
Mesenchymal–Epithelial Transition in Fibroblasts of Human Normal Lungs and Interstitial Lung Diseases

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture
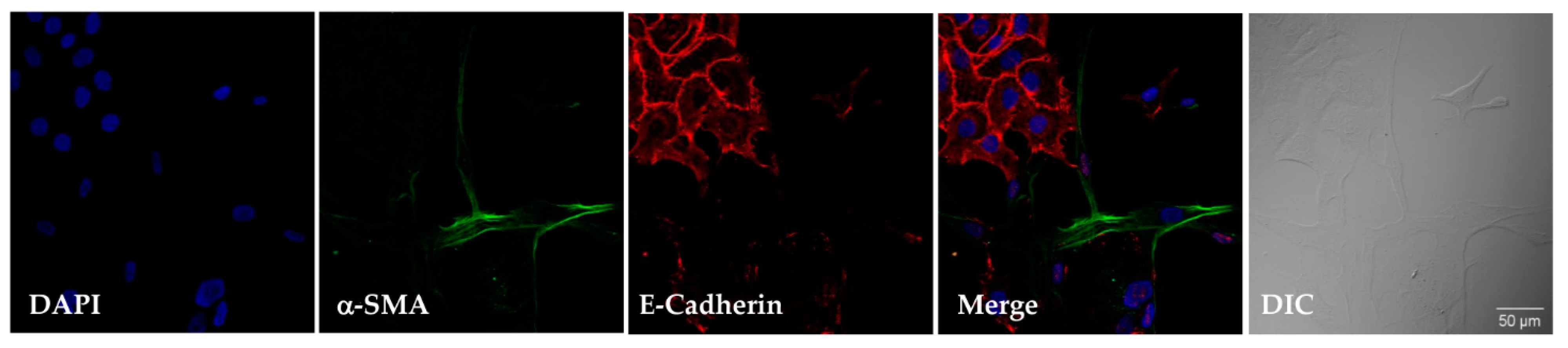
2.3. Immunofluorescence and Image Acquisition
2.4. Immunocytochemistry and Image Acquisition
2.5. Immunoblotting
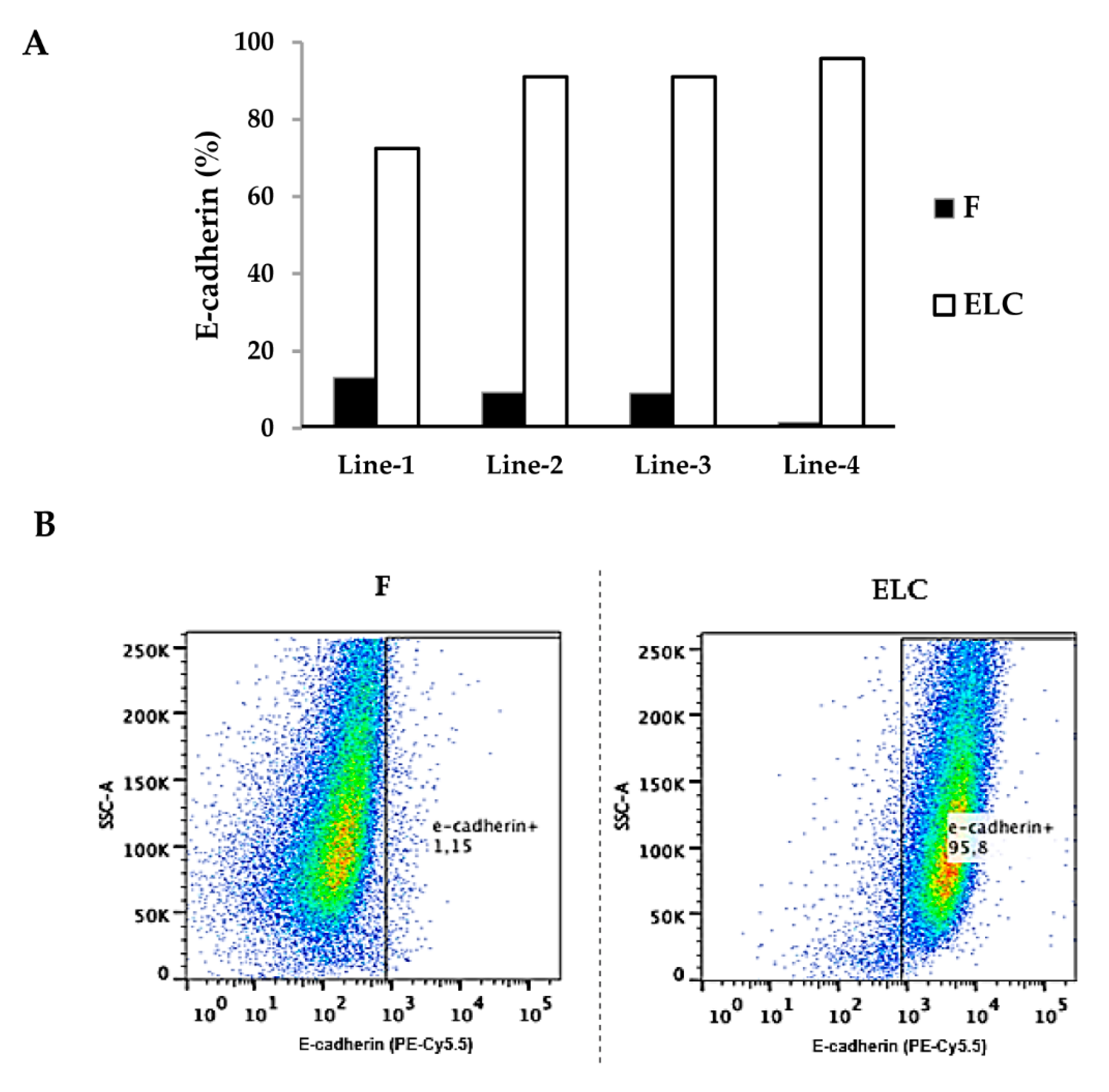
2.6. Flow Cytometry Analysis
2.7. Gelatin Zymography
2.8. Microarray Analysis
2.9. Ingenuity Pathway Analysis (IPA)
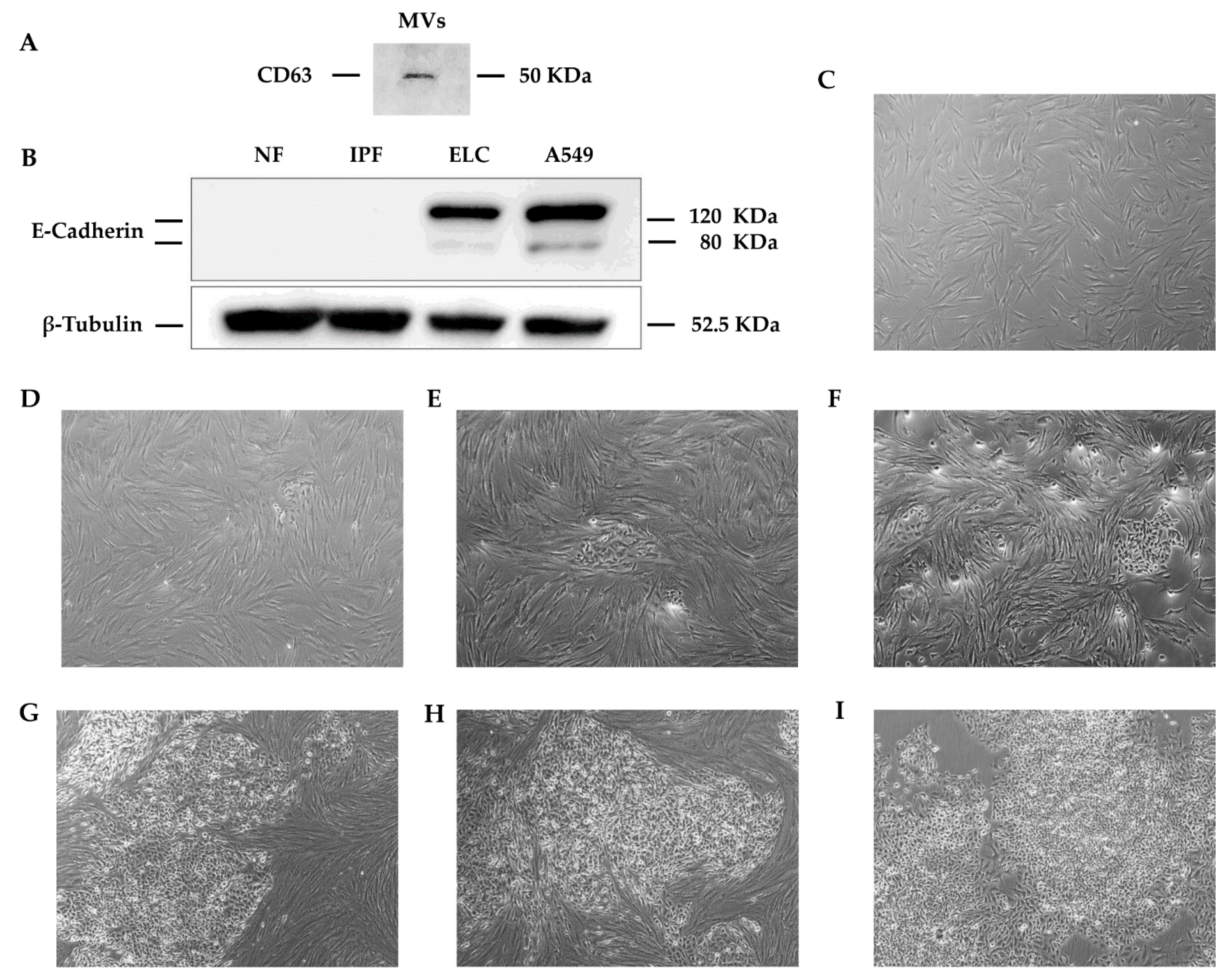
2.10. Isolation of Macrovesicles (MVs)
2.11. Quantitative RT-PCR of MicroRNAs from MVs
2.12. Statistical Analysis
3. Results
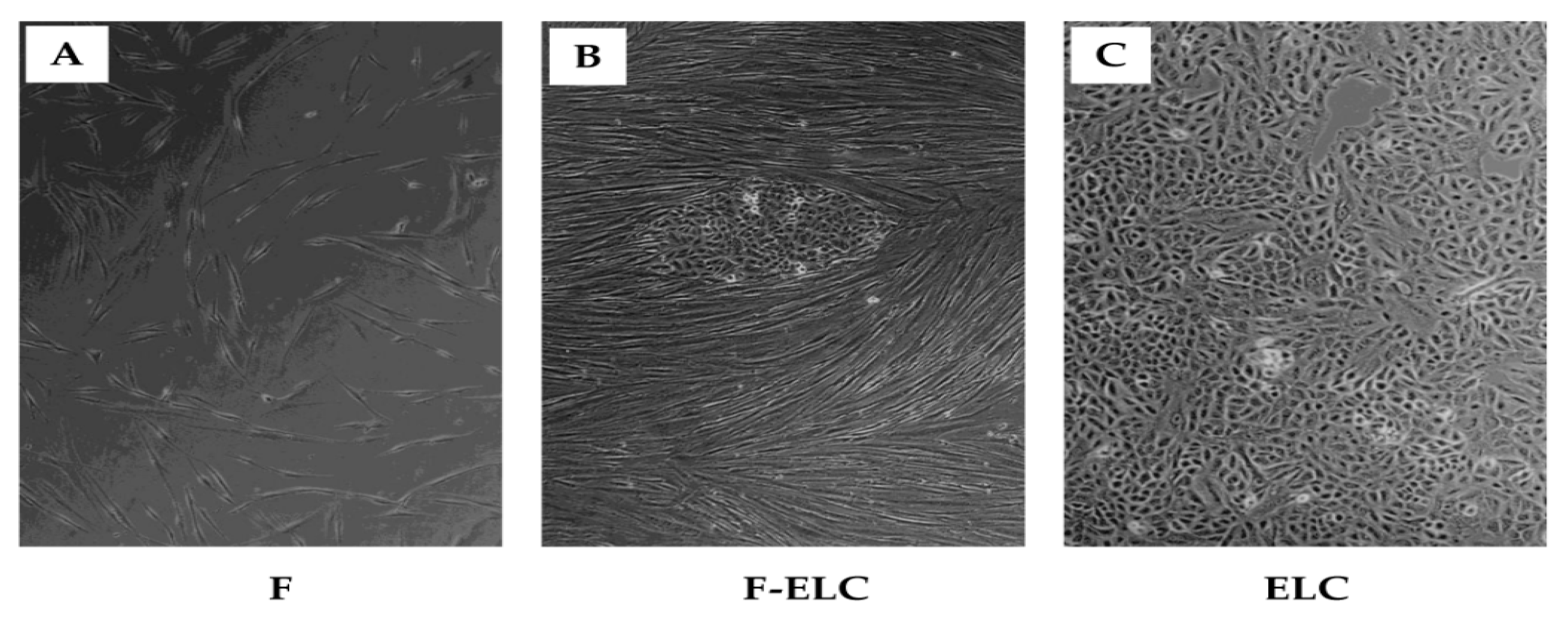
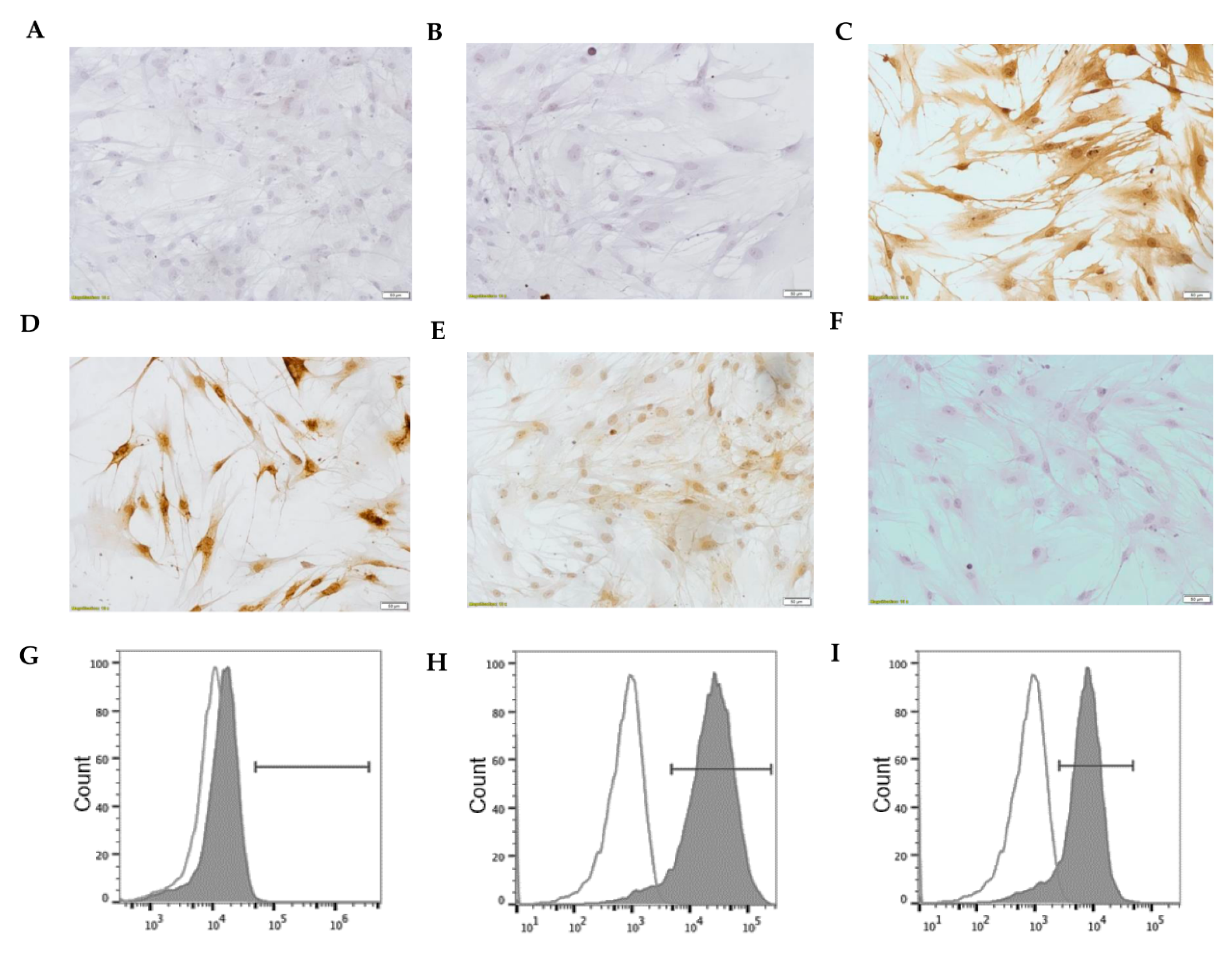
3.1. Spontaneous Changes in Human Lung Fibroblasts in Culture
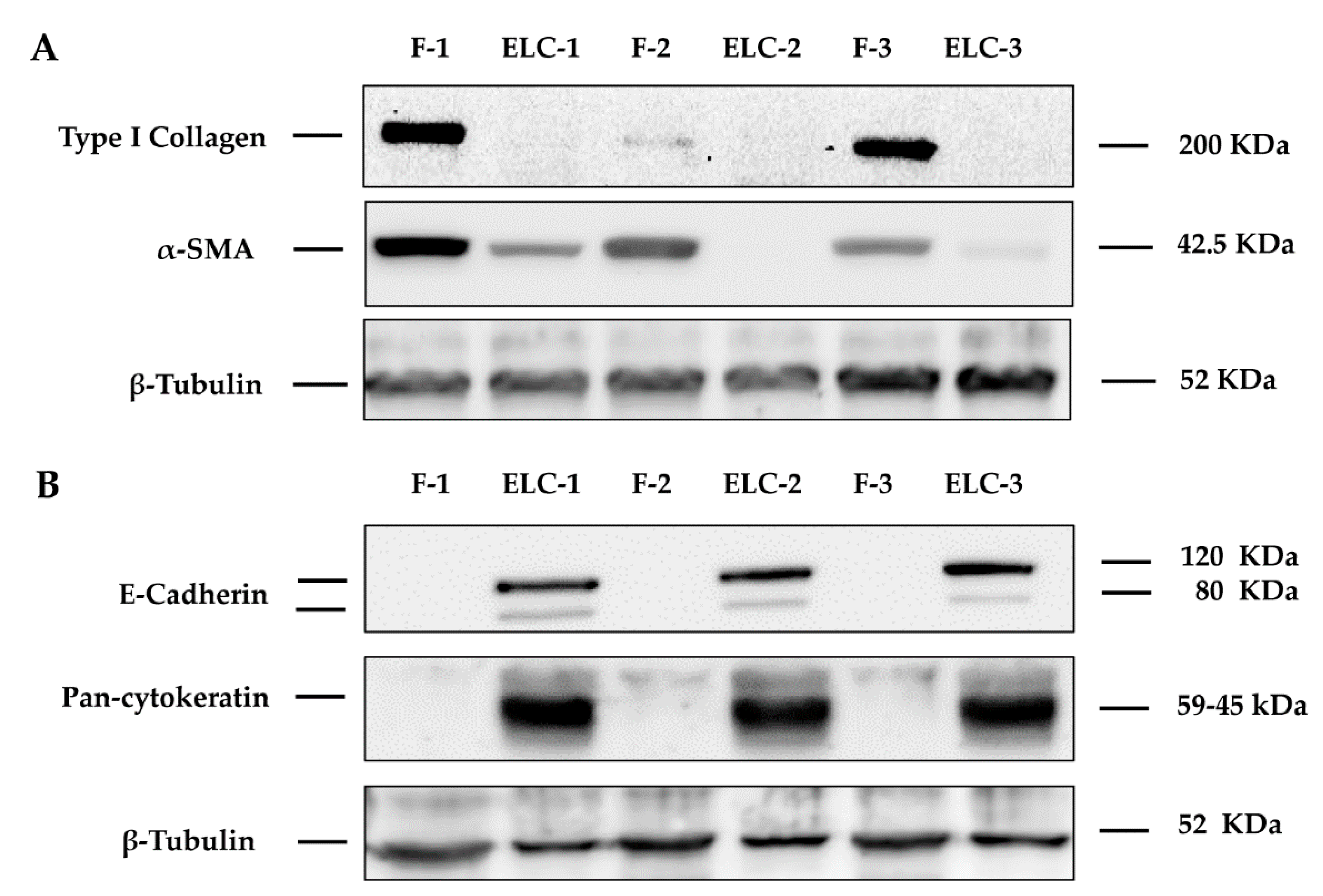
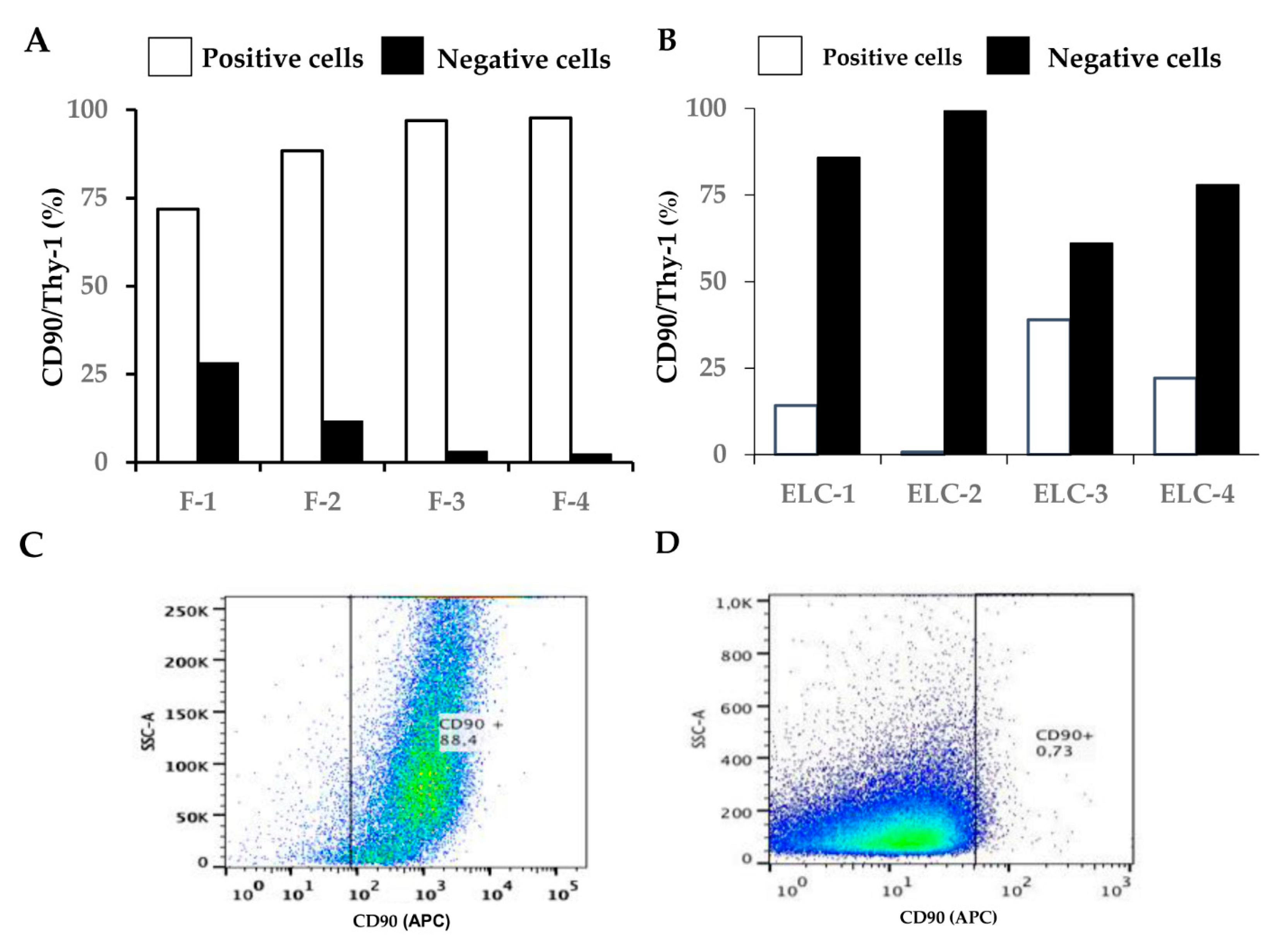
3.2. ELC Overexpresses Epithelial Biomarkers and Downregulates Mesenchymal Biomarkers
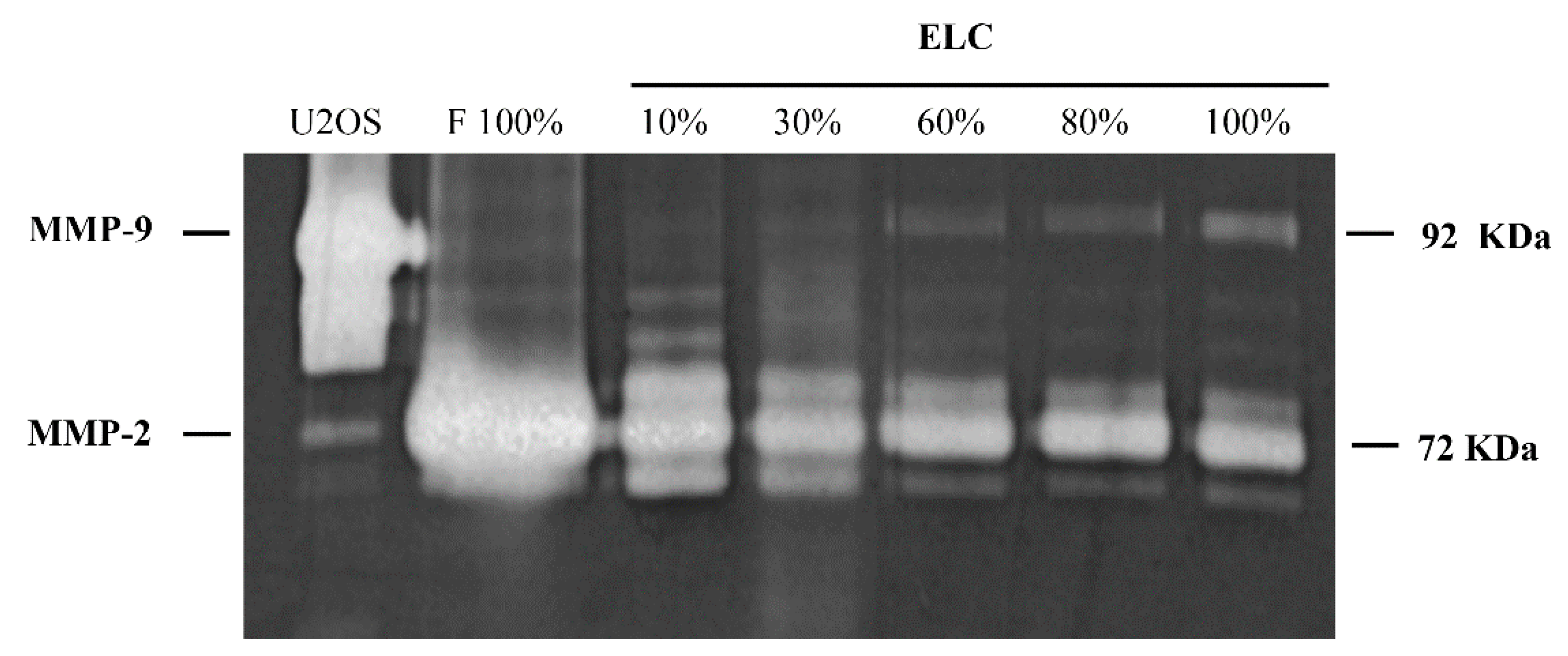
3.3. Increased MMP-9 and Decreased MMP-2 Activities in ELC
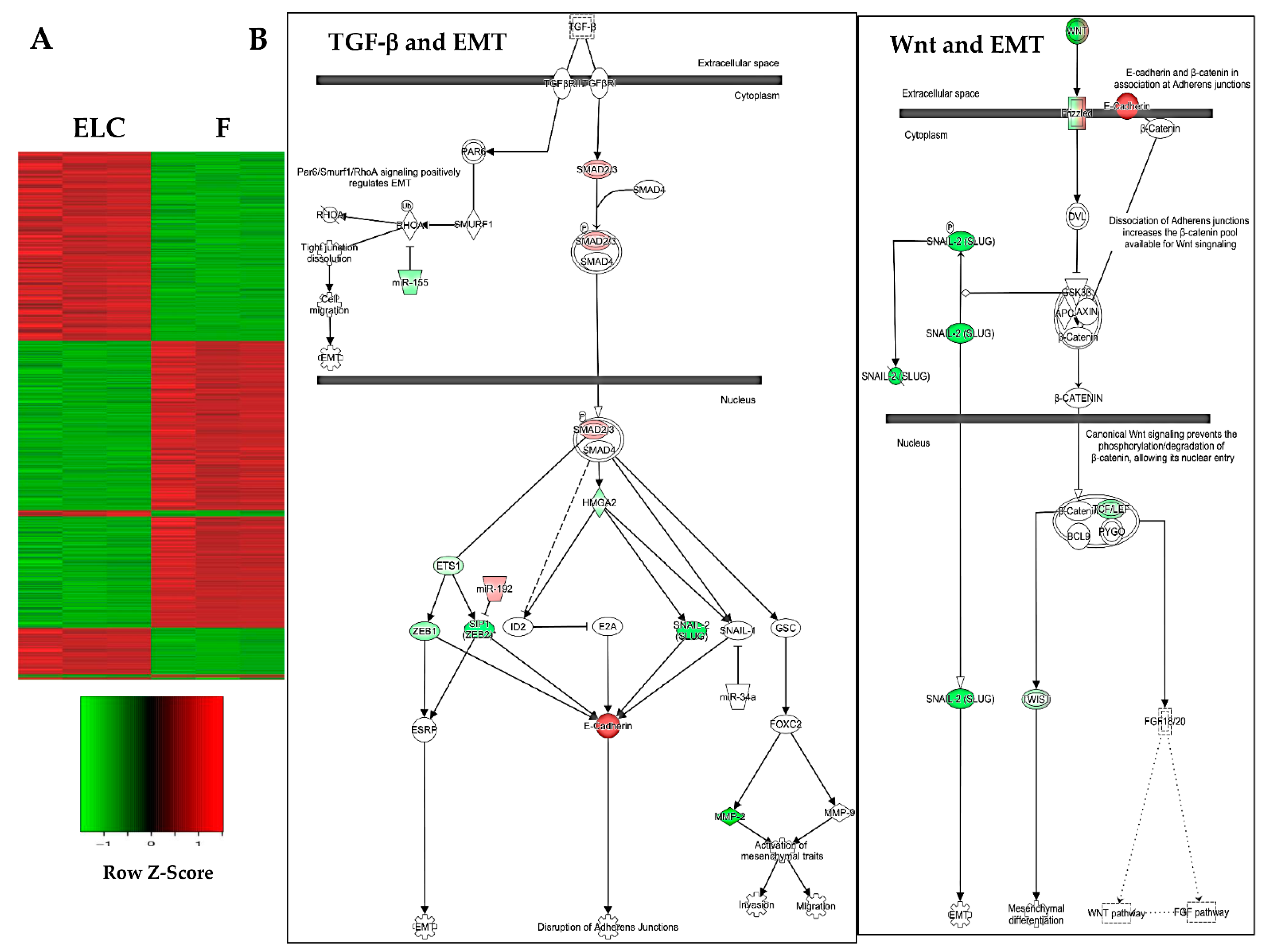
3.4. Microarray Analysis Shows Changes in the Pattern of Gene Expression
3.5. Canonical Pathways Related to MET Are Downregulated in ELC
3.6. Secreted MVs from ELC Induce a MET-Like Process in Human Lung Fibroblast
3.7. MVs from ELC Showed miRNAs Specific and Downregulated, But One Overexpressed Compared to Fibroblast
3.8. Characterization of Human Lung Fibroblast Phenotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phan, S.H. Biology of Fibroblasts and Myofibroblasts. Proc. Am. Thorac. Soc. 2008, 5, 334–337. [Google Scholar] [CrossRef]
- Ramos, C.; Montaño, M.; García-Alvarez, J.; Ruiz, V.; Uhal, B.D.; Selman, M.; Pardo, A. Fibroblasts from idiopathic pulmo-nary fibrosis and normal lungs differ in growth rate, apoptosis, and tissue inhibitor of metalloproteinases expression. Am. J. Respir. Cell. Mol. Biol. 2001, 24, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Chen, D.; Zhou, Y.; Han, Z.; Che, G. Fibroblast phenotypes in different lung diseases. J. Cardiothorac. Surg. 2014, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Liang, J.; Ni, S.; Zhou, T.; Qing, X.; Li, H.; He, W.; Chen, J.; Li, F.; Zhuang, Q.; et al. A mesenchymal-to-epithelial tran-sition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell. Stem. Cell. 2010, 7, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Pei, D. The function and regulation of mesenchymal-to-epithelial transition in somatic cell reprogramming. Curr. Opin. Genet. Dev. 2014, 28, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Dai, C.; Peng, S. Mechanism of the Mesenchymal–Epithelial Transition and Its Relationship with Metastatic Tumor Formation. Mol. Cancer Res. 2011, 9, 1608–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Høffding, M.; Hyttel, P. Ultrastructural visualization of the Mesenchymal-to-Epithelial Transition during reprogramming of human fibroblasts to induced pluripotent stem cells. Stem Cell Res. 2015, 14, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J.P. Mesenchymal-epithelial transition in development and reprogramming. Nat. Cell. Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef]
