematic review and meta-analysis. Signa Vitae 2021, 17, 25–33. [Google Scholar]
- Keller, S.A.; Klukowska-Rötzler, J.; Schenk-Jaeger, K.M.; Kupferschmidt, H.; Exadaktylos, A.K.; Lehmann, B.; Liakoni, E. Mushroom poisoning—A 17 year retrospective study at a level I University Emergency Department in Switzerland. Int. J. Environ. Res. Public Health 2018, 15, 2855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yardan, T.; Baydin, A.; Eden, A.O.; Akdemir, H.U.; Aygun, D.; Acar, E.; Arslan, B. Wild mushroom poisonings in the Middle Black Sea region in Turkey: Analyses of 6 years. Hum. Exp. Toxicol. 2010, 29, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Sarawi, S.; Shi, Y.-N.; Lotz-Winter, H.; Reschke, K.; Bode, H.B.; Piepenbring, M. Occurrence and chemotaxonomical analysis of amatoxins in Lepiota spp. (Agaricales). Phytochemistry 2022, 195, 113069. [Google Scholar] [CrossRef]
- Enjalbert, F.; Gallion, C.; Jehl, F.; Monteil, H.; Faulstich, H. Amatoxins and phallotoxins in Amanita species: High-performance liquid chromatographic determination. Mycologia 1993, 85, 579–584. [Google Scholar] [CrossRef]
- Enjalbert, F.; Rapior, S.; Nouguier-Soulé, J.; Guillon, S.; Amouroux, N.; Cabot, C. Treatment of amatoxin poisoning: 20-year retrospective analysis. J. Toxicol. Clin. Toxicol. 2002, 40, 715–757. [Google Scholar] [CrossRef]
- Enjalbert, F.; Cassanas, G.; Rapior, S.; Renault, C.; Chaumont, J.P. Amatoxins in wood-rotting Galerina marginata. Mycologia 2004, 96, 720–729. [Google Scholar] [CrossRef]
- Xiang, H.; Zhou, Y.; Zhou, C.; Lei, S.; Yu, H.; Wang, Y.; Zhu, S. Investigation and analysis of Galerina sulcipes poisoning in a canteen. Clin. Toxicol. 2018, 56, 365–369. [Google Scholar] [CrossRef]
- Landry, B.; Whitton, J.; Bazzicalupo, A.L.; Ceska, O.; Berbee, M.L. Phylogenetic analysis of the distribution of deadly amatoxins among the little brown mushrooms of the genus Galerina. PLoS ONE 2021, 16, e0246575. [Google Scholar] [CrossRef]
- Hazani, E.; Taitelman, U.; Shasha, S.M. Amanita verna poisoning in Israel-Report of a rare case out of time and place. Arch. Toxicol. 1983, 6, 186–189. [Google Scholar]
- Yilmaz, I.; Kaya, E.; Sinirlioglu, Z.A.; Bayram, R.; Surmen, M.G.; Colakoglu, S. Clinical importance of toxin concentration in Amanita verna mushroom. Toxicon 2014, 87, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, M.; Afshari, A.; Arsene, A.L.; Mégarbane, B.; Dumanov, J.; Bastos Paoliello, M.M.; Tsatsakis, A.; Carvalho, F.; Hashemzaei, M.; Karimi, G.; et al. Toxicological profile of Amanita virosa-A narrative review. Toxicol. Rep. 2019, 6, 143–150. [Google Scholar] [CrossRef]
- Khatir, I.G.; Hosseininejad, S.M.; Ghasempouri, S.K.; Asadollahpoor, A.; Moradi, S.; Jahanian, F. Demographic and epidemiologic evaluation of mushroom poisoning: A retrospective study in 4-year admissions of Razi Hospital (Qaemshahr, Mazandaran, Iran). Med. Glas. 2020, 17, 117–122. [Google Scholar]
- Petekkaya, S.; Börk, T.; Ayaz, N.; Göktürk, C.; Şamdancı, E.; Celbiş, O. Fatal mushroom poisoning in Syrian refugees. Azerbaijan Med. Assoc. J. 2016, 1, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Euronews. Available online: https://www.euronews.com/2021/09/02/afghan-refugee-5-dies-after-eating-poisonous-mushrooms-in-poland (accessed on 2 September 2021).
