
Paternal High-Protein Diet Programs Offspring Insulin Sensitivity in a Sex-Specific Manner

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets
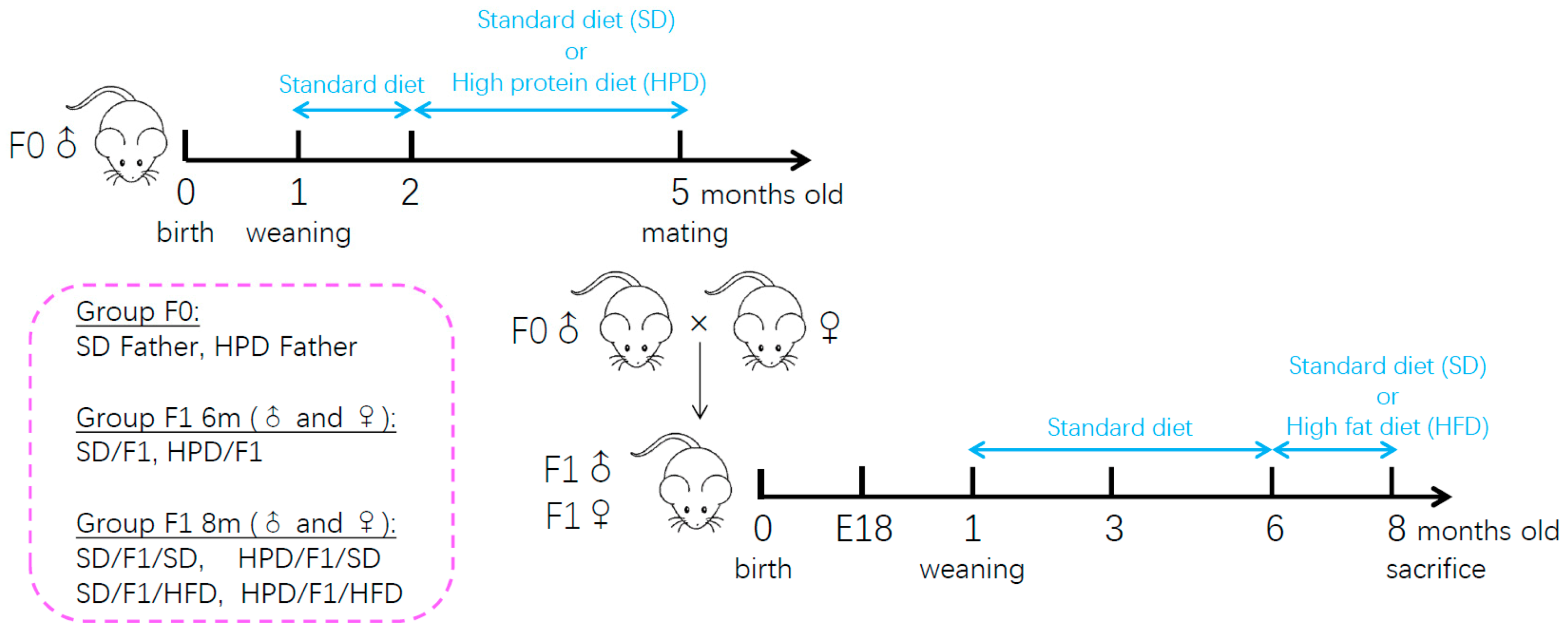
2.3. Experimental Design
2.4. Body Composition
2.5. Insulin Tolerance Test
2.6. Glucose Tolerance Test
2.7. Quantitative Real-Time PCR
2.8. Histological Studies
2.9. Analysis of Islet Fibrosis
2.10. Sperm Collection
2.11. Small RNA-Sequencing Analysis
2.12. Data and Statistical Analysis
3. Results
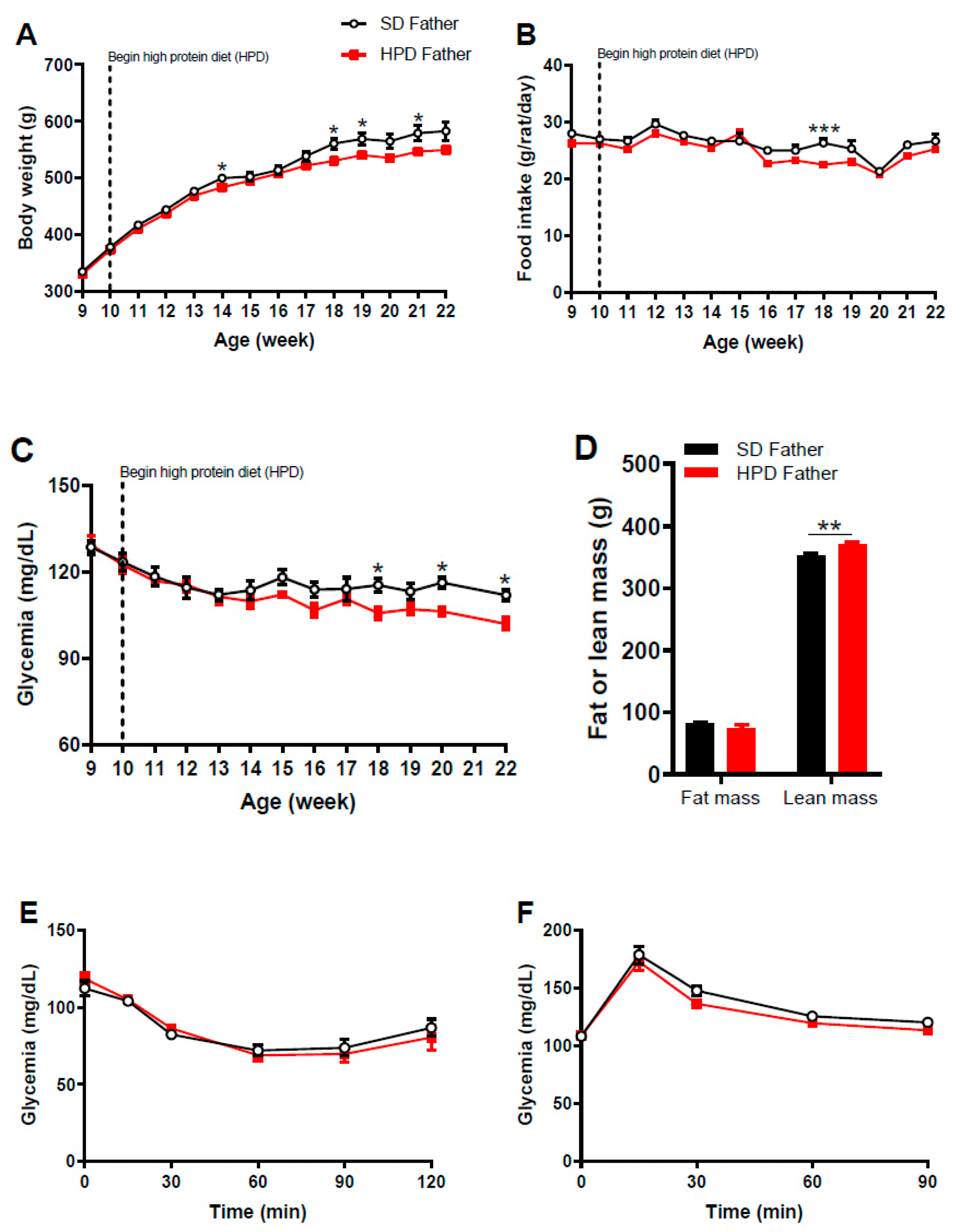
3.1. Phenotype of Fathers
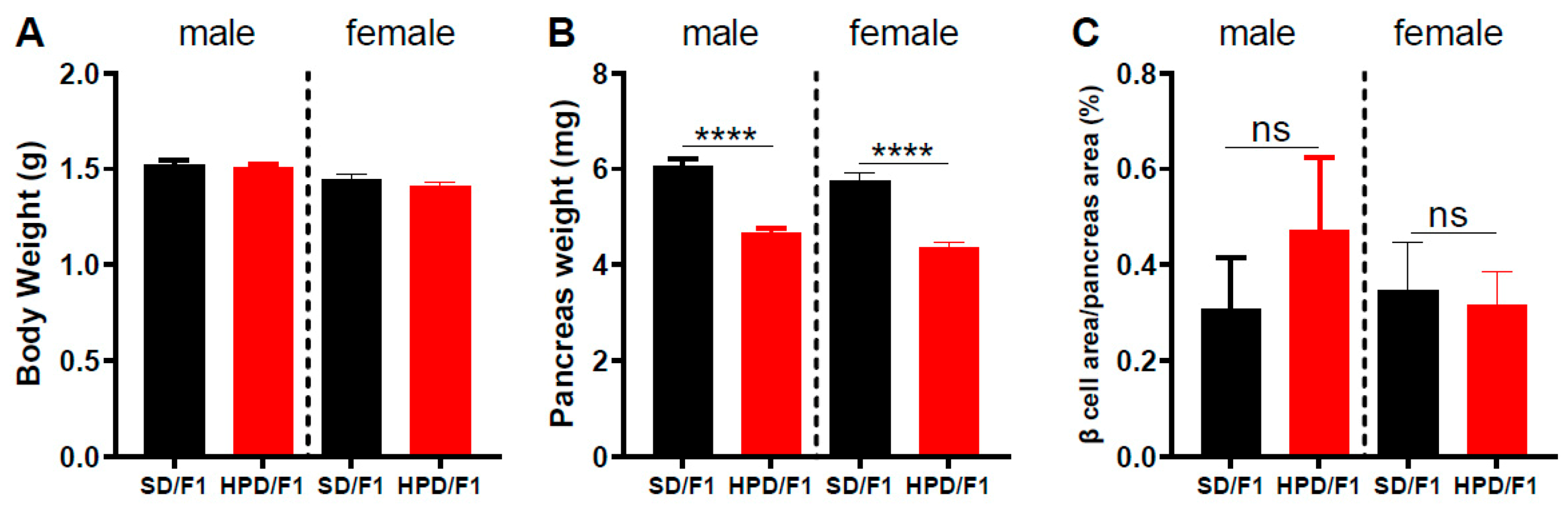
3.2. Phenotype of F1 Fetuses
3.3. Phenotype of Adult F1
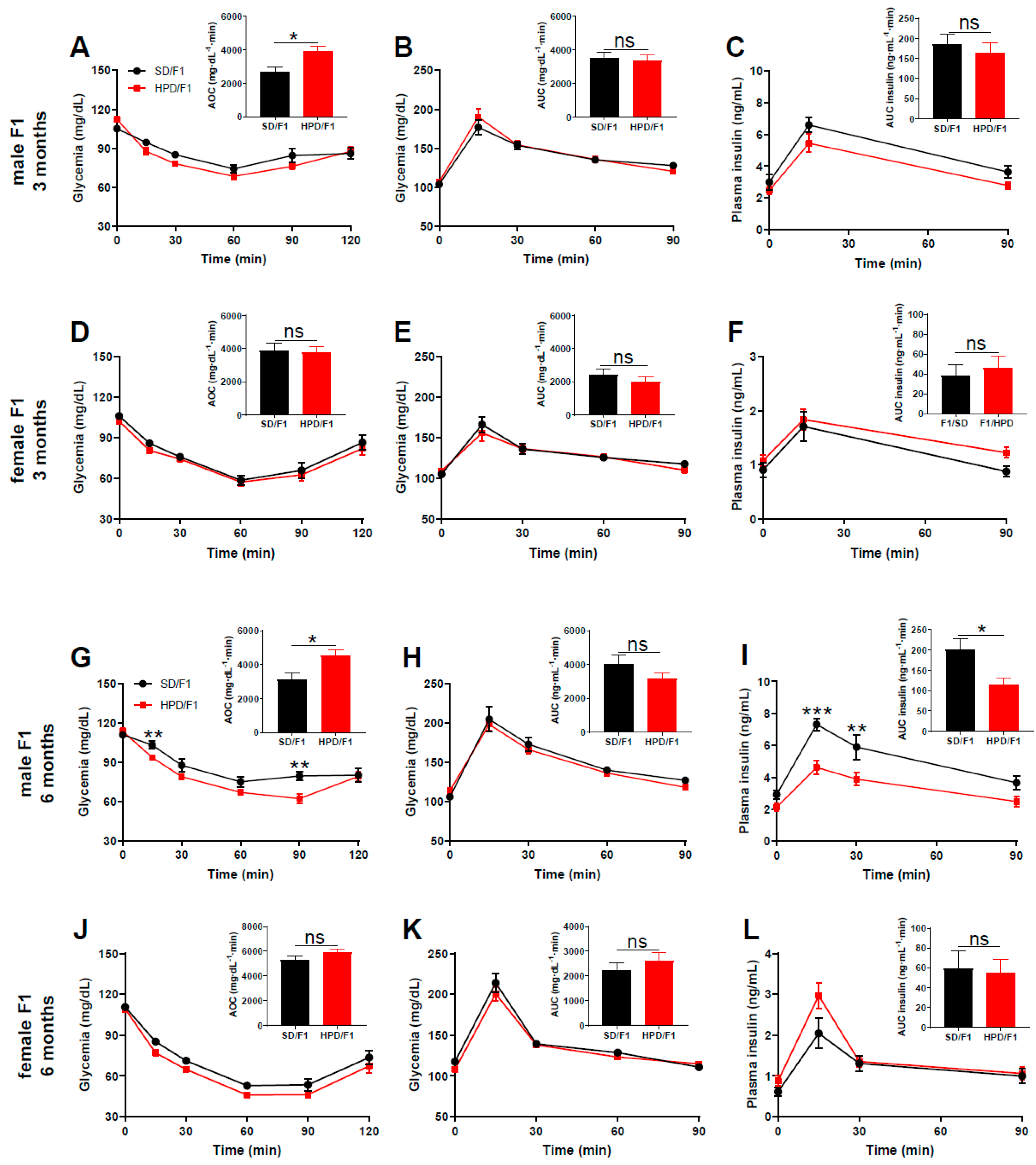
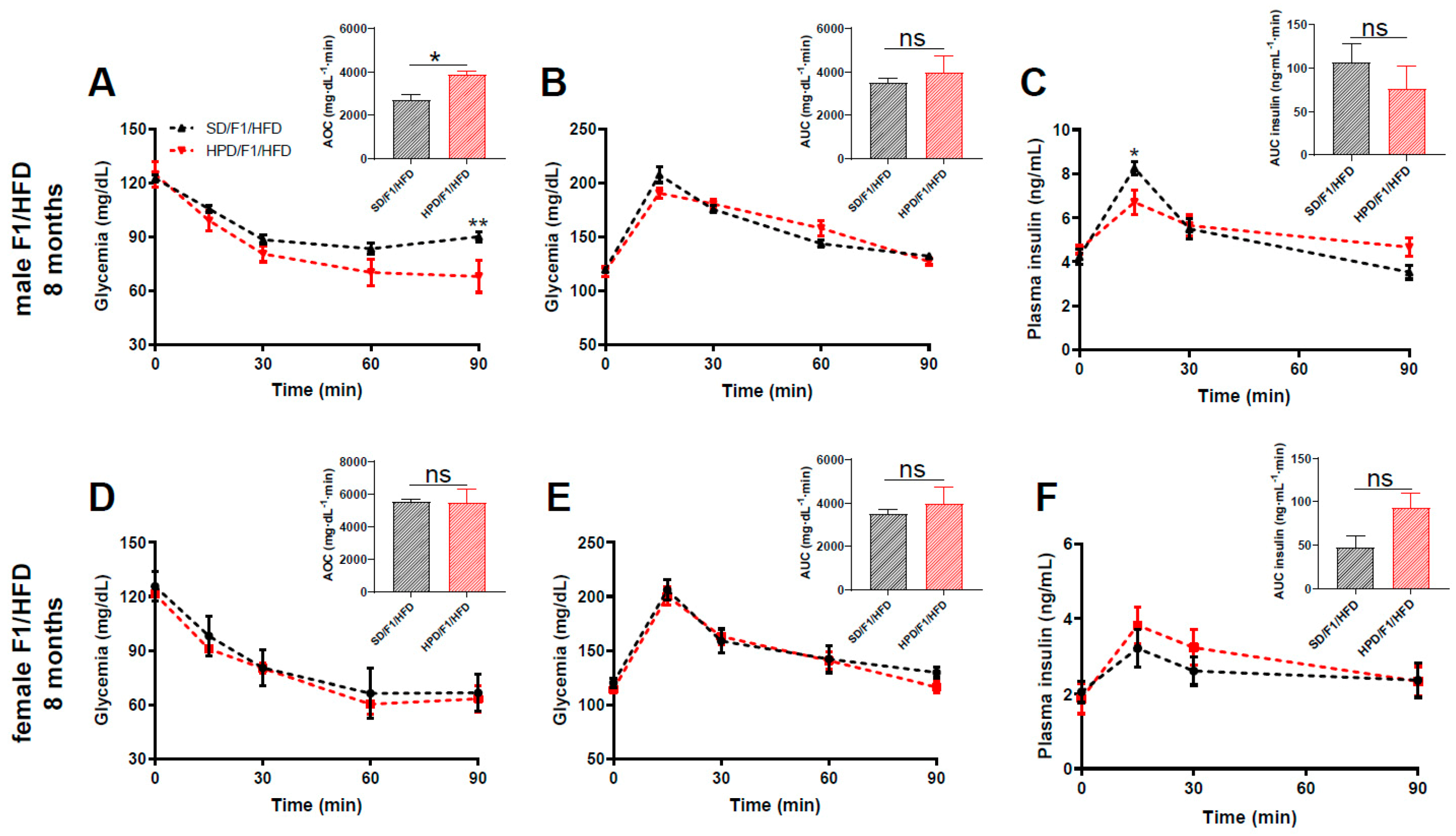
3.4. Phenotype of Adult F1 under Metabolic Challenge
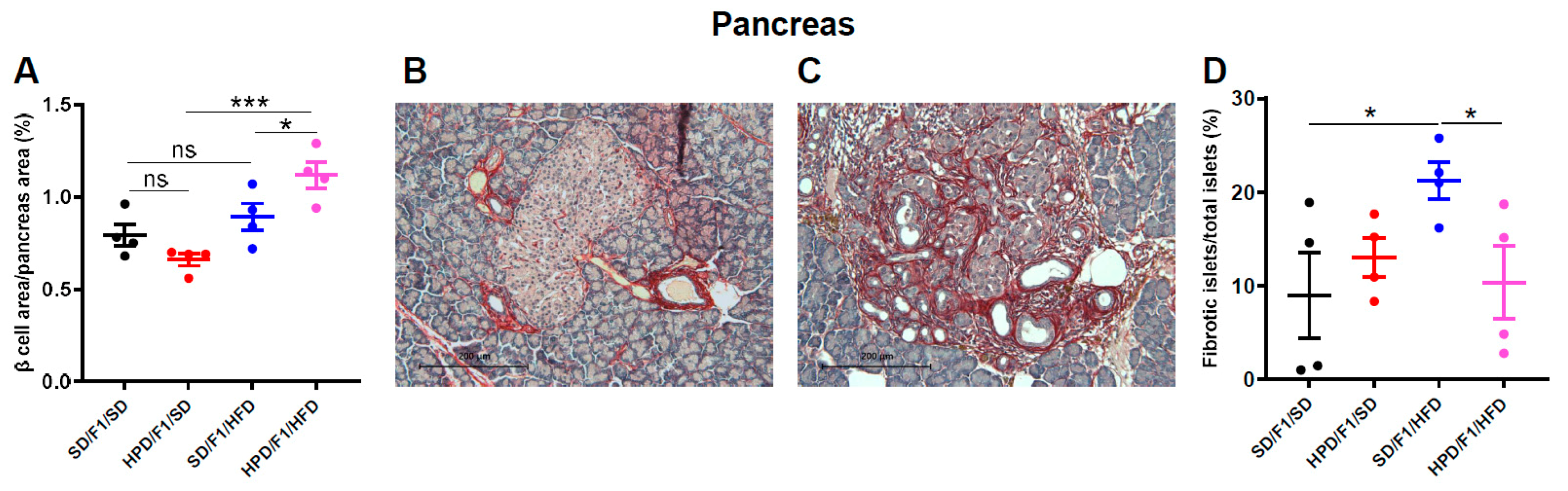
3.5. Pancreatic Parameters
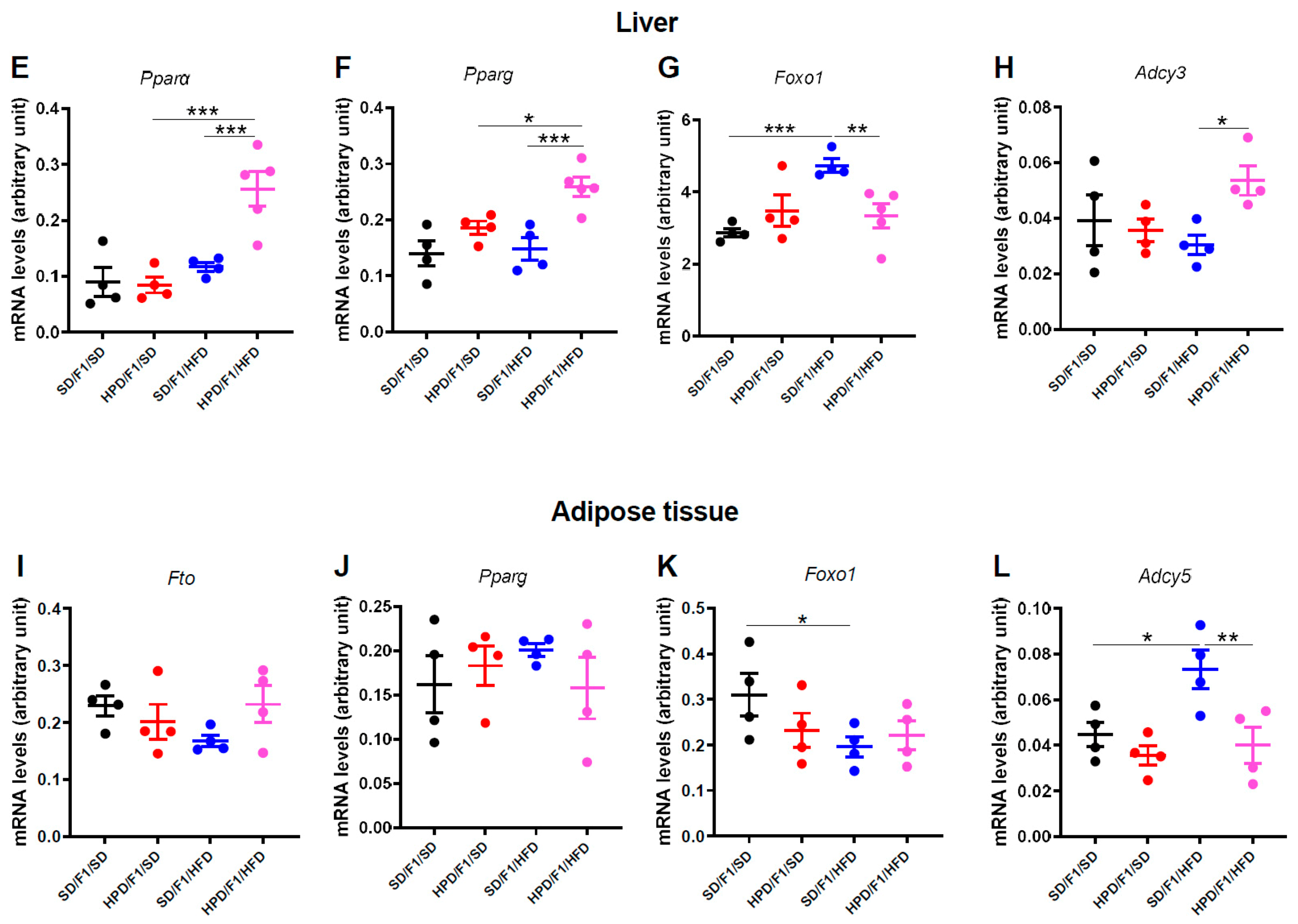
3.6. Gene Expression Studies
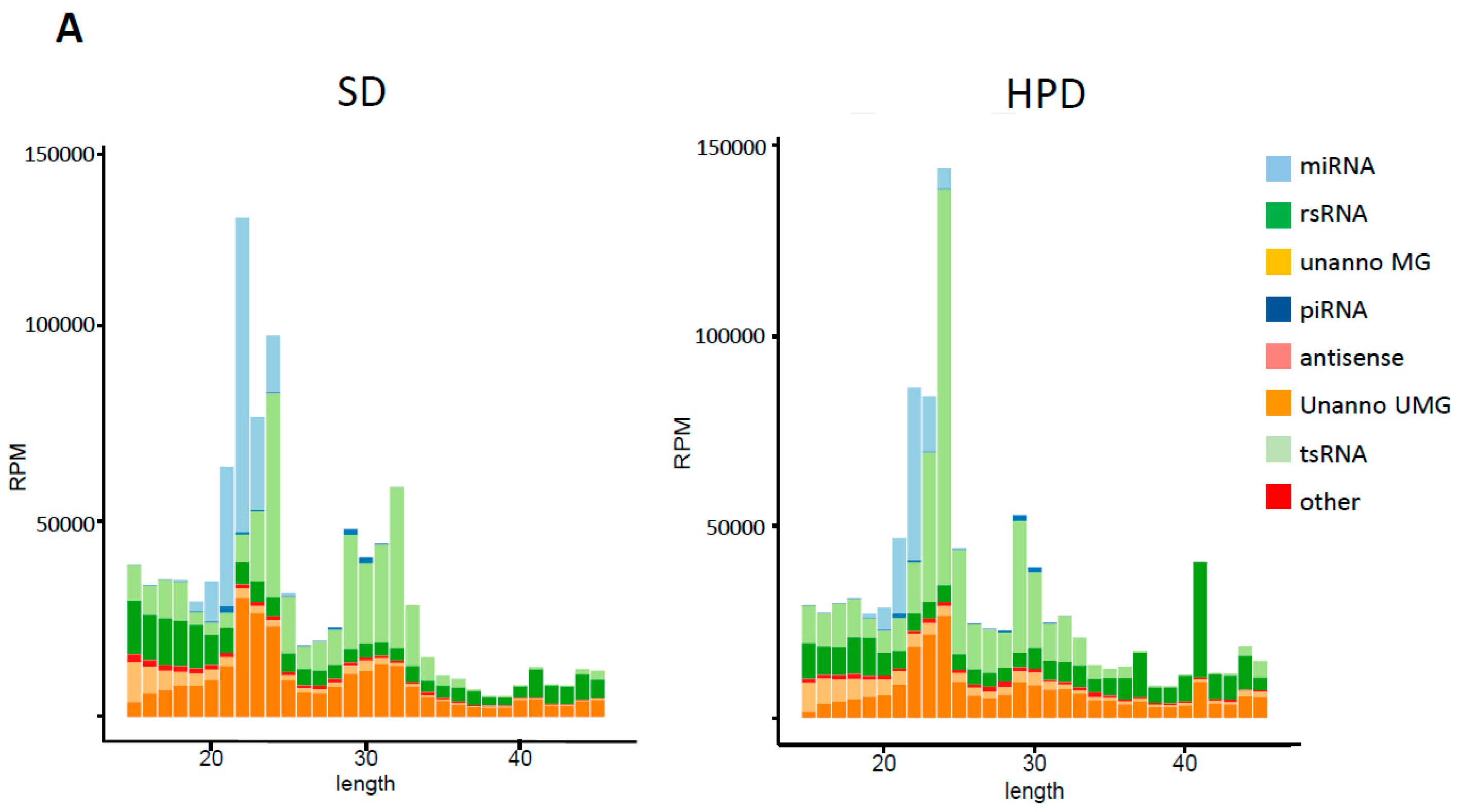
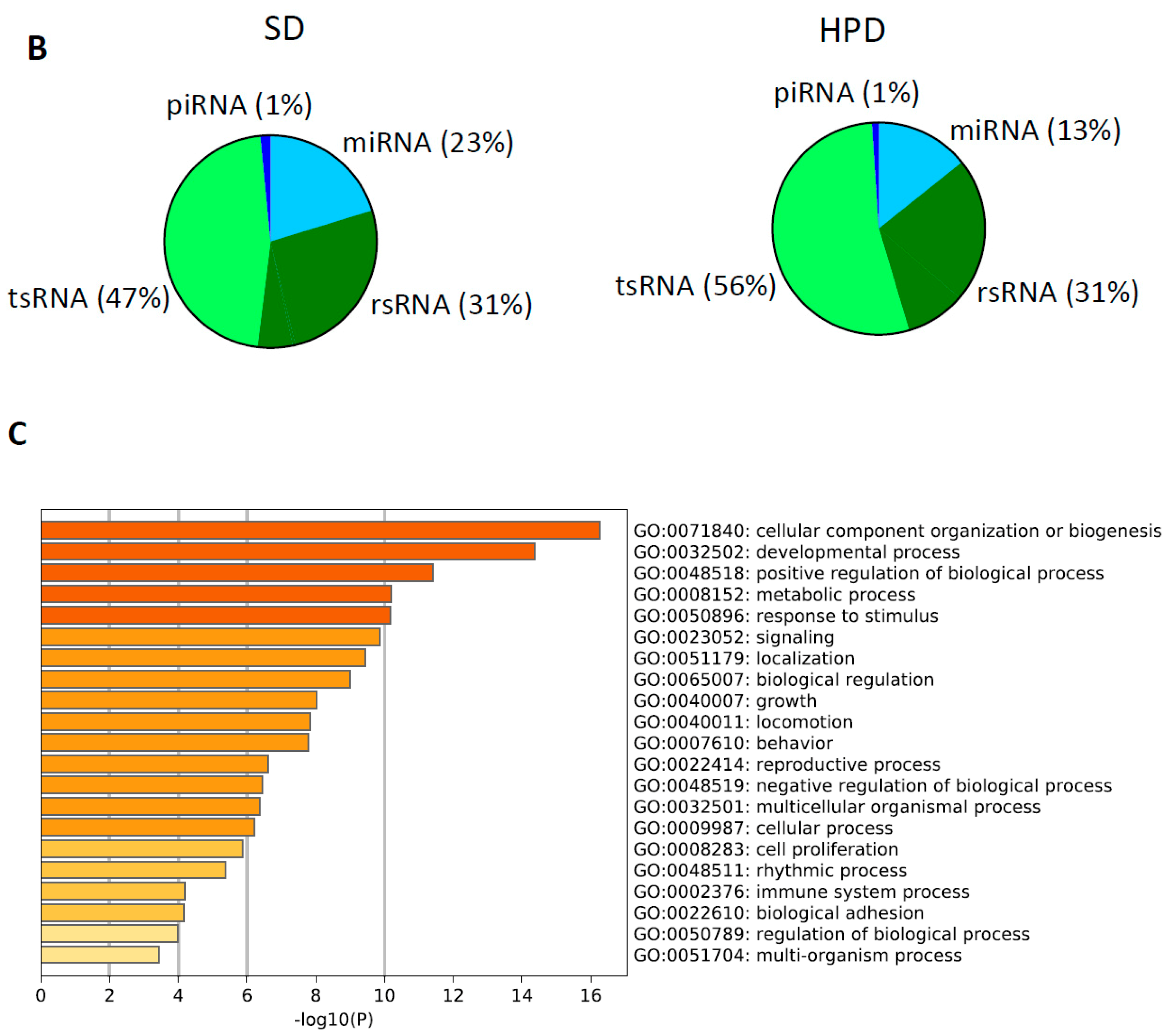
3.7. Spermatic Small RNA-Sequencing Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Barker, D. Developmental Origins of Adult Health and Disease. J. Epidemiol. Community Health 2004, 58, 114–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J.P. The Origins of the Developmental Origins Theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Soubry, A. POHaD: Why We Should Study Future Fathers. Environ. Epigenet. 2018, 4. [Google Scholar] [CrossRef]
- Dunford, A.R.; Sangster, J.M. Maternal and Paternal Periconceptional Nutrition as an Indicator of Offspring Metabolic Syndrome Risk in Later Life through Epigenetic Imprinting: A Systematic Review. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, S655–S662. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.-F.; Lin, R.C.Y.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic High-Fat Diet in Fathers Programs β-Cell Dysfunction in Female Rat Offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef] [PubMed]
- de Castro Barbosa, T.; Ingerslev, L.R.; Alm, P.S.; Versteyhe, S.; Massart, J.; Rasmussen, M.; Donkin, I.; Sjögren, R.; Mudry, J.M.; Vetterli, L.; et al. High-Fat Diet Reprograms the Epigenome of Rat Spermatozoa and Transgenerationally Affects Metabolism of the Offspring. Mol. Metab. 2015, 5, 184–197. [Google Scholar] [CrossRef]