- Morrison, C.D.; Parvani, J.G.; Schiemann, W.P. The relevance of the TGF-β Paradox to EMT-MET programs. Cancer Lett. 2013, 341, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Alotaibi, H.; Basilicata, M.F.; Shehwana, H.; Kosowan, T.; Schreck, I.; Braeutigam, C.; Konu, O.; Brabletz, T.; Stemmler, M.P. Enhancer cooperativity as a novel mechanism underlying the transcriptional regulation of E-cadherin during mesenchymal to epithelial transition. Biochim. Et Biophys. Acta (BBA) Gene Regul. Mech. 2015, 1849, 731–742. [Google Scholar] [CrossRef]
- Ramos, C.; Becerril, C.; Montaño, M.; García-De-Alba, C.; Ramírez, R.; Checa, M.; Pardo, A.; Selman, M. FGF-1 reverts epi-thelial-mesenchymal transition induced by TGF-β1 through MAPK/ERK kinase pathway. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2010, 299, L222–L231. [Google Scholar] [CrossRef] [Green Version]
- Cisneros-Lira, J.; Gaxiola, M.; Ramos, C.; Selman, M.; Pardo, A. Cigarette smoke exposure potentiates bleomycin-induced lung fibrosis in guinea pigs. Am. J. Physiol. Cell. Mol. Physiol. 2003, 285, L949–L956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Quackenbush, J.F.; Cassidy, P.B.; Pfeffer, L.M.; Boucher, K.M.; Hawkes, J.E.; Pfeffer, S.R.; Kopelovich, L.; Leachman, S.A. Isolation of Circulating MicroRNAs from Microvesicles Found in Human Plasma. Methods Mol. Biol. 2014, 1102, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, G.; Hagood, J.S.; Sanders, Y.; Ramírez, R.; Becerril, C.; Segura, L.; Barrera, L.; Selman, M.; Pardo, A. Absence of Thy-1 results in TGF-[beta] induced MMP-9 expression and confers a profibrotic phenotype to human lung fibroblasts. Lab. Invest. 2011, 91, 1206–1218. [Google Scholar]
- Pardo, A.; Selman, M. Role of matrix metaloproteases in idiopathic pulmonary fibrosis. Fibrogenesis Tissue Repair 2012, 5, S9. [Google Scholar] [CrossRef] [Green Version]
- Tricarico, C.; Clancy, J.; D’Souza-Schorey, C. Biology and biogenesis of shed microvesicles. Small Gtpases 2017, 8, 220–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullock, M.D.; Sayan, A.E.; Packham, G.K.; Mirnezami, A.H. MicroRNAs: Critical regulators of epithelial to mesenchymal (EMT) and mesenchymal to epithelial transition (MET) in cancer progression. Biol. Cell 2011, 104, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Chaffer, C.L.; Brennan, J.P.; Slavin, J.L.; Blick, T.; Thompson, E.W.; Williams, E.D. Mesenchymal-to-Epithelial Transition Facilitates Bladder Cancer Metastasis: Role of Fibroblast Growth Factor Receptor-2. Cancer Res. 2006, 66, 11271–11278. [Google Scholar] [CrossRef] [Green Version]