- Preston, J.F.; Stark, H.J.; Kimbrough, J.W. Quantitation of amanitins in Amanita verna with calf thymus RNA polymerase B. Lloydia 1975, 38, 153–161. [Google Scholar]
- Cai, Q.; Tulloss, R.E.; Tang, L.P.; Tolgor, B.; Zhang, P.; Chen, Z.H.; Yang, Z.L. Multi-locus phylogeny of lethal amanitas: Implications for species diversity and historical biogeography. BMC Evol. Biol. 2014, 14, 143. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Cui, Y.-Y.; Yang, Z.L. Lethal Amanita species in China. Mycologia 2016, 108, 993–1009. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.-Y.; Cai, Q.; Tang, L.-P.; Liu, J.-W.; Yang, Z.L. The family Amanitaceae: Molecular phylogeny, higher-rank taxonomy and the species in China. Fungal Divers. 2018, 91, 5–230. [Google Scholar] [CrossRef]
- Codjia, J.E.I.; Cai, Q.; Zhou, S.W.; Luo, H.; Ryberg, M.; Yorou, N.S.; Yang, Z.L. Morphology, multilocus phylogeny, and toxin analysis reveal Amanita albolimbata, the first lethal Amanita species from Benin, West Africa. Front. Microbiol. 2020, 11, 599047. [Google Scholar] [CrossRef]
- Fraiture, A.; Amalfi, M.; Raspé, O.; Kaya, E.; Akata, I.; Degreef, J. Two new species of Amanita sect. Phalloideae from Africa, one of which is devoid of amatoxins and phallotoxins. MycoKeys 2019, 53, 93–125. [Google Scholar]
- Codjia, J.E.I.; Wang, P.M.; Ryberg, M.; Yorou, N.S.; Yang, Z.L. Amanita sect. Phalloideae: Two interesting non-lethal species from West Africa. Mycol. Progr. 2022, 21, 39. [Google Scholar]
- Tulloss, R.E.; Kudzma, L.V.; Tulloss, M.K.; Rockefeller, A. Amanita amerivirosa—A new toxic North American species of Amanita section Phalloideae. Amanitaceae 2021, 1, 1–15. [Google Scholar]
- Neville, P.; Poumarat, S. Amaniteae. Amanita, Limacella & Torrendia. Fungi Europaei; Candusso, M., Ed.; Edizioni Candusso: Alassio, Italy, 2004; pp. 1–1120. [Google Scholar]
- Bas, C. Morphology and subdivision of Amanita and a monograph on its section Lepidella. Persoonia 1969, 5, 385–579. [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- Mullis, K.; Faloona, F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes-Application to the identification of mycorrhizae and rust. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically ampliWed ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Cubeta, M.A.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- van Tuinen, D.; Zhao, B.; Gianinazzi-Pearson, V. PCR studies of AM fungi, from studies to application. In Mycorrhiza Manual; Varma, A., Ed.; Springer: Heidelberg, Germany, 1998; pp. 387–399. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Weiß, M.; Yang, Z.L.; Oberwinkler, F. Molecular phylogenetic studies in the genus Amanita. Can. J. Bot. 1998, 76, 1170–1179. [Google Scholar]
- Oda, T.; Tanaka, C.; Tsuda, M. Molecular phylogeny of Japanese Amanita species based on nucleotide sequences of the internal transcribed spacer region of nuclear ribosomal DNA. Mycoscience 1999, 40, 57–64. [Google Scholar] [CrossRef]