- Wei, Y.; Yang, C.-R.; Wei, Y.-P.; Zhao, Z.-A.; Hou, Y.; Schatten, H.; Sun, Q.-Y. Paternally Induced Transgenerational Inheritance of Susceptibility to Diabetes in Mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 1873–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, H.L.; Ampong, I.; Eid, N.; Rouillon, C.; Griffiths, H.R.; Watkins, A.J. Low Protein Diet and Methyl-Donor Supplements Modify Testicular Physiology in Mice. Reproduction 2020, 159, 627–641. [Google Scholar] [CrossRef] [Green Version]
- Morgan, H.L.; Aljumah, A.; Rouillon, C.; Watkins, A.J. Paternal Low Protein Diet and the Supplementation of Methyl-Donors Impact Fetal Growth and Placental Development in Mice. Placenta 2021, 103, 124–133. [Google Scholar] [CrossRef]
- Westerterp-Plantenga, M.S.; Nieuwenhuizen, A.; Tomé, D.; Soenen, S.; Westerterp, K.R. Dietary Protein, Weight Loss, and Weight Maintenance. Annu. Rev. Nutr. 2009, 29, 21–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cintineo, H.P.; Arent, M.A.; Antonio, J.; Arent, S.M. Effects of Protein Supplementation on Performance and Recovery in Resistance and Endurance Training. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.W.; Murphy, K.T.; McKellar, S.R.; Schoenfeld, B.J.; Henselmans, M.; Helms, E.; Aragon, A.A.; Devries, M.C.; Banfield, L.; Krieger, J.W.; et al. A Systematic Review, Meta-Analysis and Meta-Regression of the Effect of Protein Supplementation on Resistance Training-Induced Gains in Muscle Mass and Strength in Healthy Adults. Br. J. Sports Med. 2018, 52, 376–384. [Google Scholar] [CrossRef]
- Farnsworth, E.; Luscombe, N.D.; Noakes, M.; Wittert, G.; Argyiou, E.; Clifton, P.M. Effect of a High-Protein, Energy-Restricted Diet on Body Composition, Glycemic Control, and Lipid Concentrations in Overweight and Obese Hyperinsulinemic Men and Women. Am. J. Clin. Nutr. 2003, 78, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Gannon, M.C.; Nuttall, F.Q.; Saeed, A.; Jordan, K.; Hoover, H. An Increase in Dietary Protein Improves the Blood Glucose Response in Persons with Type 2 Diabetes. Am. J. Clin. Nutr. 2003, 78, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Gannon, M.C.; Nuttall, F.Q. Effect of a High-Protein, Low-Carbohydrate