- Demirkan, B. The Roles of Epithelial-to-Mesenchymal Transition (EMT) and Mesenchymal-to-Epithelial Transition (MET) in Breast Cancer Bone Metastasis: Potential Targets for Prevention and Treatment. J. Clin. Med. 2013, 2, 264–282. [Google Scholar] [CrossRef] [Green Version]
- Huleihel, L.; Ben-Yehudah, A.; Milosevic, J.; Yu, G.; Pandit, K.; Sakamoto, K.; Yousef, H.; Lejeune, M.; Coon, T.A.; Redinger, C.J.; et al. Let-7d microRNA affects mesenchymal phenotypic properties of lung fibroblasts. Am. J. Physiol. Cell. Mol. Physiol. 2014, 306, L534–L542. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Wan, X.; Geng, J.; Li, F.; Wang, C.; Dai, H. Kinase inhibitors fail to induce mesenchymal-epithelial transition in fi-broblasts from fibrotic lung tissue. Int. J. Mol. Med. 2013, 32, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.K.; Wei, Y.; Szekeres, C.; Kugler, M.C.; Wolters, P.C.; Hill, M.L.; Frank, J.A.; Brumwell, A.N.; Wheeler, S.E.; Kreidberg, J.A.; et al. Epithelial cell α3β1 integrin links β-catenin and Smad signaling to promote myofibroblast formation and pulmonary fibrosis. J. Clin. Investig. 2009, 119, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.K.; Kugler, M.C.; Wolters, P.J.; Robillard, L.; Galvez, M.G.; Brumwell, A.N.; Sheppard, D.; Chapman, H.A. Alveolar epithelial cell mesenchymal transition develops in vivo during pulmonary fibrosis and is regulated by the extracellular matrix. Proc. Natl. Acad. Sci. USA 2006, 103, 13180–13185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epitheli-al-mesenchymal transition in alveolar epithelial cells by transforming growth factor-β1: Potential role in idiopathic pulmo-nary fibrosis. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef]
- Lee, C.G.; McCarthy, S.; Gruidl, M.; Timme, C.; Yeatman, T.J. MicroRNA-147 Induces a Mesenchymal-To-Epithelial Transition (MET) and Reverses EGFR Inhibitor Resistance. PLoS ONE 2014, 9, e84597. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Lu, J.; Li, X.; Zhu, H.; Fan, X.; Zhu, S.; Wang, Y.; Guo, Q.; Wang, L.; Huang, Y.; et al. MiR-200a inhibits epithelial-mesenchymal transition of pancreatic cancer stem cell. BMC Cancer 2014, 14, 85. [Google Scholar] [CrossRef] [Green Version]
- Takaishi, M.; Tarutani, M.; Takeda, J.; Sano, S. Mesenchymal to Epithelial Transition Induced by Reprogramming Factors Attenuates the Malignancy of Cancer Cells. PLoS ONE 2016, 11, e0156904. [Google Scholar] [CrossRef] [Green Version]
- Zeisberg, M.; Shah, A.A.; Kalluri, R. Bone Morphogenic Protein-7 Induces Mesenchymal to Epithelial Transition in Adult Renal Fibroblasts and Facilitates Regeneration of Injured Kidney. J. Biol. Chem. 2005, 280, 8094–8100. [Google Scholar] [CrossRef] [Green Version]