- Drehmel, D.; Moncalvo, J.M.; Vilgalys, R. Molecular phylogeny of Amanita based on large-subunit ribosomal DNA sequences: Implications for taxonomy and character evolution. Mycologia 1999, 91, 610–618. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, J.; Yang, Z. Molecular phylogeny of eastern Asia species of Amanita (Agaricales, Basidiomycota): Taxonomic and biogeographic implications. Fungal Divers. 2004, 17, 219–238. [Google Scholar]
- Zhang, P.; Chen, Z.H.; Xiao, B.; Tolgor, B.; Bao, H.Y.; Yang, Z.L. Lethal amanitas of East Asia characterized by morphological and molecular data. Fungal Divers. 2010, 42, 119–133. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP *. Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2003; Version 4. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Enjalbert, F.; Cassanas, G.; Salhi, S.L.; Guinchard, C.; Chaumont, J.P. Distribution of the amatoxins and phallotoxins in Amanita phalloides. Influence of the tissues and the collection site. Comptes Rendus Séances Acad. Sci. Sér. 3 Sci. Vie 1999, 322, 855–862. [Google Scholar] [CrossRef]
- Kaya, E.; Yilmaz, I.; Sinirlioglu, Z.A.; Karahan, S.; Bayram, R.; Yaykasli, K.O.; Colakoglu, S.; Saritas, A.; Severoglu, Z. Amanitin and phallotoxin concentration in Amanita phalloides var. alba mushroom. Toxicon 2013, 76, 225–233. [Google Scholar] [CrossRef]
- Enjalbert, F.; Gallion, C.; Jehl, F.; Monteil, H. Simultaneous assay for amatoxins and phallotoxins in Amanita phalloides Fr. by high-performance liquid chromatography. J. Chromatogr. 1992, 598, 227–236. [Google Scholar] [CrossRef]
- McKnight, T.A.; McKnight, K.B.; Skeels, M.C. Amatoxin and phallotoxin concentration in Amanita bisporigera spores. Mycologia 2010, 102, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Sgambelluri, R.M.; Epis, S.; Sassera, D.; Luo, H.; Angelos, E.R.; Walton, J.D. Profiling of amatoxins and phallotoxins in the genus Lepiota by liquid chromatography combined with UV absorbance and mass spectrometry. Toxins 2014, 6, 2336–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, S.; Fons, F.; Rapior, S.; Dubois, V.; Vitou, M.; Portet, K.; Doré, J.-C.; Poucheret, P. Decision-making for the detection of amatoxin poisoning: A comparative study of standard analytical methods. Cryptogam. Mycol. 2016, 37, 217–239. [Google Scholar] [CrossRef]
- Davison, E.M.; Giustiniano, D.; Busetti, F.; Gates, G.M.; Syme, K. Death cap mushrooms from southern Australia: Additions to Amanita (Amanitaceae, Agaricales) section Phalloideae Clade IX. Aust. Syst. Bot. 2017, 30, 371–389. [Google Scholar] [CrossRef]
- Morini, S.; Gennari, A.; Agnello, C. Amanita sabulicola, una nuova specie descritta dal Salento (Puglia-Italia). Riv. Micol. 2020, 62, 239–251. [Google Scholar]
- Davison, E.M.; Giustiniano, D.; Bougher, N.L.; McGurk, L.E.; Watkin, E.L.J. Additions to Amanita (Amanitaceae, Agaricales) section Arenariae from south-western Australia. Aust. Syst. Bot. 2021, 34, 541–569. [Google Scholar] [CrossRef]
- Marchand, A. Champignons du Nord et du Midi. Tome 1: Les Meilleurs Comestibles et les Principaux Vénéneux; Marchand, A., Ed.; Diffusion Hachette: Perpignan, France, 1971; pp. 1–282. [Google Scholar]
- Moreno, G.; Manjón, J.L. Guía de Hongos de la Península Ibérica; Omega: Barcelona, Spain, 2010; pp. 1–1417. [Google Scholar]