Diet on Blood Glucose Control in People With Type 2 Diabetes. Diabetes 2004, 53, 2375–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasem, R.J.; Cherala, G.; D’mello, A.P. Maternal Protein Restriction during Pregnancy and Lactation in Rats Imprints Long-Term Reduction in Hepatic Lipid Content Selectively in the Male Offspring. Nutr. Res. 2010, 30, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Sosa-Larios, T.C.; Miliar-Garcia, A.A.; Reyes-Castro, L.A.; Morimoto, S.; Jaramillo-Flores, M.E. Retracted Article: Alterations in Lipid Metabolism Due to a Protein-Restricted Diet in Rats during Gestation and/or Lactation. Food Funct. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xiao, X.; Zhang, Q.; Wang, T.; Yu, M.; Xu, J. Maternal Low-Protein Diet Modulates Glucose Metabolism and Hepatic MicroRNAs Expression in the Early Life of Offspring. Nutrients 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Chamson-Reig, A.; Thyssen, S.M.; Hill, D.J.; Arany, E. Exposure of the Pregnant Rat to Low Protein Diet Causes Impaired Glucose Homeostasis in the Young Adult Offspring by Different Mechanisms in Males and Females. Exp. Biol. Med. 2009, 234, 1425–1436. [Google Scholar] [CrossRef]
- Snoeck, A.; Remacle, C.; Reusens, B.; Hoet, J.J. Effect of a Low Protein Diet during Pregnancy on the Fetal Rat Endocrine Pancreas. NEO 1990, 57, 107–118. [Google Scholar] [CrossRef]
- Bertin, E.; Gangnerau, M.-N.; Bellon, G.; Bailbé, D.; De Arbelot Vacqueur, A.; Portha, B. Development of β-Cell Mass in Fetuses of Rats Deprived of Protein and/or Energy in Last Trimester of Pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R623–R630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Desclée Maredsous, C.; Carlin, G.; Oosting, A.; Delteil, C.; Azzout-Marniche, D.; Chaumontet, C.; Blachier, F.; Barbillon, P.; Mary-Huard, T.; Tomé, D.; et al. Increased Susceptibility to Obesity and Glucose Intolerance in Adult Female Rats Programmed by High-Protein Diet during Gestation, But Not during Lactation. Nutrients 2020, 12, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslova, E.; Rytter, D.; Bech, B.H.; Henriksen, T.B.; Rasmussen, M.A.; Olsen, S.F.; Halldorsson, T.I. Maternal Protein Intake during Pregnancy and Offspring Overweight 20 y Later. Am. J. Clin. Nutr. 2014, 100, 1139–1148. [Google Scholar] [CrossRef] [Green Version]