- Hamazaki, Y.; Itoh, M.; Sasaki, H.; Furuse, M.; Tsukita, S. Multi-PDZ Domain Protein 1 (MUPP1) Is Concentrated at Tight Junctions through Its Possible Interaction with Claudin-1 and Junctional Adhesion Molecule. J. Biol. Chem. 2002, 277, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Selman, M.; Pardo, A. Idiopathic pulmonary fibrosis: An epithelial/fibroblastic cross-talk disorder. Respir. Res. 2002, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Behrens, P.; Rothe, M.; Wellmann, A.; Krischler, J.; Wernert, N. The Ets-1 transcription factor is up-regulated together with MMP 1 and MMP 9 in the stroma of pre-invasive breast cancer. J. Pathol. 2001, 194, 43–50. [Google Scholar] [CrossRef]
- Suzuki, M.; Hirao, A.; Mizuno, A. Microtubule-associated protein 7 increases the membrane expression of transient receptor potential vanilloid 4 (TRPV4). Vol. 278 (2003)51448–51453. J. Biol. Chem. 2005, 280, 25948. [Google Scholar] [CrossRef]
- Acharya, P.S.; Zukas, A.; Chandan, V.; Katzenstein, A.-L.A.; Puré, E. Fibroblast activation protein: A serine protease expressed at the remodeling interface in idiopathic pulmonary fibrosis. Hum. Pathol. 2006, 37, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Veronese, A.; Pichiorri, F.; Lee, T.J.; Jeon, Y.-J.; Volinia, S.; Pineau, P.; Marchio, A.; Palatini, J.; Suh, S.-S.; et al. p53 regulates epithelial–mesenchymal transition through microRNAs targeting ZEB1 and ZEB2. J. Exp. Med. 2011, 208, 875–883. [Google Scholar] [CrossRef]
- Ru, P.; Steele, R.; Newhall, P.; Phillips, N.J.; Toth, K.; Ray, R.B. miRNA-29b suppresses prostate cancer metastasis by reg-ulating epithelial–mesenchymal transition signaling. Mol. Cancer. Ther. 2012, 11, 1166–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicchini, C.; De Nonno, V.; Battistelli, C.; Cozzolino, A.M.; Puzzonia, M.D.S.; Ciafrè, S.A.; Brocker, C.; Gonzalez, F.J.; Amicone, L.; Tripodi, M. Epigenetic control of EMT/MET dynamics: HNF4α impacts DNMT3s through miRs-29. Biochim. Et Biophys. Acta (BBA) Bioenergy 2015, 1849, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-H.; Hsu, W.-L.; Tseng, Y.-J.; Liu, D.-W.; Weng, C.-F. Involvement of DNMT 3B promotes epithelial-mesenchymal transition and gene expression profile of invasive head and neck squamous cell carcinomas cell lines. BMC Cancer 2016, 16, 431. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhao, S.; Li, H.; Chang, H.; Huang, Z.; Ding, Z.; Dong, L.; Chen, J.; Zang, Y.; Zhang, J. MicroRNA-30b Suppresses Epithelial-Mesenchymal Transition and Metastasis of Hepatoma Cells. J. Cell. Physiol. 2016, 232, 625–634. [Google Scholar] [CrossRef]
- Liu, W.; Li, H.; Wang, Y.; Zhao, X.; Guo, Y.; Jin, J.; Chi, R. MiR-30b-5p functions as a tumor suppressor in cell proliferation, me-tastasis and epithelial-to-mesenchymal transition by targeting G-protein subunit α-13 in renal cell carcinoma. Gene 2017, 26, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, P.; Yang, R.; Cheng, R.; Zhang, F.; Wang, Y.; Chen, X.; Sun, Q.; Zang, W.; Du, Y.; et al. microRNA-30b inhibits cell invasion and migration through targeting collagen triple helix repeat containing 1 in non-small cell lung cancer. Cancer Cell Int. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Li, Y.; He, T.; Hu, J.; Liu, J.; Chen, M.; Shi, M.; Jiang, Z.; Yang, S.; Mao, X.; et al. Identification of miR-30b as an oncogene in renal cell carcinoma. Mol. Med. Rep. 2017, 15, 1837–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, H.; Liu, J.; Tu, X.; Zang, Y.; Zhu, J.; Chen, J.; Dong, L.; Zhang, J. miR-30 inhibits TGF-β1-induced epithe-lial-to-mesenchymal transition in hepatocyte by targeting Snail1. Biochem. Biophys. Res. Commun. 2012, 417, 100–1105. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change | Symbol | Entrez Gene Name | Fold Change | Symbol | Entrez Gene Name |
|---|---|---|---|---|---|
| 8.39 | AKR1B10 | aldo-keto reductase family 1, member B10 | 4.83 | UPK1B | uroplakin 1B |
| 7.74 | SLCO1B3 | solute carrier organic anion transporter family member 1B3 | 4.83 | ST6GAL2 | ST6 beta-galactoside alpha-2,6-sialyltransferase 2 |
| 7.72 | CYP24A1 | cytochrome P450 family 24 subfamily A member 1 | 4.79 | TIMD4 | T cell immunoglobulin and mucin domain containing 4 |
| 7.5 | GPX2 | glutathione peroxidase 2 | 4.77 | TM4SF20 | transmembrane 4 L six family member 20 |
| 7.37 | CEACAM6 | CEA cell adhesion molecule 6 | 4.77 | CXADR | CXADR Ig-like cell adhesion molecule |
| 7.07 | KYNU | kynureninase | 4.76 | SMOC1 | SPARC related modular calcium binding 1 |
| 6.97 | PON3 | paraoxonase 3 | 4.71 | NR5A2 | nuclear receptor subfamily 5 group A member 2 |
| 6.76 | SPP1 | secreted phosphoprotein 1 | 4.67 | ANKRD18A | ankyrin repeat domain 18A |
| 6.64 | ABCC2 | ATP binding cassette subfamily C member 2 | 4.66 | CST1 | cystatin SN |
| 6.61 | CP | ceruloplasmin | 4.62 | TBC1D8 | TBC1 domain family member 8 |
| 6.55 | HAVCR1 | hepatitis A virus cellular receptor 1 | 4.59 | ACSM3 | acyl-CoA synthetase medium chain family member 3 |
| 6.44 | SLC27A2 | solute carrier family 27 member 2 | 4.56 | CLDN1 | claudin 1 |
| 6.44 | TNS4 | tensin 4 | 4.5 | ANO5 | anoctamin 5 |
| 6.42 | CNTN1 | contactin 1 | 4.49 | PLS1 | plastin 1 |
| 6.3 | HORMAD1 | HORMA domain containing 1 | 4.49 | ITGB6 | integrin subunit beta 6 |
| 6.2 | LGSN | lengsin, lens protein with glutamine synthetase domain | 4.48 | HNF1B | HNF1 homeobox B |
| 6.17 | HPGD | 15-hydroxyprostaglandin dehydrogenase | 4.46 | ARHGAP26 | Rho GTPase activating protein 26 |
| 6.08 | CEACAM7 | CEA cell adhesion molecule 7 | 4.46 | AKR1C1 | aldo-keto reductase family 1 member C1 |
| 6.07 | KRT81 | keratin 81 | 4.41 | ELOVL7 | ELOVL fatty acid elongase 7 |
| 6.06 | HGD | homogentisate 1,2-dioxygenase | 4.41 | SLPI | secretory leukocyte peptidase inhibitor |
| 6.06 | AKR1C3 | aldo-keto reductase family 1, member C3 | 4.41 | F5 | coagulation factor V |
| 5.87 | NTRK3 | neurotrophic receptor tyrosine kinase 3 | 4.37 | EHF | ETS homologous factor |
| 5.8 | AREG | anphiregulin | 4.34 | MLPH | melanophilin |
| 5.8 | KRTS | keratin 8 | 4.31 | HIST1H2AB | H2A clustered histone 4 |