- Freire, L.; Castro, M.L. Nueva especie del género “Amanita” Anales Jard. Bot. Madrid 1987, 44, 533–534. [Google Scholar]
- Castro, M.L. Amanita porrinensis L. Freire et M.L. Castro, estudio comparativo con otros taxons da sección Phalloideae (Fr.) Quél. Mykes 1998, 1, 57–59. [Google Scholar]
- Neville, P.; Poumarat, S.; Monterumici, G. Una rara Amanita della sezione Phalloideae, nuova per l’Italia: Amanita porrinensis. Boll. Gr. Micol. G. Bresadola Trento 2000, 43, 143–150. [Google Scholar]
- Miceli, A. Amanita porrinensis, una specie rarissima ritrovata sui Monti Peloritani. MicoPonte 2019, 12, 27–33. [Google Scholar]
- Gasparini, F.; Serafin, F. La quarta Amanita mortale. Boll. Gr. Micol. G. Bresadola Trento Sez. Vincenza 2012, 32, 40–44. [Google Scholar]
- Moreno, G.; Platas, G.; Peláez, F.; Bernedo, M.; Vargas, A.; Daza, A.; Santamaría, C.; Camacho, M.; de la Osa, L.R.; Manjón, J.L. Molecular phylogenetic analysis shows that Amanita ponderosa and A. curtipes are distinct species. Mycol. Progr. 2008, 7, 41–47. [Google Scholar] [CrossRef]
- Romagnesi, H. Contribution à la solution du problème d’Amanita verna Bull. Bull. Soc. Mycol. Franc. 1984, 100, 237–241. [Google Scholar]
- Konrad, P.; Maublanc, A. Icones Selectae Fungorum, Vols. I-IV, Encyclopédie Mycologique 1; P. Lechevallier: Paris, France, 1924–1937. [Google Scholar]
- Bataille, F. Les réactions macrochimiques chez les champignons: Suivies d’indications sur la morphologie des spores. Bull. Soc. Mycol. France 1947, 63 (Suppl. Sl), 1–172. [Google Scholar]
- Gilbert, E.-J. Iconographia mycologica, Amanitaceae. Iconogr. Mycol. 1941, 27 (Suppl. Sl), 203–427. [Google Scholar]
- Malençon, G.; Bertault, R. Flore des champignons supérieurs du Maroc, tome 1. Trav. Inst. Sci. Chérifien Sér. Bot. Biol. 1970, 32, 1–601. [Google Scholar]
- Bertault, R. Amanites du Maroc (deuxième contribution). Bull. Soc. Mycol. Franc. 1965, 81, 345–371. [Google Scholar]
- Bertault, R. Amanites du Maroc. Bull. Soc. Mycol. Franc. 1964, 80, 364–384. [Google Scholar]
- Raumi, M. Amanita verna f. xantho-viridis, ad Interim. Available online: www.gruppomicologicocecinese.it/SchedaFunghi.php?id=933 (accessed on 1 March 2022).
- Bertault, R. Amanites du Maroc (troisième contribution et additions). Bull. Soc. Mycol. Franc. 1980, 96, 271–286. [Google Scholar]
- Trimbach, J. Amanita verna variété decipiens n. var. Riviera Sci. 1970, 1, 15–18. [Google Scholar]
- La Chiusa, L. Amanite bianche tossiche. Boll. Gr. Micol. G. Bresadola Trento 2000, 43, 109–120. [Google Scholar]
- Contu, M. Saggio di una ciave per la determinazione delle specie del genere Amanita osservate in Sardegna. Boll. Gr. Micol. G. Bresadola Trento 2000, 43, 67–86. [Google Scholar]
- Tang, S.; Zhou, Q.; He, Z.; Luo, T.; Zhang, P.; Cai, Q.; Yang, Z.; Chen, J.; Chen, Z. Cyclopeptide toxins of lethal amanitas: Compositions, distribution and phylogenetic implication. Toxicon 2016, 120, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Wu, J.; Chen, J.; Wu, B.; He, Z.; Zhang, P.; Li, H.; Sun, C.; Liu, C.; Chen, Z.; et al. Determination of cyclopeptide toxins in Amanita subpallidorosea and Amanita virosa by high-performance liquid chromatography coupled with high-resolution mass spectrometry. Toxicon 2017, 133, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, J. The Cyclic Peptide Toxins of Amanita and other Poisonous Mushrooms. Chapter 3. In Distribution and Taxonomic Variation in the Amanita Cyclic Peptide Toxins; Springer International Publishing AG: New York, NY, USA, 2018; pp. 59–91. [Google Scholar]