- Movassat, J.; Saulnier, C.; Serradas, P.; Portha, B. Impaired Development of Pancreatic Beta-Cell Mass Is a Primary Event during the Progression to Diabetes in the GK Rat. Diabetologia 1997, 40, 916–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figeac, F.; Uzan, B.; Faro, M.; Chelali, N.; Portha, B.; Movassat, J. Neonatal Growth and Regeneration of β-Cells Are Regulated by the Wnt/β-Catenin Signaling in Normal and Diabetic Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 298, E245–E256. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Ko, E.-A.; Sanders, K.M.; Chen, Q.; Zhou, T. SPORTS1.0: A Tool for Annotating and Profiling Non-Coding RNAs Optimized for RRNA- and TRNA-Derived Small RNAs. Genom. Proteom. Bioinform. 2018, 16, 144–151. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Delghingaro-Augusto, V.; Madad, L.; Chandra, A.; Simeonovic, C.J.; Dahlstrom, J.E.; Nolan, C.J. Islet Inflammation, Hemosiderosis, and Fibrosis in Intrauterine Growth-Restricted and High Fat-Fed Sprague-Dawley Rats. Am. J. Pathol. 2014, 184, 1446–1457. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm TsRNAs Contribute to Intergenerational Inheritance of an Acquired Metabolic Disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef] [Green Version]
- Grandjean, V.; Fourré, S.; De Abreu, D.A.F.; Derieppe, M.-A.; Remy, J.-J.; Rassoulzadegan, M. RNA-Mediated Paternal Heredity of Diet-Induced Obesity and Metabolic Disorders. Sci. Rep. 2015, 5, 18193. [Google Scholar] [CrossRef] [Green Version]
- Ferey, J.L.A.; Boudoures, A.L.; Reid, M.; Drury, A.; Scheaffer, S.; Modi, Z.; Kovacs, A.; Pietka, T.; DeBosch, B.J.; Thompson, M.D.; et al. A Maternal High-Fat, High-Sucrose Diet Induces Transgenerational Cardiac Mitochondrial Dysfunction Independently of Maternal Mitochondrial Inheritance. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H1202–H1210. [Google Scholar] [CrossRef] [PubMed]