| 5.75 | MMP7 | matrix metallopeptidase 7 | 4.31 | ELF3 | E74 like ETS transcription factor 3 |
| 5.7 | GCNT3 | glucosaminyl (N-acetyl) transferase 3, mucin type | 4.3 | CEACAM5 | CEA cell adhesion molecule 5 |
| 5.67 | AKR1C2 | aldo-keto reductase family 1, member C2 | 4.25 | WDR72 | WD repeat domain 72 |
| 5.66 | TM4SF18 | transmembrane 4 L six family member 18 | 4.23 | ITGB4 | integrin subunit beta 4 |
| 5.66 | SLC22A3 | solute carrier family 22 member 3 | 4.23 | MAP7 | microtubule associated protein 7 |
| 5.63 | CDH1 | cadherin 1 | 4.22 | FGB | fibrinogen beta chain |
| 5.52 | CDH17 | cadherin 17 | 4.21 | TESC | tescalcin |
| 5.46 | PPARGC1A | PPARG coactivator 1 alpha | 4.19 | NRCAM | neuronal cell adhesion molecule |
| 5.41 | HNF1A-AS1 | HNF1A antisense RNA 1 | 4.16 | IGFBP1 | insulin like growth factor binding protein 1 |
| 5.36 | GPRIN3 | GPRIN family member 3 | 4.15 | OAS1 | 2’-5’-oligoadenylate synthetase 1 |
| 5.31 | GABRB3 | gamma-aminobutyric acid type A receptor subunit beta3 | 4.11 | CDRT1 | CMT1A duplicated region transcript 1 |
| 5.31 | EREG | epiregulin | 4.09 | TFAP2A | transcription factor AP-2 alpha |
| 5.31 | TSPAN7 | tetraspanin 7 | 4.06 | TC2N | tandem C2 domains, nuclear |
| 5.29 | ARSE | arylsulfatase E | 4.06 | POF1B | POF1B actin binding protein |
| 5.2 | RAB27B | RAB27B, member RAS oncogene family | 4.04 | mir622 | microRNA 622 |
| 5.16 | SLC17A3 | solute carrier family 17 member 3 | 4.01 | MTUS1 | microtubule associated scaffold protein 1 |
| 5.13 | CYP4F11 | cytochrome P450 family 4 subfamily F member 11 | 3.99 | CPS1 | carbamoyl-phosphate synthase 1 |
| 5.13 | SLCO1B7 | solute carrier organic anion transporter family member 1B7 (putative) | 3.99 | FILIP1 | filamin A interacting protein 1 |
| 5.11 | RSPO3 | R-spondin 3 | 3.98 | CYP4F3 | cytochrome P450 family 4 subfamily F member 3 |
| 5.01 | FGL1 | fibrinogen-like 1 | 3.98 | FZD3 | frizzled class receptor 3 |
| 5 | AREG | amphiregulin | 3.98 | CHRNA5 | cholinergic receptor nicotinic alpha 5 subunit |
| 4.99 | ALDH3A1 | aldehyde dehydrogenase 3 family member A1 | 3.97 | KRT18 | keratin 18 |
| 4.98 | MUC13 | mucin 13, cell surface associated | 3.96 | KCNJ16 | potassium inwardly rectifying channel subfamily J member 16 |
| 4.86 | CLMN | calmin | 3.93 | GABRA5 | gamma-aminobutyric acid type A receptor subunit alpha5 |
| 4.85 | CA12 | carbonic anhydrase 12 | 3.93 | GRB14 | growth factor receptor bound protein 14 |
| 4.84 | TMEM156 | transmembrane protein 156 | 3.91 | INSL4 | insulin like 4 |
| Fold Change | Symbol | Entrez Gene Name | Fold Change | Symbol | Entrez Gene Name |
|---|---|---|---|---|---|
| −8.32 | SPARC | secreted protein acidic and cysteine rich | −4.93 | JAM2 | junctional adhesion molecule 2 |
| −7.5 | SERPINB2 | serpin family B member 2 | −4.92 | FLNC | filamin C |
| −7.3 | POSTN | periostin | −4.92 | COL3A1 | collagen type III alpha 1 chain |