- Beutler, J.A.; Der Marderosian, A.H. Chemical variation in Amanita. J. Nat. Prod. 1981, 44, 422–431. [Google Scholar] [CrossRef]
- Bonnet, M.S.; Basson, P.W. The toxicology of Amanita virosa: The destroying angel. Homeopathy 2004, 93, 216–220. [Google Scholar] [CrossRef]
- Ahmed, W.H.A.; Gonmori, K.; Suzuki, M.; Watanabe, K.; Suzuki, O. Simultaneous analysis of α-amanitin, β-amanitin, and phalloidin in toxic mushrooms by liquid chromatography coupled to time-of-flight mass spectrometry. Forensic Toxicol. 2010, 28, 69–76. [Google Scholar] [CrossRef]
- Garcia, J.; Oliveira, A.; de Pinho, P.G.; Freitas, V.; Carvalho, A.; Baptista, P.; Pereira, E.; Bastos, M.D.L.; Carvalho, F. Determination of amatoxins and phallotoxins in Amanita phalloides mushrooms from northeastern Portugal by HPLC-DAD-MS. Mycologia 2015, 107, 679–687. [Google Scholar] [CrossRef] [Green Version]
- Bon, M.; Andary, C. A new species of Amanita: Amanita dunensis Heim ex Bon et Andary. Doc. Mycol. 1983, 50, 13–14. [Google Scholar]
- Lamarck, J.-B.M. Encyclopédie Méthodique. Botanique. Tome Premier, Partie 1; Panckoucke: Paris, France, 1783; pp. 1–344. [Google Scholar]
- Fries, E.M. Systema Mycologicum I; “Ex Officina Berlingiana”: Lund, Sweden, 1821. [Google Scholar]
- Russi, M.; Josserand, M. Etude sur Amanita verna (Bull.) Persoon (Basidiomycète Agaricales) récoltée dans la région lyonnaise. Bull. Mens. Soc. Linn. Lyon 1983, 52, 6–10. [Google Scholar] [CrossRef]
- Trimbach, J. Amanita verna (Bull. ex Fr.) ss. str. et ses variétés. Ann. Mus. Hist. Nat. Nice 1972, 72, 83–86. [Google Scholar]
- Stijve, T.; Seeger, R. Determination of α-, ß- and γ-amanitin by high performance thin-layer chromatography in Amanita phalloides (Vaill. ex Fr.) Secr. from various origin. Z. Naturforsch. 1979, 34, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Hallen, H.E.; Adams, G.C.; Eicker, A. Amatoxins and phallotoxins in indigenous and introduced South African Amanita species. S. Afr. J. Bot. 2002, 68, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Kaya, E.; Hancı, M.; Karahan, S.; Bayram, S.; Yaykaşlı, K.O.; Sürmen, M.G. Alfa amanitinin su ve metanoldeki termostabilitesi. Eur. J. Basic Med. Sci. 2012, 2, 106–111. [Google Scholar] [CrossRef]
- Sharma, S.; Aydin, M.; Bansal, G.; Kaya, E.; Singh, R. Determination of amatoxin concentration in heat-treated samples of Amanita phalloides by high-performance liquid chromatography: A forensic approach. J. Forensic Leg. Med. 2021, 78, 102111. [Google Scholar] [CrossRef] [PubMed]
- Hervé, H.; Mabon, G. Une amanite printanière à la Toussaint! Amanita decipiens (Trimbach) Jacquetant. Cahiers Mycol. Nantais 1996, 8, 3–9. [Google Scholar]
- Berger, C. Surprenantes poussées d’une amanite blanche mortelle Amanita virosa ss. auct. améric. Cahiers Mycol. Nantais 2002, 14, 5–6. [Google Scholar]