- Carone, B.R.; Fauquier, L.; Habib, N.; Shea, J.M.; Hart, C.E.; Li, R.; Bock, C.; Li, C.; Zamore, P.D.; Meissner, A.; et al. Paternally-Induced Transgenerational Environmental Reprogramming of Metabolic Gene Expression in Mammals. Cell 2010, 143, 1084–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, X. Prediction of Functional MicroRNA Targets by Integrative Modeling of MicroRNA Binding and Target Expression Data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A High-Protein Diet Induces Sustained Reductions in Appetite, Ad Libitum Caloric Intake, and Body Weight despite Compensatory Changes in Diurnal Plasma Leptin and Ghrelin Concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef]
- Sluijs, I.; Beulens, J.W.J.; Spijkerman, A.M.W.; Grobbee, D.E.; van der Schouw, Y.T. Dietary Intake of Total, Animal, and Vegetable Protein and Risk of Type 2 Diabetes in the European Prospective Investigation into Cancer and Nutrition (EPIC)-NL Study. Diabetes Care 2010, 33, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Tinker, L.F.; Sarto, G.E.; Howard, B.V.; Huang, Y.; Neuhouser, M.L.; Mossavar-Rahmani, Y.; Beasley, J.M.; Margolis, K.L.; Eaton, C.B.; Phillips, L.S.; et al. Biomarker-Calibrated Dietary Energy and Protein Intake Associations with Diabetes Risk among Postmenopausal Women from the Women’s Health Initiative1234. Am. J. Clin. Nutr. 2011, 94, 1600–1606. [Google Scholar] [CrossRef]
- van Nielen, M.; Feskens, E.J.M.; Mensink, M.; Sluijs, I.; Molina, E.; Amiano, P.; Ardanaz, E.; Balkau, B.; Beulens, J.W.J.; Boeing, H.; et al. Dietary Protein Intake and Incidence of Type 2 Diabetes in Europe: The EPIC-InterAct Case-Cohort Study. Diabetes Care 2014, 37, 1854–1862. [Google Scholar] [CrossRef] [Green Version]
- van Nielen, M.; Feskens, E.J.M.; Rietman, A.; Siebelink, E.; Mensink, M. Partly Replacing Meat Protein with Soy Protein Alters Insulin Resistance and Blood Lipids in Postmenopausal Women with Abdominal Obesity. J. Nutr. 2014, 144, 1423–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenca-Sánchez, M.; Navas-Carrillo, D.; Orenes-Piñero, E. Controversies Surrounding High-Protein Diet Intake: Satiating Effect and Kidney and Bone Health. Adv. Nutr. 2015, 6, 260–266. [Google Scholar] [CrossRef]