| −7.24 | LOX | lysyl oxidase | −4.92 | FMOD | fibromodulin |
| −7.15 | DCN | decorin | −4.9 | XYLT1 | xylosyltransferase 1 |
| −6.95 | SULF1 | sulfatase 1 | −4.88 | ENG | endoglin |
| −6.95 | IGFBP5 | insulin like growth factor binding protein 5 | −4.87 | PCDH18 | protocadherin 18 |
| −6.88 | LUM | lumican | −4.87 | MAN1A1 | mannosidase alpha class 1A member 1 |
| −6.82 | TIMP3 | TIMP metallopeptidase inhibitor 3 | −4.84 | TRPS1 | transcriptional repressor GATA binding 1 |
| −6.68 | FAP | fibroblast activation protein alpha | −4.83 | ZNF737 | zinc finger protein 737 |
| −6.51 | COL6A3 | collagen type VI alpha 3 chain | −4.83 | EGR1 | early growth response 1 |
| −6.5 | LAMA4 | laminin subunit alpha 4 | −4.81 | MIR145 | microRNA 145 |
| −6.46 | CDH11 | cadherin 11 | −4.81 | ARID5B | AT-rich interaction domain 5B |
| −6.23 | ZNF253 | zinc finger protein 253 | −4.8 | FAM26E | calcium homeostasis modulator family member 5 |
| −6.14 | THBS2 | thrombospondin 2 | −4.77 | HAS2 | hyaluronan synthase 2 |
| −6.11 | BGN | biglycan | −4.76 | IRAK3 | interleukin 1 receptor associated kinase 3 |
| −6.09 | CDH13 | cadherin 13 | −4.76 | LHFPL6 | LHFPL tetraspan subfamily member 6 |
| −6.01 | INHBA | inhibin subunit beta A | −4.7 | PDCD1LG2 | programmed cell death 1 ligand 2 |
| −5.96 | COL1A2 | collagen type I alpha 2 chain | −4.69 | TNFRSF11B | TNF receptor superfamily member 11b |
| −5.94 | MMP2 | matrix metallopeptidase 2 | −4.68 | ZNF83 | zinc finger protein 83 |
| −5.92 | SLIT2 | slit guidance ligand 2 | −4.67 | MMP14 | matrix metallopeptidase 14 |
| −5.9 | SNAI2 | snail family transcriptional repressor 2 | −4.67 | NDNF | neuron derived neurotrophic factor |
| −5.87 | FGF7 | fibroblast growth factor 7 | −4.65 | TNC | tenascin C |
| −5.76 | LY96 | lymphocyte antigen 96 | −4.63 | ANTXR2 | ANTXR cell adhesion molecule 2 |
| −5.76 | DPP4 | dipeptidyl peptidase 4 | −4.6 | FBN1 | fibrillin 1 |
| −5.64 | DDR2 | discoidin domain receptor tyrosine kinase 2 | −4.57 | PTGS1 | prostaglandin-endoperoxide synthase 1 |
| −5.61 | SERPINF1 | serpin family F member 1 | −4.55 | CRYAB | crystallin alpha B |
| −5.54 | EDIL3 | EGF like repeats and discoidin domains 3 | −4.53 | ITGB3 | integrin subunit beta 3 |
| −5.46 | GREM1 | gremlin 1, DAN family BMP antagonist | −4.51 | DSEL | dermatan sulfate epimerase like |
| −5.45 | ELTD1 | adhesion G protein-coupled receptor L4 | −4.5 | PDGFRB | platelet derived growth factor receptor beta |
| −5.43 | SERPING1 | serpin family G member 1 | −4.5 | EFEMP2 | EGF containing fibulin extracellular matrix protein 2 |
| −5.4 | DDR2 | discoidin domain receptor tyrosine kinase 2 | −4.50 | MMP3 | matrix metallopeptidase 3 |
| −5.4 | SLFN11 | schlafen family member 11 | −4.47 | KGFLP1 | fibroblast growth factor 7 pseudogene 6 |
| −5.38 | IFITM3 | interferon induced transmembrane protein 3 | −4.46 | LOC100288114 | Uncharacterized |
| −5.36 | DOK5 | docking protein 5 | −4.46 | ANGPTL2 | angiopoietin like 2 |