- Surault, J.-L. Récoltes intéressantes en 1998. Bull. Soc. Mycol. Poitou 1999, 22, 31–35. [Google Scholar]
- Dayrou, G.; Rioult, J.-P.; Shorten, D. Amanita virosa var. levipes Neville et Poumarat: Une espèce en progression dans l’Ouest de la France. Bull. Ann. Féd. Assoc. Mycol. Ouest 2018, 7, 25–34. [Google Scholar]
- Villa, A. Phytolistes, mycoliste: Les listes d’experts en appui des toxicologues. Toxicol. Anal. Clin. 2018, 30, 166–167. [Google Scholar] [CrossRef]
- Gravito Henriques, J.L. Estudo sobre casos de intoxicação por ingestão de cogumelos silvestres de Primavera: Equívocos entre Amanita ponderosa e Amanita boudieri. AÇAFA On-Line 2019, 12, 272–294. [Google Scholar]
- Kirchmair, M.; Carrilho, P.; Pfab, R.; Haberl, B.; Felgueiras, J.; Carvalho, F.; Cardoso, J.; Melo, I.; Vinhas, J.; Neuhauser, S. Amanita poisonings resulting in acute, reversible renal failure: New cases, new toxic Amanita mushrooms. Nephrol. Dial. Transplant. 2012, 27, 1380–1386. [Google Scholar] [CrossRef] [PubMed]
- Centro Nacional de Epidemiología (CNE). Resultados de la Vigilancia Epidemiologica de las Enfermedades Tasmisibles. Informe Annual, años 2017–2018; Instituto de Salud Carlos III: Madrid, Spain, 2020. [Google Scholar]
- Pennisi, L.; Lepore, A.; Gagliano-Candela, R.; Santacroce, L.; Charitos, I.A. A report on mushrooms poisonings in 2018 at the Apulian Regional Poison Center. Maced. J. Med. Sci. 2020, 8E, 616–622. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | A. verna KOH+ | A. verna KOH- |
|---|---|---|
| Bulliard (1783) | Undefined | |
| Konrad & Maublanc (1934) | A. phalloides f. alba | |
| Bataille (1945) | Not cited | A. verna |
| Bertault (1965) | Not cited | A. verna |
| Trimbach (1970) | A. verna var. decipiens | A. verna/A. verna var. tarda |
| Romagnesi (1984) | A. verna | A. verna f. ellipsospora |
| Contu (2000) | A. decipiens | A. verna/A. tarda |
| Neville & Poumarat (2005) | A. verna var. decipiens | A. verna |
| Present study | A. verna | A. vidua |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado, P.; Gasch-Illescas, A.; Morel, S.; Dagher-Kharrat, M.B.; Moreno, G.; Manjón, J.L.; Carteret, X.; Bellanger, J.-M.; Rapior, S.; Gelardi, M.; et al. Amanita Section Phalloideae Species in the Mediterranean Basin: Destroying Angels Reviewed. Biology 2022, 11, 770. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050770
Alvarado P, Gasch-Illescas A, Morel S, Dagher-Kharrat MB, Moreno G, Manjón JL, Carteret X, Bellanger J-M, Rapior S, Gelardi M, et al. Amanita Section Phalloideae Species in the Mediterranean Basin: Destroying Angels Reviewed. Biology. 2022; 11(5):770. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050770
Chicago/Turabian StyleAlvarado, Pablo, Antonia Gasch-Illescas, Sylvie Morel, Magda Bou Dagher-Kharrat, Gabriel Moreno, José Luis Manjón, Xavier Carteret, Jean-Michel Bellanger, Sylvie Rapior, Matteo Gelardi, and et al. 2022. "Amanita Section Phalloideae Species in the Mediterranean Basin: Destroying Angels Reviewed" Biology 11, no. 5: 770. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11050770