- Watkins, A.J.; Sinclair, K.D. Paternal Low Protein Diet Affects Adult Offspring Cardiovascular and Metabolic Function in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, H1444–H1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morselli, E.; Santos, R.S.; Criollo, A.; Nelson, M.D.; Palmer, B.F.; Clegg, D.J. The Effects of Oestrogens and Their Receptors on Cardiometabolic Health. Nat. Rev. Endocrinol. 2017, 13, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Dearden, L.; Bouret, S.G.; Ozanne, S.E. Sex and Gender Differences in Developmental Programming of Metabolism. Mol. Metab. 2018, 15, 8–19. [Google Scholar] [CrossRef]
- Bertin, E.; Gangnerau, M.-N.; Bailbé, D.; Portha, B. Glucose Metabolism and β-Cell Mass in Adult Offspring of Rats Protein and/or Energy Restricted during the Last Week of Pregnancy. Am. J. Physiol. Endocrinol. Metab. 1999, 277, E11–E17. [Google Scholar] [CrossRef]
- Skinner, M.K. Fathers’ Nutritional Legacy. Nature 2010, 467, 922–923. [Google Scholar] [CrossRef] [Green Version]
- Krout, D.; Roemmich, J.N.; Bundy, A.; Garcia, R.A.; Yan, L.; Claycombe-Larson, K.J. Paternal Exercise Protects Mouse Offspring from High-Fat-Diet-Induced Type 2 Diabetes Risk by Increasing Skeletal Muscle Insulin Signaling. J. Nutr. Biochem. 2018, 57, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-M.; Obici, S.; Dong, H.H.; Haas, M.; Lou, D.; Kim, D.H.; Liu, M.; D’Alessio, D.; Woods, S.C.; Tso, P. Impaired Insulin Secretion and Enhanced Insulin Sensitivity in Cholecystokinin-Deficient Mice. Diabetes 2011. [Google Scholar] [CrossRef] [Green Version]
- Freitas, I.N.; dos Reis Araujo, T.; Vettorazzi, J.F.; Magalhães, E.A.; Carneiro, E.M.; Bonfleur, M.L.; Ribeiro, R.A. Taurine Supplementation in High-Fat Diet Fed Male Mice Attenuates Endocrine Pancreatic Dysfunction in Their Male Offspring. Amino Acids 2019, 51, 727–738. [Google Scholar] [CrossRef]
- Cherif, H.; Reusens, B.; Ahn, M.T.; Hoet, J.J.; Remacle, C. Effects of Taurine on the Insulin Secretion of Rat Fetal Islets from Dams Fed a Low-Protein Diet. J. Endocrinol. 1998, 159, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.P.; Caffrey, R.; Marioneaux, J.; Santhekadur, P.K.; Bhat, M.; Alonso, C.; Koduru, S.V.; Philip, B.; Jain, M.R.; Giri, S.R.; et al. The PPAR α/γ Agonist Saroglitazar Improves Insulin Resistance and Steatohepatitis in a Diet Induced Animal Model of Nonalcoholic Fatty Liver Disease. Sci. Rep. 2020, 10, 9330. [Google Scholar] [CrossRef]