| −5.34 | CDK14 | cyclin dependent kinase 14 | −4.45 | IFITM1 | interferon induced transmembrane protein 1 |
| −5.31 | IL6 | interleukin 6 | −4.45 | ZFPM2 | zinc finger protein, FOG family member 2 |
| −5.29 | PDGFRA | platelet derived growth factor receptor alpha | −4.44 | ENPP2 | ectonucleotide pyrophosphatase/phosphodiesterase 2 |
| −5.24 | GLT8D2 | glycosyltransferase 8 domain containing 2 | −4.43 | EDNRA | endothelin receptor type A |
| −5.23 | PTGIS | prostaglandin I2 synthase | −4.41 | OLFML3 | olfactomedin like 3 |
| −5.21 | KIAA1199 | cell migration inducing hyaluronidase 1 | −4.37 | ADAMTS6 | ADAM metallopeptidase with thrombospondin type 1 motif 6 |
| −5.18 | PRRX1 | paired related homeobox 1 | −4.36 | EPB41L3 | erythrocyte membrane protein band 4.1 like 3 |
| −5.14 | WNT5A | Wnt family member 5A | −4.34 | PDE5A | phosphodiesterase 5A |
| −5.1 | PXDN | peroxidasin | −4.31 | MFAP4 | microfibril associated protein 4 |
| −5.06 | FLRT2 | fibronectin leucine rich transmembrane protein 2 | −4.31 | CRISPLD2 | cysteine rich secretory protein LCCL domain containing 2 |
| −5.06 | CDH6 | cadherin 6 | −4.3 | NEXN | nexilin F-actin binding protein |
| −5.01 | NID1 | nidogen 1 | −4.29 | SNORD113-4 | small nucleolar RNA, C/D box 113-4 |
| −4.97 | PAMR1 | peptidase domain containing associated with muscle regeneration 1 | −4.28 | DNAJC15 | DnaJ heat shock protein family (Hsp40) member C15 |
| −4.97 | MME | membrane metalloendopeptidase | −4.24 | ZEB2 | ZEB2 antisense RNA 1 |
| −4.94 | COL1A1 | collagen type I alpha 1 chain | −4.23 | FN1 | fibronectin 1 |
| F vs. ELC | FC |
|---|---|
| hsa-let-7a-5p | 3.1 |
| hsa-let-7b-5p | 66.5 |
| hsa-let-7c | 4.4 |
| hsa-let-7d-5p | 19.7 |
| hsa-let-7e-5p | 5.2 |
| hsa-miR-21-3p | 8.1 |
| hsa-miR-23a-3p | 3.0 |
| hsa-miR-23b-3p | 4.9 |
| hsa-miR-24-2-5p | 2.2 |
| hsa-miR-26a-5p | 49.4 |
| hsa-miR-27a-3p | 16.8 |
| ELC vs. F | |
| hsa-miR-30b-5p | 2.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becerril, C.; Montaño, M.; Cisneros, J.; Mendoza-Milla, C.; Pardo, A.; Ortiz-Quintero, B.; Selman, M.; Ramos, C. Mesenchymal–Epithelial Transition in Fibroblasts of Human Normal Lungs and Interstitial Lung Diseases. Biomolecules 2021, 11, 378. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030378
Becerril C, Montaño M, Cisneros J, Mendoza-Milla C, Pardo A, Ortiz-Quintero B, Selman M, Ramos C. Mesenchymal–Epithelial Transition in Fibroblasts of Human Normal Lungs and Interstitial Lung Diseases. Biomolecules. 2021; 11(3):378. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030378
Chicago/Turabian StyleBecerril, Carina, Martha Montaño, José Cisneros, Criselda Mendoza-Milla, Annie Pardo, Blanca Ortiz-Quintero, Moisés Selman, and Carlos Ramos. 2021. "Mesenchymal–Epithelial Transition in Fibroblasts of Human Normal Lungs and Interstitial Lung Diseases" Biomolecules 11, no. 3: 378. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11030378