- Wang, Z.; Li, V.; Chan, G.C.K.; Phan, T.; Nudelman, A.S.; Xia, Z.; Storm, D.R. Adult Type 3 Adenylyl Cyclase–Deficient Mice Are Obese. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitman, J.L.; Wheeler, M.C.; Lloyd, D.J.; Walker, J.R.; Glynne, R.J.; Gekakis, N. A Gain-of-Function Mutation in Adenylate Cyclase 3 Protects Mice from Diet-Induced Obesity. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, Z.; Liang, S.; Li, Y.; Yang, L.; Lu, M.; Gu, H.F.; Xia, N. Hepatic Adenylate Cyclase 3 Is Upregulated by Liraglutide and Subsequently Plays a Protective Role in Insulin Resistance and Obesity. Nutr. Diabetes 2016, 6, e191. [Google Scholar] [CrossRef] [PubMed]
- Griffin, C.A.; Kafadar, K.A.; Pavlath, G.K. MOR23 Promotes Muscle Regeneration and Regulates Cell Adhesion and Migration. Dev. Cell 2009, 17, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Tong, T.; Shen, Y.; Lee, H.-W.; Yu, R.; Park, T. Adenylyl Cyclase 3 Haploinsufficiency Confers Susceptibility to Diet-Induced Obesity and Insulin Resistance in Mice. Sci. Rep. 2016, 6, 34179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knigge, A.; Klöting, N.; Schön, M.R.; Dietrich, A.; Fasshauer, M.; Gärtner, D.; Lohmann, T.; Dreßler, M.; Stumvoll, M.; Kovacs, P.; et al. ADCY5 Gene Expression in Adipose Tissue Is Related to Obesity in Men and Mice. PLoS ONE 2015, 10, e0120742. [Google Scholar] [CrossRef] [Green Version]
- Puigserver, P.; Rhee, J.; Donovan, J.; Walkey, C.J.; Yoon, J.C.; Oriente, F.; Kitamura, Y.; Altomonte, J.; Dong, H.; Accili, D.; et al. Insulin-Regulated Hepatic Gluconeogenesis through FOXO1–PGC-1α Interaction. Nature 2003, 423, 550–555. [Google Scholar] [CrossRef]
- Lee, S.; Dong, H.H. FoxO Integration of Insulin Signaling with Glucose and Lipid Metabolism. J. Endocrinol. 2017, 233, R67–R79. [Google Scholar] [CrossRef] [Green Version]
- Altomonte, J.; Richter, A.; Harbaran, S.; Suriawinata, J.; Nakae, J.; Thung, S.N.; Meseck, M.; Accili, D.; Dong, H. Inhibition of Foxo1 Function Is Associated with Improved Fasting Glycemia in Diabetic Mice. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E718–E728. [Google Scholar] [CrossRef]
- Wang, H.; Song, Y.; Wu, Y.; Kumar, V.; Mahato, R.I.; Su, Q. Activation of DsRNA-Dependent Protein Kinase R by MiR-378 Sustains Metabolic Inflammation in Hepatic Insulin Resistance. Diabetes 2021, 70, 710–719. [Google Scholar] [CrossRef]
- Gu, N.; You, L.; Shi, C.; Yang, L.; Pang, L.; Cui, X.; Ji, C.; Zheng, W.; Guo, X. Expression of MiR-199a-3p in Human Adipocytes Is Regulated by Free Fatty Acids and Adipokines. Mol. Med. Rep. 2016, 14, 1180–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.-T.; Li, C.-L.; Tian, H.; Li, J.; Pei, Y.; Liu, Y.; Gong, Y.-P.; Fang, F.-S.; Sun, B.-R. MiR-199a Is Overexpressed in Plasma of Type 2 Diabetes Patients Which Contributes to Type 2 Diabetes by Targeting GLUT4. Mol. Cell Biochem. 2014, 397, 45–51. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | |
|---|---|---|
| Foxo1 | F | CTCACACATCTGCCATGAACCG |
| R | AAATCCAAGGTATCTCCGTCCA | |
| Pparg | F | CCACACTATGAAGACATCCCGTT |
| R | ATGCTTTATCCCCACAGACTCG | |
| Pparα | F | CTTCATCACCCGAGAGTTCCT |
| R | TCATCCAGTTCGAGGGCATT | |
| Adcy3 | F | CGGCCATGGTGGAGATACTTA |
| R | CCAAGATGAGAAGGCCACGA | |
| Adcy5 | F | GGGAGAACCAGCAACAGG |
| R | CATCTCCATGGCAACATGAC | |
| Fto | F | CGCCGCATGTCAGACCTTC |
| R | TCCACTTCATCATCGCAGGAC | |
| Cyclophilin A | F | AACCCCACCGTGTTCTTCGAC |
| R | TGCCTTCTTTCACCTTCCCAA | |
| Hprt1 | F | TTGTTGGATATGCCCTTGACT |
| R | CCGCTGTCTTTTAGGCTTTG | |
| Tbp | F | CGTGAATCTTGGCTGTAAACTTGA |
| R | GCTGCTAGTCTGGATTGTTCTTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, P.; Bailbé, D.; Bianchi, L.; Pommier, G.; Liu, J.; Tolu, S.; Stathopoulou, M.G.; Portha, B.; Grandjean, V.; Movassat, J. Paternal High-Protein Diet Programs Offspring Insulin Sensitivity in a Sex-Specific Manner. Biomolecules 2021, 11, 751. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050751
Gong P, Bailbé D, Bianchi L, Pommier G, Liu J, Tolu S, Stathopoulou MG, Portha B, Grandjean V, Movassat J. Paternal High-Protein Diet Programs Offspring Insulin Sensitivity in a Sex-Specific Manner. Biomolecules. 2021; 11(5):751. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050751
Chicago/Turabian StyleGong, Pengfei, Danielle Bailbé, Lola Bianchi, Gaëlle Pommier, Junjun Liu, Stefania Tolu, Maria G. Stathopoulou, Bernard Portha, Valérie Grandjean, and Jamileh Movassat. 2021. "Paternal High-Protein Diet Programs Offspring Insulin Sensitivity in a Sex-Specific Manner" Biomolecules 11, no. 5: 751. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050751